
古茶树种质资源遗传多样性ISSR分析
2021-06-22 07:52赵德刚赵懿琛
种子 2021年5期
刘 青, 赵德刚,2, 赵懿琛
(1.贵州大学山地植物资源保护与种质创新省部共建教育部重点实验室/贵州省农业生物工程重点实验室/生命科学学院/茶学院, 贵阳 550025;2.国家农业部植物新品种DUS测试贵阳分中心/贵州省农业科学院, 贵阳 550006)
古茶树(Camelliasinensis)是指生长在自然林中且树龄超过百年的野生茶树,还包括半驯化的人工栽培型野生茶树以及人工栽培超过百年的古茶园中的茶树[1]。经历自然选择生长下来的古茶树具有优良的抗性基因,不需要打药施肥,制成的茶叶绿色无污染,因此,近年对古茶树的研究越来越受重视。目前关于古茶树资源的研究主要集中在遗传多样性[2-4]、生化成分多样性[5-6]、抗性研究[7-8]等方面。随着生物技术的发展,遗传多样性研究已经不再局限于形态标记[9-10]、细胞标记[11]、生理生化标记[12]等传统的研究方法,基于基因组DNA水平的分子标记技术在种质资源的遗传多样性研究[13-14]、指纹图谱构建[15]、品种鉴定[16]等领域被广泛应用。
贵州是当今中国野生古茶树种质资源保存最丰富的省份之一[17],据统计,贵州省88个县级行政区域中有52个县都有古茶树生长。其中,三都水族自治县地处云贵高原,境内山峦叠嶂、土壤肥沃、矿质元素和有机物质丰富,常年云雾缭绕、无霜期长、雨量充沛、年积温可达6 600 ℃[18],是茶树的天然生长林,茶树资源丰富。
ISSR分子标记技术由于稳定性好、多态性高、成本低、DNA模板用量少[19]等特点被广泛应用于茶树的遗传多样性研究。吴田等[20]利用12条ISSR引物对17份云南茶树种质资源的亲缘关系进行分析,结果表明遗传相似系数介于0.50~0.88之间,聚类结果表明17分供试材料被分为两大类。邹瑞等[21]利用28条ISSR引物对15份紫鹃实生苗和亲本的基因组DNA进行扩增,结果发现16份材料共扩出92条带,多态性条带比率为78.3%,多样性指数(H)为 0.31,Shannon信息指数(I)为0.45,遗传一致度在0.53~0.79之间;聚类结果显示,在0.66水平上16份供试材料被分为三类,其中编号为SS 14的实生苗遗传相似度与亲本最高。目前,关于茶树遗传多样性的研究较多,但是三都地区古茶树的遗传多样性研究尚未见报道,并且在采集样品过程中发现部分古茶树受到人为破坏。因此,本研究利用ISSR分子标记技术对贵州省三都县古茶树进行遗传多样性分析,从分子水平探讨古茶树的遗传特点,为当地古茶树资源的保护和进一步研究提供理论基础。
1 材料与方法
1.1 实验材料
实验材料为贵州省三都水族自治县145份古茶树,详见表1。
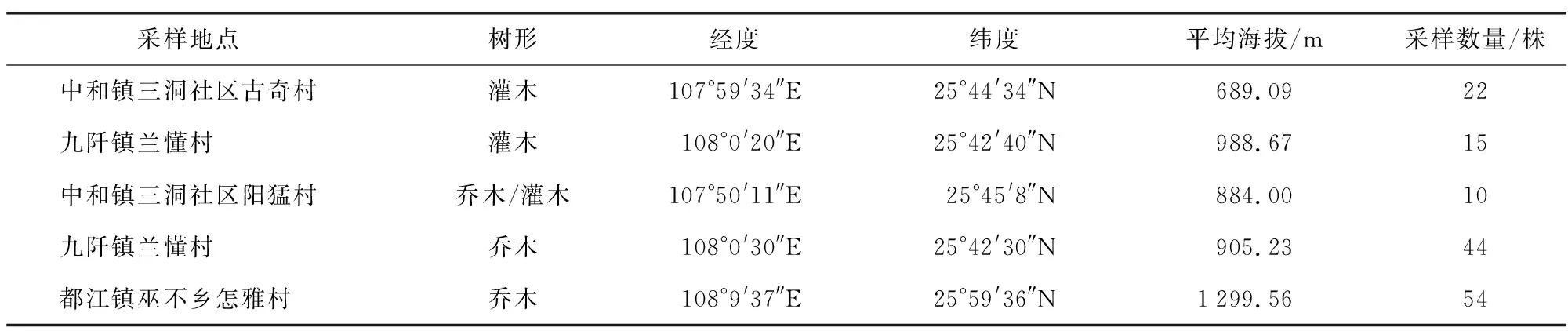
表1 用于ISSR分析的材料Table 1 Materials used for ISSR analysis
1.2 基因组DNA提取及检测
茶树基因组DNA提取参照唐祥凯等[22]的方法进行提取。用超微量全波长读数仪检测DNA的浓度和纯度,用1%的琼脂糖凝胶检测完整性。将提取的模板DNA稀释至20 ng·μL-1,于-20 ℃冰箱保存备用。
1.3 ISSR体系优化和引物筛选
选用哥伦比亚大学公布的第9套ISSR通用引物,通过对模板DNA浓度、引物浓度、MgCl2浓度、dNTP浓度、Taq酶浓度进行梯度筛选,优化PCR体系,利用优化的PCR体系设置温度梯度对引物最适退火温度进行筛选。
1.4 数据统计与分析
将3组重复的ISSR-PCR产物分别进行电泳,电泳结束后进行人工读带。将清晰的条带记为1,模糊或没有的条带记为0,建立0,1原始矩阵。统计扩增产物的总条带数和多态性条带数,其中多态性条带比率P(%)=(多态性条带数/总条带数)×100%。用Popgene 32软件统计观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(He)、shannon多样性信息指数(I)、遗传一致度。利用NTSYS pc-2.1软件按非加权组平均法(UPGMA)进行聚类分析,构建聚类图。
2 结果与分析
2.1 PCR体系优化和引物筛选
通过对酶各成分的浓度优化后得到一个最适ISSR-PCR体系(10μL),包括:10×PCR buffer,1μL;MgCl2,0.8μL;dNTPs;0.6μL;Taq酶,0.05μL;引物,0.5μL;DNA,1μL;ddH2O,6.05μL(图1)。PCR扩增体系:94 ℃预变性4 min;94 ℃变性30 s,最适退火温度退火45 s,72 ℃延伸90 s,35个循环;72 ℃延伸5 min,12 ℃保存。利用优化的PCR体系对引物进行筛选,从100条引物中筛选得到15条扩增条带清晰、重复性好、稳定性高的ISSR引物(表2),对所有模板进行扩增。
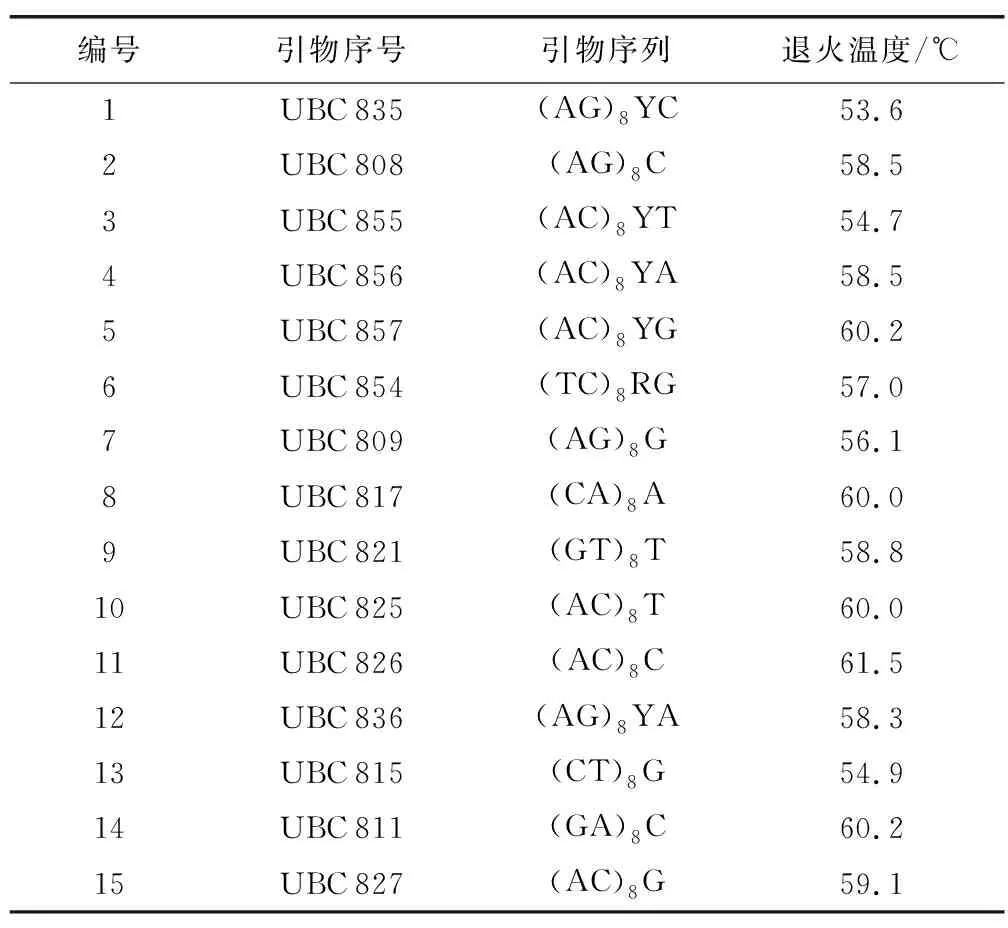
表2 引物最佳退火温度Table 2 Optimal annealing temperature for primers
2.2 引物扩增多态性分析
用筛选出的15条引物对供
试的145份材料(3个重复)进行PCR扩增(图2~图4)。从扩增结果(表3)可以看出:15条引物共扩出116条带,平均每条引物扩出7.73条,其中多态性条带110条,多态性条带比率为94.37%,不同的引物扩出的总条带数有差异,多态性条带数介于4~11之间,平均每个引物检测出的多态性位点为7.33条。多态性信息指数(PIC)在0.68~0.86之间,引物UBC 808的多态性信息指数最低,为0.68,UBC 835的多态性信息指数最高,为0.86。无论是扩增条带的大小,还是扩增条带的多少,不同引物在同一采样点古茶树样品的扩增结果存在差异,同一引物在不同材料之间的扩增结果也不同,说明供试的古茶树种质资源间存在丰富的多态性。因此,利用ISSR分子标记可以从DNA水平检测出茶树种质资源间的差异。
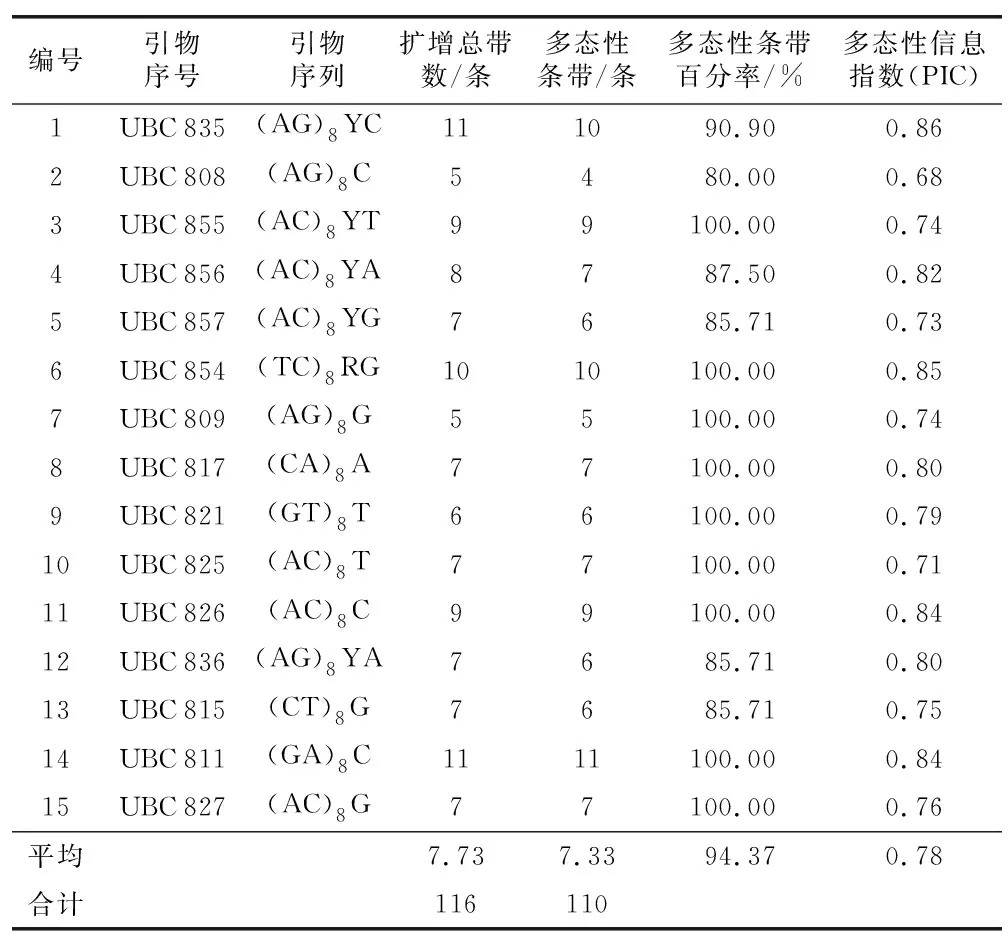
表3 ISSR 引物扩增产物的多态性Table 3 Polymorphism of amplification products byISSR markers
2.3 供试古茶树种质资源的亲缘关系分析
145份古茶树材料经过ISSR-PCR扩增,用Popgene 32软件分析各材料间遗传一致度。结果(表4)表明,145份古茶树材料间的遗传相似性系数为0.448 3~0.965 5。其中乔木间的遗传一致度为0.543 1~0.965 5,最大的是8号、9号、92号、93号乔木古茶树,最大值为0.965 5,说明其遗传背景相近,遗传相似度较高,遗传一致度最小的是27号、101号乔木古茶树,最小值为0.543 1,说明其遗传距离较远;灌木古茶树间的遗传一致度介于0.706 9~0.956 9之间,108号、109号、117号、118号、125号、127号的灌木古茶树间的遗传一致度最大,均为0.956 9,说明其遗传背景相似,遗传距离较近,118号、133号灌木古茶树的遗传一致度最小,为0.706 9,说明其遗传距离较远;乔木与灌木间的遗传距离一致度介于0.448 3~0.784 5之间,91号的乔木古茶树和137号的灌木古茶树间的遗传一致度最大为0.784 5,37号的乔木古茶树和118号的灌木古茶树的遗传一致度最小,为0.448 3,说明这2株古茶树的遗传距离较远。由表5可知,Na为1.945 7,Ne为1.529 3,He为0.312 6,I为0.471 2。说明145份古茶树材料间的遗传多样性较为丰富。
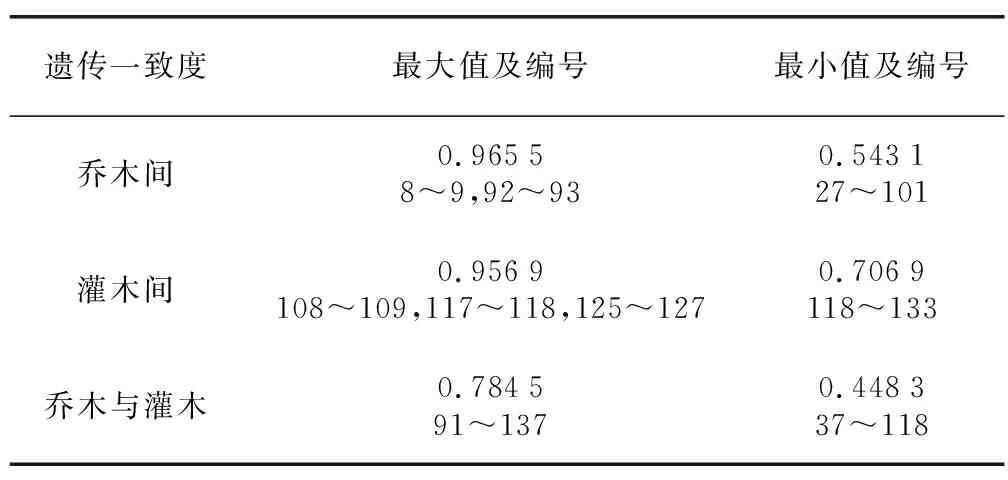
表4 145份古茶树种质资源的遗传一致度Table 4 Genetic consistency of 145 ancient tea germplasm resources

表5 供试材料的遗传多样性指数Table 5 Genetic diversity index of the tested materials
2.4 不同采样点古茶树遗传多样性参数分析
比较不同采样点样品之间同一遗传参数结果(表6)显示,不同采样点的古茶树群体具有一定的差异(p<0.05)。相对于兰懂村乔木古茶树群体,怎雅村乔木古茶树群体的等位基因数增加了3.6%,古奇村的灌木古茶树群体的等位基因数降低了19.84%,兰懂村的灌木古茶树群体的等位基因数降低了13.52%。兰懂村和怎雅村的乔木古茶树群体的有效等位基因数没有明显差异;相对于兰懂村古茶树群体的Nei’s遗传多样性,与怎雅村乔木古茶树群体的Nei’s遗传多样性指数差异不大,阳猛村乔木古茶树群体的He值降低了14.65,古奇村灌木古茶树群体的He值降低了48.17%,兰懂村灌木古茶树群体的He值降低了26.27;相对于兰懂村乔木古茶树I怎雅村乔木古茶树群体的I值增加了5.0%,其他值相对于怎雅村乔木古茶树群体的I值均较低,其中降低幅度最大的是古奇村的灌木古茶树群体,降低了47.98%;相对于兰懂村乔木古茶树群体的多样性位点数,除怎雅村增加了8.59%外,其他分别降低了17.09%、49.58%、33.76%,其中降低幅度最大的是古奇村的灌木古茶树群体;相对于兰懂村的乔木古茶树群体的多态性位点百分率(PPB),怎雅村的乔木古茶树群体增加了8.58%,PPB值最小的是古奇村的灌木古茶树群体,相对于兰懂村乔木古茶树降低了49.57%。整体来说乔木古茶树的遗传多样性比灌木古茶树的遗传多样性丰富。
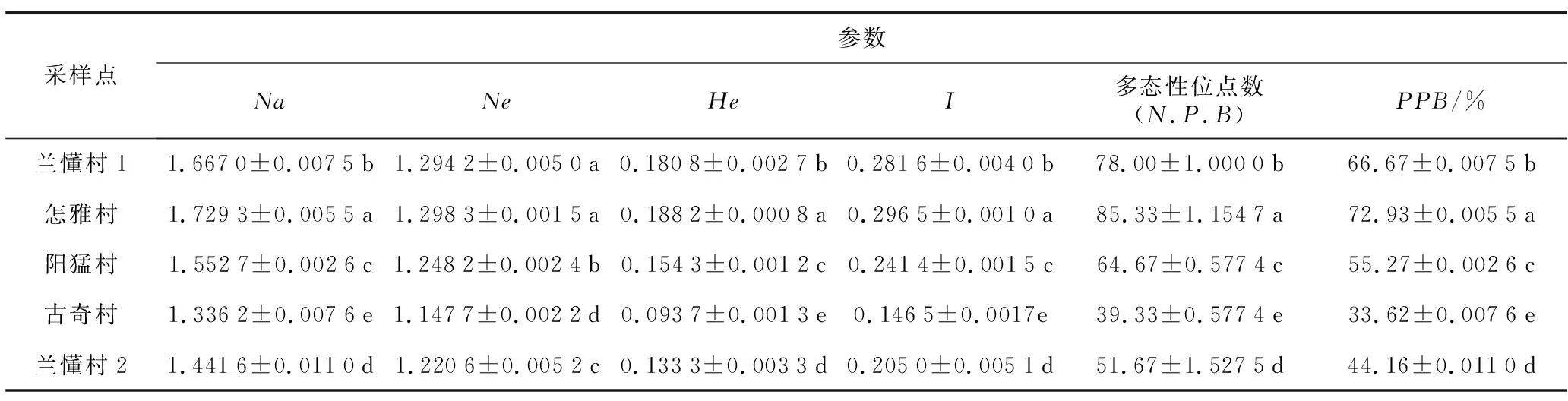
表6 5个群体的遗传多样性参数分析Table 6 Genetic diversity analysis of 5 populations
2.5 UPGMA聚类分析
利用NTSYS-pc 2.1软件对 145份古茶树材料进行聚类分析,得到相应的UPGMA树状图(图5),在遗传相似性系数为0.72时,将145份供试古茶树材料分为四大类。第一大类包括44份兰懂村的乔木古茶树;第二类包括54份怎雅村乔木古茶树;第三类包含阳猛村的9份乔木古茶树,第四类包括两个亚类,一个亚类包括22份古奇村的灌木古茶树,另一个亚类包括15份兰懂村的灌木古茶树和1份阳猛村的灌木古茶树,说明这2个地方的古茶树遗传背景相似,亲缘关系较近,聚为一类。从聚类结果可以看出,灌木古茶树与乔木古茶树的遗传距离较大,亲缘关系较远,符合表型聚类结果,并且聚类结果与分布地域有很大的关系,基本上同一地域的古茶树聚为一类。表明ISSR分子标记可用于古茶树种质资源的分类鉴定及亲缘关系研究。
3 讨 论
本研究以三都野生古茶树为研究对象,分析其遗传多样性。PCR反应过程中,反应体系的每一种成分都可能对扩增结果产生影响,浓度偏低会导致扩增不充分或背景模糊,浓度过高会引起非特异性扩增,影响实验结果,因此本研究对模板DNA、引物、MgCl2、dNTP、Taq酶的浓度进行了优化。从扩增结果看,15条引物共扩增出116条带,其中多态性条带有110条,平均多态性条带比率为94.37%,引物的多态性不仅反映了古茶树种质基因组的多态性,也反映了遗传变异的差异性。从供试的145份茶树资源分析结果来看,古茶树材料两两间遗传一致度为0.448 3~0.965 5,说明古茶树各株系之间的遗传背景差异较大,遗传多样性较丰富。古茶树材料的Na为1.945 7,Ne为1.529 3,He为0.312 6,I为0.471 2。前期有研究表明,贵州省26个县的古茶树有丰富的遗传多样性[2],本研究主要分析三都水族自治县145份古茶树的遗传多样性,结果表明,该地区古茶树的遗传多样性较丰富,但与前期研究结果相比偏低,说明生长在一定区域内古茶树比全省范围内古茶树基因交流的程度低。另外,从UPGMA聚类树状图可以看出,乔木型和灌木型古茶树能够明显被区分开,并且聚类结果与样品生长点密切相关,说明该地区古茶树基因的稳定性和聚集性。产生这种现象的原因可能是:首先,茶树经过长期的异花授粉,遗传背景复杂[23],且古茶树经历长期的自然选择和进化,使其产生一定数量的遗传变异,通过群体内个体间的自然杂交,其基因交流不断丰富,从而形成了三都水族自治县古茶树资源丰富的遗传多样性。其次,该地区群山环绕、交通不便、地域的隔离阻断了群体间的基因交流,也没有经过人为的良种选育,因此形成了该地区茶树资源遗传基因的稳定性和聚集性。
茶树的树形是由乔木型向灌木型进化的[24]。本研究4个采样点的古茶树群体的遗传多样性水平存在差异:怎雅村>兰懂村1>阳猛村>兰懂村2>古奇村,其中前2个采样点的古茶树类型为乔木型,阳猛村有1株灌木型古茶树,后2个采样点的古茶树类型为灌木型。本研究结果表明,乔木型古茶树的各遗传多样性参数均高于灌木型古茶树,结果与张德全等[25]研究结果一致。从时间上,相比灌木,乔木为原始类型,经历了更长时间的自然选择,在不断变化的自然环境中生存下来,保留了更多的优良基因,因此乔木的遗传多样性较高。从垂直分布格局上,在一定的海拔范围内茶树居群的遗传多样性随海拔梯度的增加呈现低-高-低的分布[26-27],随着海拔的不断升高,环境中温度、水分等胁迫增强,植株为了能够保持稳定生长,就会通过伸长枝条增加抗胁迫能力[26,28-29],因此,乔木型古茶树多生长在海拔较高的地方,且遗传多样性较高。
本研究利用ISSR分子标记技术分析了三都古茶树的遗传多样性。结果发现,乔木型比灌木型古茶树遗传多样性高,且该地区的古茶树群体间有一定的遗传稳定性和聚集性,群体内具有丰富的遗传多样性。研究结果为该地区古茶树的保护和进一步研究提供理论基础。
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
油气地质与采收率(2022年3期)2022-05-20
自然灾害学报(2022年2期)2022-05-10
中国学校体育(2021年10期)2021-04-26
农业与技术(2020年5期)2020-03-25
诗潮(2019年10期)2019-11-19
阅读与作文(英语高中版)(2019年9期)2019-11-07
读写算·小学中年级版(2017年4期)2017-05-08
诗潮(2017年2期)2017-03-16
读写算·小学中年级版(2015年12期)2015-12-12
