
球孢白僵菌Bbchitinase 1和Bbchitinase 2在侵染宿主过程中的不同作用
2021-06-15 13:25汪永松耿涛卢芙萍武华周娄德钊涂娜娜王树昌
热带作物学报 2021年4期
关键词:家蚕
汪永松 耿涛 卢芙萍 武华周 娄德钊 涂娜娜 王树昌


摘 要:球孢白僵菌(Beauveria bassiana)在宿主昆蟲体壁穿透和体内定植过程中分泌了Bbchitinase 1和Bbchitinase 2两种胞外几丁质酶,但二者的作用分工尚不明确。本文通过菌株胞外总几丁质酶活性与侵染毒力的相关性分析,Bbchitinase 1和Bbchitinase 2基因表达水平与侵染毒力的相关性分析,Bbchitinase 1和Bbchitinase 2的生物信息学分析以及在侵染过程中的表达模式分析,以期明晰Bbchitinase 1和Bbchitinase 2在菌株侵染宿主过程中的作用分工。结果表明,菌株胞外几丁质酶活性与侵染毒力呈正相关,但Bbchitinase 1和Bbchitinase 2表达水平不同。菌株间Bbchitinase 1表达水平与菌株侵染毒力(MMT和LT50)呈显著正相关,而Bbchitinase 2表达水平却与侵染毒力不相关。生物信息学分析结果显示,Bbchitinase 1和Bbchitinase 2具有显著的结构和功能差异,可能结合不同的配体蛋白,参与不同的代谢过程。表达模式分析结果表明,Bbchitinase 1和Bbchitinase 2基因在侵染宿主过程中均上调表达;但Bbchitinase 1仅在侵染家蚕的体壁穿透阶段显著上调表达,而Bbchitinase 2在芽生孢子生成阶段和菌丝大量增殖阶段显著上调表达。以上研究结果说明,Bbchitinase 1可能参与菌株侵染早期的侵入钉形成和体壁穿透过程,与侵染毒力呈正相关;而Bbchitinase 2可能参与侵染过程中的芽生孢子发育和菌丝大量增殖代谢调控。本研究明晰了球孢白僵菌不同几丁质酶在侵染过程中的作用分工,建立了以Bbchitinase 1基因表达水平判定菌株毒力的数学模型,为高毒、高稳定性球孢白僵菌生防菌株的筛选提供重要科学依据,对提升绿色生态防治应用效果具有重要意义。
关键词:球孢白僵菌;毒力因子;几丁质酶;家蚕
中图分类号:S476.12 文献标识码:A
Different Roles of Beauveria bassiana Bbchitinase 1 and Bbchitinase 2 in Host Infection
WANG Yongsong1, GENG Tao2, LU Fuping2, WU Huazhou2, LOU Dezhao1, TU Nana1, WANG Shuchang2*
1. Hainan University, Haikou, Hainan 570228, China; 2. Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China
Abstract: Two extracellular chitinases were secreted by Beauveria bassiana during the infection process, named as Bbchitinase 1 and Bbchitinase 2, but the roles were not clear yet. The correlation analysis between chitinase activity and virulence, correlation analysis between gene expression level and virulence, bioinformatics analysis and expression patterns assay were performed in this paper in order to clarify different roles of Bbchitinase 1 and Bbchitinase 2. Statistical analysis and inhibition test showed that extracellular chitinase activitiy was positively correlated with the virulence, while the expression levels of Bbchitinase 1 and Bbchitinase 2 were different in strains. The expression level of Bbchitinase 1 was positively correlated with the virulence of the strains (MMT and LT50); meanwhile, there was no significant relationship between Bbchitinase 1 and virulence of the strains. In addition, the bioinformatics analysis showed that there were significant differences in structure and function of Bbchitinase 1 and Bbchitinase 2, which might combine with different ligands and participate in different metabolic processes. Furthermore, the spatiotemporal expression pattern analysis showed that Bbchitinase 1 was significantly up-regulated only in the formation of penetration peg and penetration of perisoma stage of infection, meanwhile Bbchitinase 2 was slightly up-regulated in the formation of blastospore stage and significantly up-regulated in the mycelial vegetative growth stage. The results suggested that Bbchitinase 1 might be involved into body wall penetration at early infection stage and had positive correlation with virulence, while Bbchitinase 2 into the proliferation and metabolism of mycelium at the later infection stage. The results of this study clarified the role of different chitinases in the infection process of B. bassiana, and established a mathematical model to determine the virulence of B. bassiana based on the expression level of Bbchitinase 1 gene, which provides the important scientific basis for screening of high toxicity and high stability biocontrol strains and great significance for improving the application effect of green ecological control.
Keywords: Beauveria bassiana; virulence factor; chitinase; Bombyx mori
DOI: 10.3969/j.issn.1000-2561.2021.04.027
球孢白僵菌(Beauveria bassiana)是研究最早、应用最广的生防病原微生物[1-2]。具有安全性高、兼容性好、适应性强、寄生性广、孢子易培养、专性寄生可塑性强等特点,是最好的生防应用真菌[3]。然而球孢白僵菌具有侵染毒力稳定性差,菌株退化严重,防治速度慢,应用成本高等缺点。需要深入研究和明确毒力因子的作用机制,并作为菌株毒力的量化指标。
病原真菌侵染宿主昆虫是侵入钉机械压力和胞外水解酶系共同作用的结果[4-5]。菌株在孢子附着与萌发、侵入钉发育和体内定植等侵染过程中合成分泌了几丁质酶、蛋白酶、酯酶、淀粉酶、脂肪酶等水解酶类[6]。几丁质是昆虫体壁前表皮和后表皮的主要成分,构成了体壁的刚性结构和防御功能,因此病原真菌的几丁质酶活性成为侵染毒力的首要关键毒力因子[7]。
目前已经从常用生防真菌如球孢白僵菌中鉴定出2个几丁质酶基因(Bbchitinase 1和Bbchitinase 2),金龟子绿僵菌(Metarhizium anisopliae)中鉴定出5个几丁质酶基因(Machit 1、2、3、4、5),黄绿绿僵菌(Metarhizium flavoviride)中鉴定出2个几丁质酶基因(Mfchit 1和Mfchit 2)[8-10]。生物信息学分析和功能研究表明,不同菌株的胞外几丁质酶基因氨基酸序列和功能差异显著,参与侵入宿主、形态建成、营养发育、自溶增殖等生命过程[11-12]。球孢白僵菌Bbchitinase 1和Bbchitinase 2在侵染家蚕幼虫的过程中均上调表达[4]。郝蕾蕾等[13]过表达Bbchitinase 1显著提高了白僵菌对桃蚜的毒力,而Ramanujam等[14]过表达Machit 1却未能提高金龟子绿僵菌对烟草天蛾(Manduca sexta)的毒力。因此不同真菌的Chitinase 1的具体功能尚未统一,而Bbchitinase 2的具体功能亦不明确。
本研究拟通过胞外几丁质酶活性测定、几丁质酶抑制剂试验、基因表达水平分析、侵染毒力相关性分析、生物信息学分析以及侵染家蚕过程中时空表达模式分析等试验,研究分析球孢白僵菌Bbchitinase 1和Bbchitinase 2在菌株侵染宿主过程中可能的作用分工,对球孢白僵菌侵染毒力量化指标的确定和高毒菌株的分子改造具有重要科学意义。
1 材料与方法
1.1 材料
球孢白僵菌Bbcaasipp-Hv菌株、Bbcaasipp-Lv菌株由中国农业科学院植物保护研究所吴圣勇副研究员惠赠;球孢白僵菌HN6、BbMQ-04、B8、BbMQ-09菌株保存于江苏科技大学蚕业研究所家蚕生理病理研究室;家蚕(Bombyx mori)p50(大造),由江苏科技大学蚕业研究所提供。
1.2 方法
1.2.1 单孢子悬液制备 取球孢白僵菌菌株原种,接种于虫尸粉PDA平板上,(27±1)℃,培养10 d;刮下孢子,加入适量0.15 mol/L NaCl溶液,充分吹打;将孢子和菌丝的混合液转移至50 mL离心管,滴加1滴吐温80,漩涡震荡10 min,使孢子分散,用灭菌棉球过滤,即得到单孢子悬液;用血细胞计数板计数,稀释至终浓度为1×108个/mL。
1.2.2 不同菌株侵染毒力测定 取5龄起家蚕幼虫,浸于单孢子悬液中浸泡5 s,进行接种,以浸入无菌水为对照;每组50头幼虫,每组2次重复;每隔6 h记录发病数和死亡数;用Finney概率分析法和SSPS 20.0软件比较分析家蚕半数发病时间(MMT)和半数死亡时间(LT50)及95%置信区间。
1.2.3 不同菌株胞外几丁质酶活性的测定 取球孢白僵菌单孢子悬液,按1%(V/V)比例接种到虫尸粉PDA液态培养基中,(27±1)℃,120 r/min,培养4 d;真空泵抽滤,5000 r/min离心,上清液即为粗酶液;按照耿涛[4]的方法绘制标准曲线和测定酶活性。用KMO test和Bartlett test分析酶活性与侵染毒力的相关性。
1.2.4 几丁质酶抑制剂处理 分别取200 μL球孢白僵菌单孢子悬液,于2个1.5 mL无菌EP管中,分别加入200 μL H2O(对照组)、200 μL 10 μmol/L几丁质酶抑制剂Allosamidin(实验组),充分混合均匀,制成工作孢子悬液;取5 μL工作孢子悬液,涂布于5龄起蚕体壁上,1皿1头蚕饲育;每组50头幼虫,每组2次重复;每隔8 h记录家蚕幼虫的发病数和死亡数,用SSPS 20.0软件进行统计分析,计算MMT和LT50,用T-test检测差异显著性(P<0.05,以*標记)。
1.2.5 不同菌株Bbchitinase 1和Bbchitinase 2表达水平测定 取球孢白僵菌菌株原种,接种于虫尸粉PDA平板上,(27±1)℃,培养8 d;刮下菌丝和孢子,液氮速冻,研磨充分;按照1.2.8提取和纯化总RNA,完成cDNA的合成和qRT-PCR。用KMO test和Bartlett test分析基因表达水平与侵染毒力的相关性。
1.2.6 Bbchitinase 1和Bbchitinase 2生物信息学分析 应用邻接法(neighbor joining)和MEGA 7.0软件构建Bbchitinase 1和Bbchitinase 2基因的系统进化树;用SMART和SWISS-MODEL在线分析软件和数据库建立Bbchitinase 1和Bbchitinase 2蛋白二级和三级结构模型并分析其功能。
1.2.7 Bbchitinase 1和Bbchitinase 2侵染家蚕过程中表达模式分析 取球孢白僵菌单孢子悬液,浸泡感染家蚕2龄幼虫,每隔6 h采集2龄蚕;5头幼虫1管,为1个样本,每个样本2次重复;液氮冻存。按照1.2.8提取和纯化总RNA,完成cDNA的合成和qRT-PCR。
1.2.8 样本总RNA提取、纯化、cDNA的合成和qRT-PCR 按照RNA真菌提取试剂盒(OMEGA)说明书完成菌株Total RNA的提取和纯化。按照PrimeScript Kit(TaKaRa)说明书合成cDNA。
登录NCBI,依据Bbchitinase 1(GenBank登录号:GU073382.1)、Bbchitinase 2(GenBank登录号:AY147011.1)和Bbtubulin(GenBank登录号:AJ312228)的CDS序列;应用Primer 5.0引物设计软件设计实时定量引物,序列为Bbchitinase 1:F5′-CCATTGTGCCGATTCTGA-3′,R5′-AGG TTGGTCTGCGAGGTG-3′;Bbchitinase 2:F5′-AGT CGCTGCTCTAAT-3′,R5′-TCTGTTGGCTGGA A-3′;Bbtubulin:F5′-TCACCTCCAGACCG-3′,R5′-CCACAACGAAATCAA-3′。
按照TaKaRa说明书,应用ABI 7300 qRT-PCR仪完成qRT-PCR,反应条件为:95 ℃预变性5 min;95 ℃变性10 s,55 ℃退火10 s,70 ℃延伸30 s,35个循环。用2-△△CT计算基因相对表达量。
2 结果与分析
2.1 球孢白僵菌胞外总几丁质酶活性与侵染毒力的相关性分析
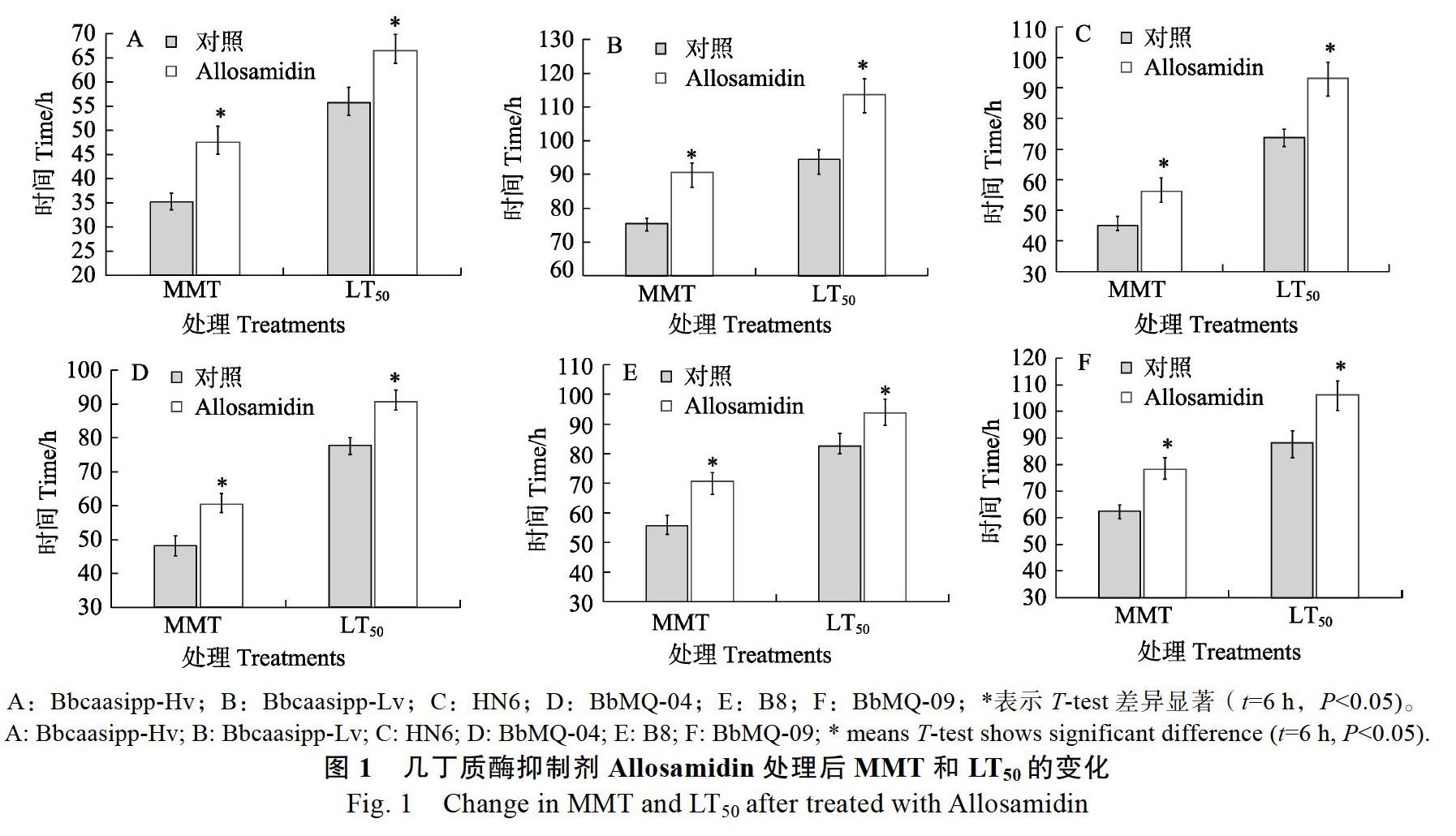
各菌株的MMT和LT50见表1,侵染毒力强弱依次为Bbcaasipp-Hv>HN6>BbMQ04>B8>BbM Q09>Bbcaasipp-Lv。各菌株胞外总几丁质酶活性见表2,酶活性高低依次为Bbcaasipp-Hv>HN6> BbMQ09>Bbcaasipp-Lv>BbMQ04>B8。抑制试验结果表明,抑制球孢白僵菌几丁质酶活性则显著降低其侵染毒力(图1)。相关性分析表明,Bbcaasipp-Hv、HN6、BbMQ04、B8菌株侵染毒力与胞外几丁质酶活性呈显著正相关,而BbMQ09和Bbcaasipp-Lv菌株为统计异质因子(图2),即酶活性相对较高,但是侵染毒力却相对较低,说明BbMQ09和Bbcaasipp-Lv菌株的胞外几丁质酶组成成分可能与其他菌株不同。
2.2 Bbchitinase 1和Bbchitinase 2表达水平与侵染毒力的相关性
针对2.1的结果,进一步检测了6个球孢白僵菌菌株的Bbchitinase 1和Bbchitinase 2的表达水平。如图3所示,Bbchitinase 1在不同菌株间的表达水平差异显著(均方=72.667,P<0.05),而Bbchitinase 2在不同菌株间表达水平差异不显著(均方=9.067,P>0.05),仅在BbMQ09和Bbcaasipp-Lv菌株中相对较高。KMO test和Bartlett test检测结果表明Bbchitinase 1表达水平与菌株的MMT和LT50呈显著负相关(图4),呈阻滞曲线模型,方程见表3,可以通过检测Bbchitinase 1的表达水平来作为菌株侵染毒力的量化指标。而Bbchitinase 2的表达水平与侵染毒力无显著相关性,推测Bbchitinase 2可能参与其他生理功能。
2.3 Bbchitinase 1和Bbchitinase 2的生物信息学分析
2.3.1 构建系统进化树 应用邻接法(neighbor joining, NJ)和MEGA 7.0软件构建了Bbchitinase 1和Bbchitinase 2的NJ系统进化树,结果表明Bbchitinase 1和Bbchitinase 2亲缘关系较远,分别与蛹虫草(Cordyceps militaris)的内切几丁质酶基因(endochitinase,CCM05744,GeneBank登录号:XM006670888)和木霉(Trichoderma sp.)几丁质酶基因(GeneBank登录号:XM006968075)亲缘关系最近(图5)。
and Bbchitinase 2
2.3.2 蛋白结构和功能预测 将Bbchitinase 1和Bbchitinase 2基因的编码氨基酸序列输入SMART在线分析软件。二者均属于18家族几丁质酶,但是含有不同的Glyco-18結构域,分别属于Chitinase A families和Chitinase families 2个不同的家族。进一步用SWISS-MODEL在线分析软件构建了二者的三维模型(图6),Bbchitinase 1和Bbchitinase 2蛋白具有不同的三维结构:Bbchitinase 1和Bbchitinase 2都含有内切几丁质酶催化域(橙色),但是活性中心(辅助催化)结构有显著差异(红色),Bbchitinase 2含有一个β发卡结构;Bbchitinase 1和Bbchitinase 2均无典型的几丁质酶结合域,但是具有特殊的辅助结构域(蓝色);Bbchitinase 1的辅助结构域为β-α-α结构,而Bbchitinase 2为β-α-α-β结构。以上结果说明它们可能识别并结合不同的辅助蛋白而催化降解不同来源的几丁质,在球孢白僵菌侵染家蚕过程中起到不同的作用。
2.4 Bbchitinase 1和Bbchitinase 2的时空表达模式
依据2.3的结果,进一步检测了Bbchitinase 1和Bbchitinase 2在球孢白僵菌侵染家蚕过程中的时空表达模式,Bbchitinase 1在球孢白僵菌侵染家蚕2龄幼虫12~24 h后显著上调表达,30 h后回到正常表达水平(图7A);而Bbchitinase 2则在侵染幼虫18~30 h后上调表达,而后回到正常水平,最后在侵染家蚕2龄幼虫54 h后显著上调表达,直至幼虫死亡(84 h)(图7B)。实验结果进一步说明Bbchitinase 1和Bbchitinase 2在球孢白僵菌侵染家蚕过程中可能起到不同的作用。
3 讨论
球孢白僵菌侵染宿主昆虫的过程分为孢子附着萌发、侵入钉形成和体壁穿透以及体内定植与致死等3个阶段,其中体壁穿透是侵染成功与否的关键前提[15]。昆虫体壁由外表皮、前表皮、后表皮、皮质层组成,前表皮和后表皮是昆虫体壁的主要组成部分,由几丁质和少量嵌入蛋白构成,形成了昆虫体壁的刚性结构和防御功能[16]。一般认为球孢白僵菌通过分泌胞外几丁质酶降解昆虫体壁进而完成侵染过程。
本研究结果表明,抑制球孢白僵菌菌株胞外几丁质酶活性会显著降低其侵染毒力(图1),相关性分析结果也表明,Bbcaasipp-Hv、HN6、BbMQ04、B8菌株的侵染毒力与胞外几丁质酶活性呈显著正相关(图2);而BbMQ09和Bbcaasipp-Lv菌株胞外几丁质酶活性虽高,但侵染毒力却相对较低(表1,表2)。
球孢白僵菌在侵染宿主昆虫过程中分泌了2种几丁质酶Bbchitinase 1和Bbchitinase 2[10]。实时定量分析结果表明Bbchitinase 1在不同菌株间的表达水平差异显著且与菌株侵染毒力呈显著正相关(图4,R2=0.98),因此可以通过检测Bbchitinase 1的表达水平作为菌株侵染毒力的量化指标。而Bbchitinase 2在不同菌株间表达水平差异不显著,仅在BbMQ09和Bbcaasipp-Lv菌株中偏高一些,且与侵染毒力无显著相关性,可能参与其他生理功能。几丁质酶抑制剂对Bbchitinase 1和Bbchitinase 2均有抑制作用,因此抑制剂处理后,菌株的Bbchitinase 1酶活性显著降低,导致侵染毒力显著降低;而BbMQ09和Bbcaasipp-Lv菌株胞外几丁质酶多为Bbchitinase 2,而Bbchitinase 1含量较低,因此虽然BbMQ09和Bbcaasipp-Lv菌株胞外总几丁质酶活性较高,但菌株的侵染毒力却较低,因此成为酶活性与侵染毒力相关性分析中的异质因子(图2)。
研究表明,虫生真菌几丁质酶在形态建成、菌丝发育、营养代谢、自溶繁殖、体壁侵染等过程中起到不同的作用[17]。本研究通过生物信息学分析和时空表达模式分析进一步探索了Bbchitinase 1和Bbchitinase 2的功能。
Fang等[18]研究发现球孢白僵菌的几丁质酶基因序列与其他昆虫真菌存在显著差异。通过构建球孢白僵菌Bbchitinase 1和Bbchitinase 2的NJ系统进化树,结果表明,Bbchitinase 1和Bbchitinase 2亲缘关系较远(遗传距离>1.4)。Bbchitinase 1与蛹虫草(Cordyceps militaris)的内切几丁质酶基因(endochitinase,CCM05744,GeneBank登录号:XM006670888)亲缘关系最近(遗传距离<0.1);而与Bbchitinase 2亲缘关系最近的是木霉(Trichoderma sp.)几丁质酶基因(GeneBank登录号:XM006968075),但遗传距离>0.2(图5)。以上结果说明二者应该具有不同的功能并参与不同的代谢过程。
几丁质酶按照一级氨基酸序列和二级结构差异分为Chitinase 18和Chitinase 19两个大家族,以Chitinase 18家族基因分布最广泛[17, 19]。二级结构建模结果表明Bbchitinase 1和Bbchitinase 2虽然都属于Chitinase 18家族,但是分属于2个不同的亚型:Glyco_hydro_18和Glyco_18。
为进一步分析二者的結构差异并了解其功能差异,构建了Bbchitinase 1和Bbchitinase 2的三维结构模型(图6)。大多数几丁质酶包涵催化域和结合域,根据催化域分为内切几丁质酶和外切几丁质酶;结合域一般有2个功能:一是识别和结合几丁质,二是破坏几丁质晶体的氢键[8, 20]。从三维模型可以看出Bbchitinase 1和Bbchitinase 2都含有内切几丁质酶催化域(橙色),均为内切几丁质酶;但Bbchitinase 2的辅助催化活性中心(红色)比Bbchitinase 1多了一个β发卡结构,说明二者具有不同的催化功能。此外,区别于纤维素酶的β-β-α-β结合域,球孢白僵菌Bbchitinase 1和Bbchitinase 2没有典型的结合域,但是具有特殊的辅助结构域(图7,蓝色结构区域)。范艳华[8]研究表明Anopheles gambiae围食膜结合蛋白与几丁质酶1结合才能特异性识别几丁质,因此球孢白僵菌内应该存在特殊的配体蛋白,与Bbchitinase 1和Bbchitinase 2的辅助结构域结合而发挥几丁质的降解作用。此外,Bbchitinase 1的辅助结构域(蓝色)为β-α-α结构,有利于几丁质酶在宿主体壁几丁质表面富集[17];而Bbchitinase 2为β-α-α-β结构,可能识别并结合不同的配体蛋白,而后催化降解不同来源的几丁质,参与不同的生理功能。
Hou等[21]研究表明球孢白僵菌侵入家蚕的过程中,8~12 h为孢子附着和萌发阶段,13~28 h为侵入钉形成和穿透体壁阶段,29~45 h为芽生孢子发育增殖阶段,46 h后为营养菌丝发育和增殖阶段。时空表达模式分析结果表明,Bbchitinase 1在侵染家蚕2龄幼虫12~24 h后显著上调表达(图7A);而Bbchitinase 2则在侵染幼虫18~30 h以及54~84 h后显著上调表达,直至幼虫死亡(84 h)(图7B);说明Bbchitinase 1主要参与侵入钉形成和体壁穿透功能,而Bbchitinase 2可能参与芽生孢子和营养菌丝的发育生殖。
本研究明晰了球孢白僵菌Bbchitinase 1和Bbchitinase 2在侵染宿主过程中的作用分工;建立了以Bbchitinase 1表达水平为菌株侵染毒力量化指标的数学模型;发现Bbchitinase 2可能参与芽生孢子和营养菌丝的发育生殖过程;结果将为其他高毒力、稳定性生防真菌的筛选提供重要科学依据,对提升真菌生防制剂的田间应用效果具有重要意义。然而,Bbchitinase 1和Bbchitinase 2辅助结合蛋白和作用底物以及诱导表达机制等问题还有待深入分析。
参考文献
[1]万方浩, 叶正楚, 郭建英, 等. 我国生物防治研究的进展及展望[J]. 昆虫知识, 2000, 37(2): 65-74.
[2]Mascarin G M, Jaronski S T. The production and uses of Beauveria bassiana as a microbial insecticide[J]. World Journal of Microbiology and Biotechnology, 2016, 32(11): 177.
[3]Portilla M, Snodgrass G, Luttrell R. Lethal and sub-lethal effects of Beauveria bassiana (Cordycipitaceae) strain NI8 on Chrysoperla rufilabris (Neuroptera: Chrysopidae)[J]. Florida Entomologist, 2017, 100(3): 627-633.
[4]耿 涛. 家蚕与白僵菌互作机制的研究[D]. 镇江: 江苏科技大学, 2016.
[5]王树昌, 赫荣帆, 陆敏泉, 等. 球孢白僵菌侵染对家蚕抗氧化水平的影响[J]. 热带作物学报, 2017, 38(11): 2136-2144.
[6]康 曼. 球孢白僵菌分子生物学研究进展[J]. 广东农业科学, 2011, 38(24): 122-124.
[7]Galidevara S, Reineke A, Koduru U D. In vivo expression of genes in the entomopathogenic fungus Beauveria bassiana during infection of lepidopteran larvae[J]. Journal of Invertebrate Pathology, 2016, 136: 32-34.
[8]范艳华. 球孢白僵菌降解寄主体壁的几丁质酶和蛋白酶的分子改良[D]. 重庆: 西南大学, 2006.
[9]Sun M, Ren Q Y, Guan G Q, et al. Virulence of Beauveria bassiana, Metarhizium anisopliae and Paecilomyces lilacinus to the engorged female Hyalomma anatolicum anatolicum tick (Acari: Ixodidae)[J]. Veterinary Parasitology, 2011, 180(3/4): 389-393.
[10]甄 伟, 杜立新, 曹伟平, 等. 球孢白僵菌HFW-05几丁质酶基因的克隆与序列分析[J]. 华北农学报, 2010, 25(1): 36-39.
[11]翟素珍, 王吉平, 陈晶晶, 等. 球孢白僵菌Bb_bm01菌株PacC基因突变体的构建及其毒力和孢子萌发率的测定[J]. 基因组学与应用生物学, 2016, 35(8): 2064-2070.
[12]Chantasingh D, Kitikhun S, Eurwilaichitr L, et al. Functional expression in Beauveria bassiana of a chitinase gene from Ophiocordyceps unilateralis, an ant-pathogenic fungus[J]. Biocontrol Science and Technology, 2011, 21(6): 677-686.
[13]郝蕾蕾, 陈珊珊, 谢 翎, 等. 几丁质酶基因在球孢白僵菌中的超表达提高其对马尾松毛虫的毒力[J]. 中国生物防治, 2010, 26(3): 267-273.
[14]Ramanujam B, Balachander M, Roopa G, et al. Chitinase activity and virulence of different isolates of Beauveria bassiana, Metarhizium anisopliae and Lecanicillium spp.[J]. Journal of Biological Control, 2011, 25(3): 223-228.
[15]呂丁丁, 李增智, 王成树. 虫生真菌分子致病机理及基因工程改造研究进展[J]. 微生物学通报, 2008, 35(3): 443-449.
[16]林海萍, 魏锦瑜, 毛胜凤, 等. 球孢白僵菌蛋白酶、几丁质酶、脂肪酶活性与其毒力相关性[J]. 中国生物防治, 2008, 24(3): 290-292.
[17]杨 帆, 段立清, 宋 钢, 等. 寄生沙棘木蠹蛾的白僵菌菌株特性及其胞外蛋白酶、几丁质酶的活性[J]. 植物保护, 2011, 37(2): 67-71.
[18]Fang W, Feng J, Fan Y, et al. Expressing a fusion protein with protease and chitinase activities increases the virulence of the insect pathogen Beauveria bassiana[J]. Journal of Invertebrate Pathology, 2009, 102(2): 155-159.
[19]Jia Z C, Sun Y M, Li Y, et al. The chitinase gene (Bbchit1) from Beauveria bassiana enhances resistance to Cytospora chrysosperma in Populus tomentosa Carr[J]. Biotechnology Letters, 2010, 32(9): 1325-1332.
[20]Fan Y, Fang W, Xiao Y, et al. Directed evolution for increased chitinase activity[J]. Applied Microbiology Biotechnology, 2007, 76(1): 135-139.
[21]Hou C X, Qin G X, Liu T, et al. Transcriptome analysis of silkworm, Bombyx mori, during early response to Beauveria bassiana challenges[J]. PLoS One, 2014, 9(3): e91189.
责任编辑:谢龙莲
猜你喜欢
科学导报(2020年32期)2020-05-26
江苏农业科学(2016年11期)2017-03-21
现代农业科技(2017年2期)2017-03-16
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年10期)2017-02-05
江苏农业科学(2016年5期)2016-07-23
江苏农业科学(2016年5期)2016-07-23
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年1期)2015-04-17
湖北农业科学(2014年21期)2014-12-22
