
Two new marine benthic diatom species of Hippodonta(Bacillariophyceae) from the coast of China: H. nanjiensis sp. nov. and H. qingdaoensis sp. nov.*
2021-06-15 08:12YuhangLIZhongminSUNTamotsuNAGUMOKuidongXU
Yuhang LI, Zhongmin SUN , Tamotsu NAGUMO,2, Kuidong XU,3,4,5,**
1 Laboratory of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
2 Echigo Natural History Laboratory, Kowada 719, Ojiya, Niigata 947-0041, Japan
3 Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology(Qingdao),Qingdao 266237, China
4 University of Chinese Academy of Sciences, Beijing 100049, China
5 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
Abstract The diversity and distribution of Hippodonta species on the coast of China are largely unknown due to limited investigations. In this paper, we report the detailed morphology and ultrastructure of two new species: H. nanjiensis sp. nov., collected from intertidal sandy sediments of Nanji Islands, the East China Sea and H. qingdaoensis sp. nov., collected from intertidal sandy sediments of the No. 1 Bathing Beach in Qingdao City, the Yellow Sea. Both species occur mainly in the middle part of the intertidal zone. H.nanjiensis possesses a unique large round central area. H. qingdaoensis diff ers from congeners in having a unique stria pattern, wherein the areolae in the middle transapical striae are less developed or absent on the primary side of the valve, and two areolae per stria are apparent under light microscopy (LM). This study expands our knowledge of Hippodonta diversity and distribution on the coast of China.
Keyword: Hippodonta; intertidal sediment; new species; Yellow Sea; East China Sea
1 INTRODUCTION
The genusHippodontaLange-Bertalot, Metzeltin et Witkowski in Lange-Bertalot et al. (1996) was established and separated fromNaviculaBorysensulatobased on its characters of a simple raphe system without an accessory rib, transapically elongated and cap-like terminal areas, and external raphe distal endings not or weakly deflected without extending to the valve mantle (Witkowski et al., 2000; Pavlov et al., 2013). Lange-Bertalot et al. (1996) described ten newHippodotnaspecies and transferred 11 species into this genus, which included eight species from coastal areas. Later, three new species from coastal areas were added to the genus (Witkowski et al.,2000; Potapova, 2013; Zhao et al., 2017). In contrast to these coastal species, most species ofHippodontahave been described from (ancient) lakes and rivers in various continents (Metzeltin et al., 2005; Blanco et al., 2012; Kulikovskiy et al., 2012; Van de Vijver et al., 2012; Pavlov et al., 2013; Peng et al., 2014).Presently, 95 species are included in the genusHippodonta(Guiry 2020), of which only 11 have been described from coastal areas. Pavlov et al. (2013)separatedHippodontainto eight morphological groups. MostHippodontaspecies from coastal areas were assigned into Groups 7 and 8, both of which have a linear valve outline.
In the past decade, marine benthic diatoms from the Bohai Sea, Yellow Sea and East China Sea have received increased attention. New genera, viz.,MoreneisPark, Koh et Witkowski andSternimirusWitkowski et Li, and many new species have been described in this region (Park et al., 2012, 2013; Li et al., 2015; Liu et al., 2015a, b, c; Witkowski et al.,2016; Chen et al., 2017; Zhao et al., 2017; Li et al.,2018). Molecular data indicate additional potential new taxa in this region (Witkowski et al., 2016; An et al., 2017). However, within the genusHippodonta,only two species have been reported in this area,along the China Fujian coast of the East China Sea(Zhao et al., 2017). In the present study, we describe two newHippodontaspecies sampled from intertidal sediments of the East China Sea and the Yellow Sea,providing insight into the diversity ofHippodontafrom the coast of China.
2 MATERIAL AND METHOD
Sampling was conducted at the Dasha’ao(27°27′45.6′′N, 121°03′ 28.80′′E) sandy beach in the Nanji Islands National Marine Natural Reserve, the East China Sea, on 13 November 2016 and at the No.1 Bathing Beach, Qingdao City, the Yellow Sea(36°03′16.97′′N, 121°20′19.27′′E) on 2 September 2016 and 1 May 2019. At each site, three stations were selected for sample collection, one in each of the high, middle and low tidal zones. The upper 1–2 cm of sediment at each station was collected using a glass tube (2 cm in diameter), and glutaraldehyde was added a final concentration of 2.5% as fixative (Round et al., 1990).
Sediments (0.5 g) were cleaned using domestic bleach (Pip Unish, Uni. Charm, Tokyo) for 20 min and repeatedly washed with distilled water (via centrifugation three to five times at 1 000×gfor 7 min;Nagumo and Kobayasi, 1990; Nagumo, 1995).Cleaned samples were mounted on glass slides with Mountmedia (Wako Pure Chemical Industries, Ltd.Osaka, Japan) for light microscopy (LM) observation.A Zeiss Imager Z2 and Nikon Eclipse 80i LM equipped with diff erential interference contrast (DIC)were used for LM observation. For scanning electron microscopy (SEM) observation, cleaned samples were placed on coverslips, air-dried and coated with osmium. A Hitachi S-3400 was used for SEM observation. For transmission electron microscopy(TEM) observation, cleaned samples were placed on Formvar-coated copper mesh grids and air-dried. A JEOL-2000EX was used for TEM observations.
Environmental factors were measured at Dasha’ao sandy beach, Nanji Islands. The temperature in situ was measured using a thermometer with the bulb immersed in the sediment to a depth of 2 cm on each sampling occasion. Pore water from the upper sediment was collected for salinity measurement using a refractometer (Model: RHS-10ATC). Grain size analysis was performed using a Laser Diff raction Particle Size Analyzer (Cilas 940L). The water content of the sediment layers was measured as a percentage of weight loss by drying the sediment at 60 °C for 48 h.Terminology follows Anonymous (1975), Ross et al. (1979) and Round et al. (1990).
3 RESULT
Species description
Class Bacillariophyceae Haeckel
Order Naviculales Bessey
Family Naviculaceae Kützing
GenusHippodontaLange-Bertalot, Metzeltin et Witkowsk
HippodontananjiensisYuhang Li, Nagumo et Kuidong Xu, sp. nov. (Figs.1 & 2)
Description: Frustule rectangular with rounded corners. Valves lanceolate with protracted acutely round apices, 13.0–31.0 μm (20.9±5.9 μm, mean±SD,n=15) in length and 2.5–5.2 μm (3.7±0.7 μm,n=15)in width. Raphe straight. Axial area narrow. Central area very large. Striae radiate near center and convergent near apices, 12–16/10 μm (Fig.1a–g).
In SEM, valves are convex and depressed at the center (Fig.1h–i). The raphe external distal endings are weakly curved to the same direction at apices, not extending to the valve mantle (Figs.1h, i, 2a arrow, &b arrow). Internally distal raphe ends terminate in small helictoglossae (Fig.1j). The external proximal raphe endings are close and slightly expanded externally (Fig.2c & d). The internal proximal raphe endings are close and straight (Fig.1j). The transapical striae are radiate near center and become convergent near apices (Fig.1a–g). The striae forming areolae are apically elongate and parallel, except those adjacent to the central area which are oblique, (38–51)/10 μm(Fig.2c & d arrowheads). The areolae are occluded internally by hymenes. The central area is large and round (Figs.1h, i, 2c, & d), with one or two areolae near the valve margin (Fig.2c & d arrows). A uniseriate rows of areolae surround the apices (Fig.1h). The two central ones are longer than the others (Fig.1h arrow).
Holotype: Holotype MBMCAS286531, illustrated in Fig.1a, deposited in the Marine Biological Museum,Chinese Academy of Sciences (MBMCAS), Qingdao,China.
Isotype: MBMCAS286533, deposited in the MBMCAS, Qingdao, China.

Fig.1 LM and SEM micrographs of Hippodonta nanjiensis
Type locality: The Dasha’ao beach in the Nanji Islands, East China Sea (27°27′45.6′′N,121°03′28.80′′E).
Etymology: Species name is derived from the type locality Nanji Islands.
Distribution and ecology:Hippodontananjiensisis currently known only from the type locality, where it occurs mainly in the middle intertidal zone of the Dasha’ao sandy beach at salinities ranging from 26 to 29. The sediment temperature at collection was approximately 26℃. The water content of the sediment at collection was 31.6%. The median grain size of the sediment is 254 μm.Amphoraspp.,Carinasigmaminuta(Donkin) Reid,Donkiniaspp.andHantzschiamarina(Donkin) Grunow were dominant diatoms at the type locality.
HippodontaqingdaoensisYuhang Li, Nagumo et Kuidong Xu, sp. nov. (Figs.3 & 4)
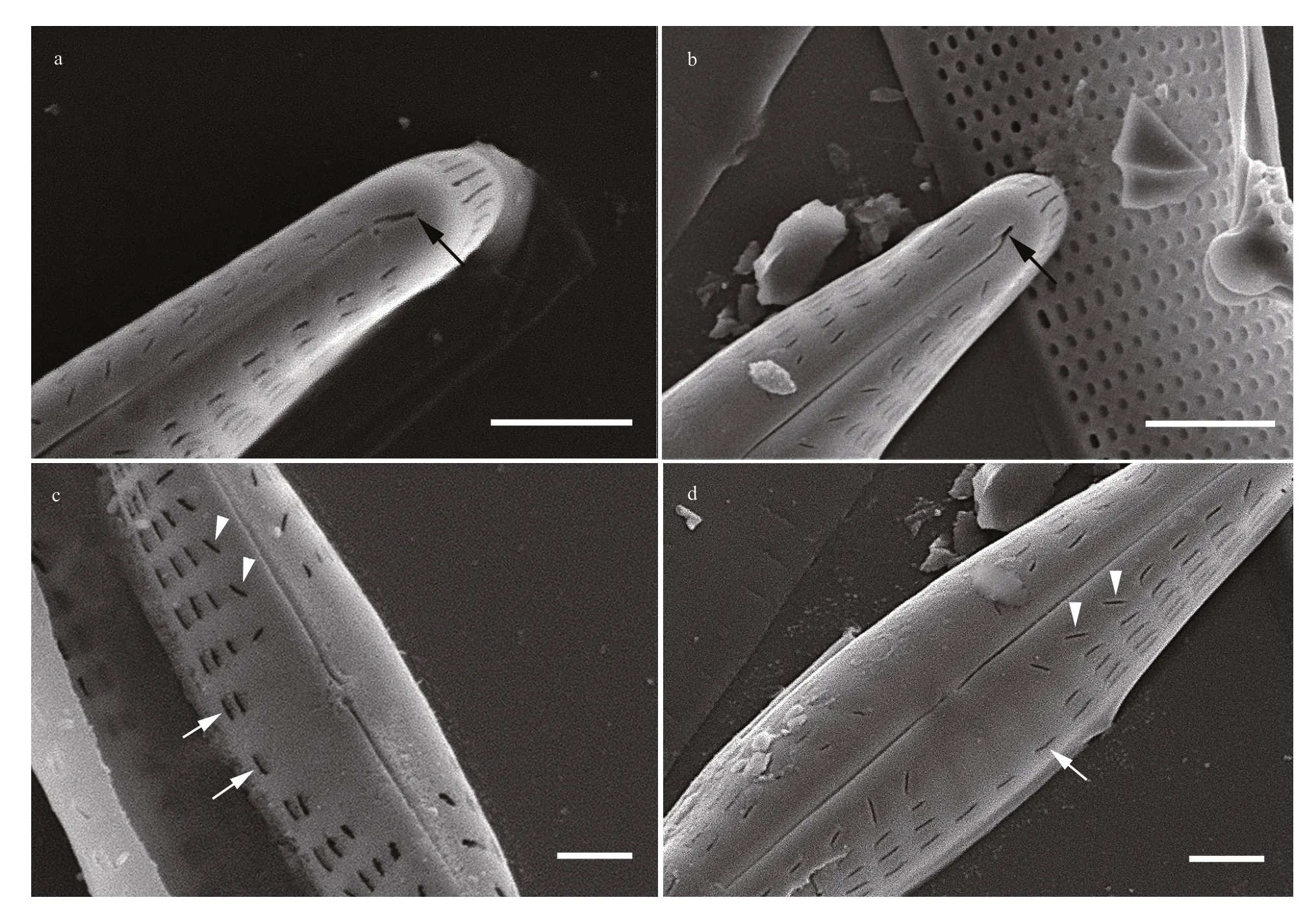
Fig.2 SEM micrographs of Hippodonta nanjiensis
Description: Valves lanceolate with obtusely round apices, 6.8–11.0 μm (8.5±1.2 μm, mean±SD,n=15) in length and 2.2–3.9 μm (3.2±0.6 μm,n=15)in width. Raphe filiform, slightly curved. Axial area narrow. Central area rectangular or bow-tie shaped.Transapical striae slightly radiate, (11–14)/10 μm.Striae appear composed of two areolae on the primary side of the valve (Fig.3a–g).
In SEM, valves are convex. The raphe external distal ends deflect to the same direction at apices, not extending to the valve mantle (Figs.3h–k, 4a, b, & d).Internally distal raphe ends terminate in small helictoglossae (Fig.3j). External proximal raphe endings are straight and slightly expanded (Fig.3h, i,& k), whereas internally raphe endings are simple and straight (Fig.3j). In TEM, the areolae in the middle transapical striae are less developed or absent on the primary side of the valve (Fig.4a–c), presenting two areolae per stria under LM (Fig.3a–g). The central area is rectangular or bow-tie-shaped, with one or two areolae near the valve margin (Figs.3a–g & 4a–b). A uniseriate row of areolae surround the apices (Fig.4d).The areolae internally occluded by hymenes, (26–33)/10 μm (Fig.4c & d).
Holotype: Holotype MBMCAS286532, illustrated in Fig.3f, deposited in the Marine Biological Museum,Chinese Academy of Sciences, Qingdao, China.
Isotype: MBMCAS286534, deposited in the MBMCAS, Qingdao, China.
Type locality: No. 1 Bathing Beach in Qingdao City, the Yellow Sea (36°03′16.97′′N, 121°20′19.27′′E).
Etymology: Species name is derived from the type locality Qingdao City.
Distribution and ecology:Hippodontaqingdaoensisis currently known only from the type locality, where it occurs mainly in the middle intertidal zone at salinities from 28 to 30. Small celledAnorthoniesexcentrica(Donkin) Grunow,Cocconeiopsiskantsiensis(Giff en) Witkowski, Lange-Bertalot et Metzeltin,Fallaciaspp. andNaviculaspp. were dominant diatoms at the type locality.
4 DISCUSSION
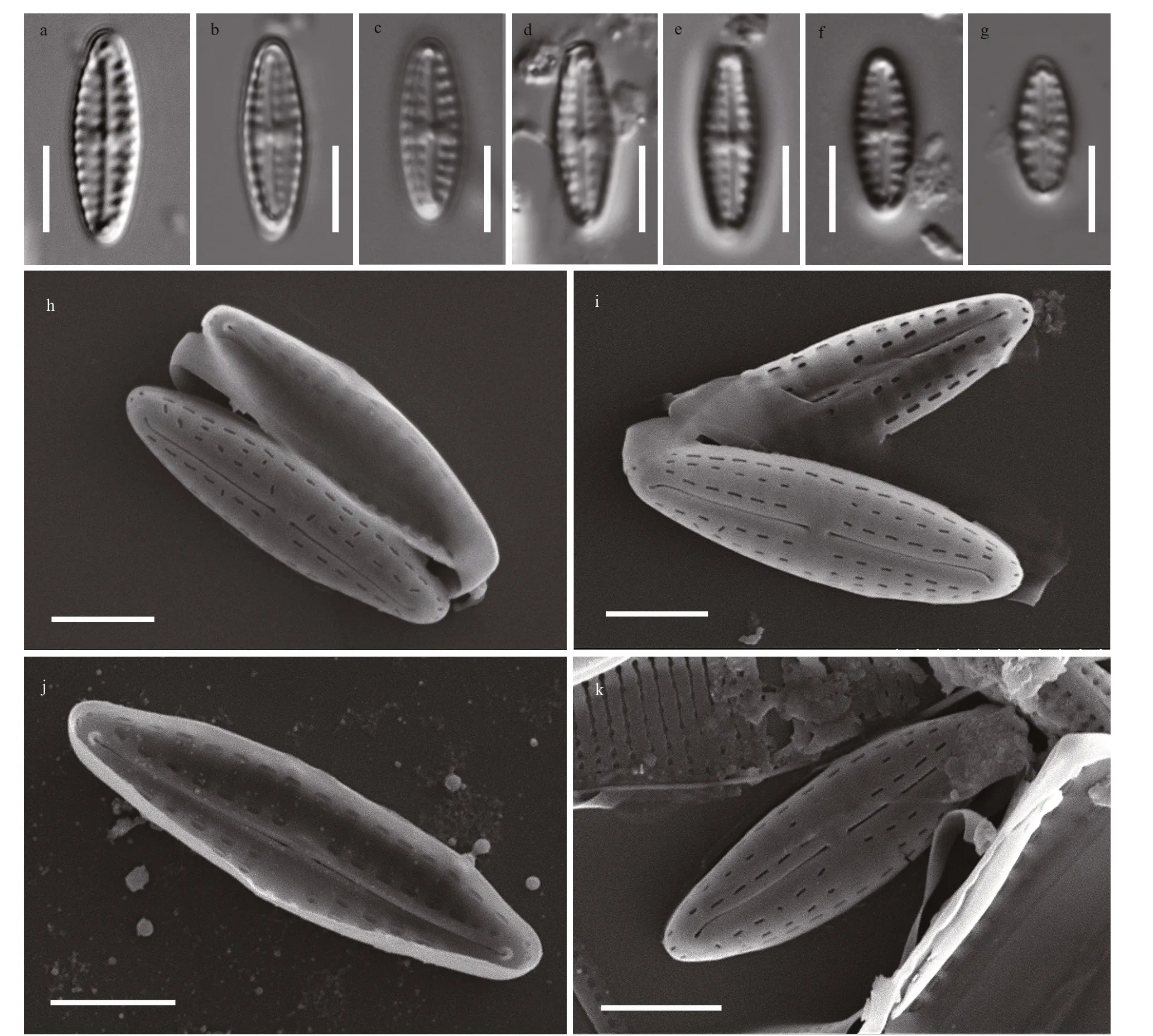
Fig.3 LM and SEM micrographs of Hippodonta qingdaoensis
BothHippodontananjiensisandH.qingdaoensishave two features that diff er from those ofNaviculas. s., viz., a simple raphe without an accessory rib on the internal valve face (vs. raphe slit opening laterally in a markedly rib-like raphe sternum for most ofits length) and raphe slits that are weakly curved at external valve apices without extending to the valve mantle (Fig.4d; vs. usually curved or hooked terminal fissures extending to the valve mantle). Both features justify the assignment of the two species to the genusHippodonta.
Hippodontananjiensishas a unique large round central area, whereas in all other congeners the central area is rhombic or lanceolate or bow-tie-shaped or rectangular fascia (Pavlov et al., 2013).Hippodontananjiensismostly closely resemblesH.subcostulata(Hustedt) Lange-Bertalot, Metzeltin et Witkowski in valve length, width, stria density, and stria orientation(Table 1). However, in addition to having a unique central area,H.nanjiensisdiff ers fromH.subcostulatain having a more acute narrow valve and a more elevated valve face.HippodontananjiensisresemblesH.grunowiiLange-Bertalot, Metzeltin et Witkowski in valve outline, length, width and stria density but diff ers from the latter in having striae on the subapical area (which are absent inH.grunowii). In addition,H.nanjiensishas a uniseriate row of areolae around the apices, with the two middle areolae being longer than the others. This feature is also exhibited byH.fujianensis, a species recently described from the southeast coast of China. However, the two species can be distinguished by the valve outline (narrowlinear with protracted apices inH.nanjiensisvs.elliptic-lanceolate to rhombic-lanceolate with nonprotracted apices inH.fujianensis), stria orientation (slightly radiate near center becoming convergent towards apices vs. radiate on the whole valve face) and the unique large round central area inH.nanjiensis.
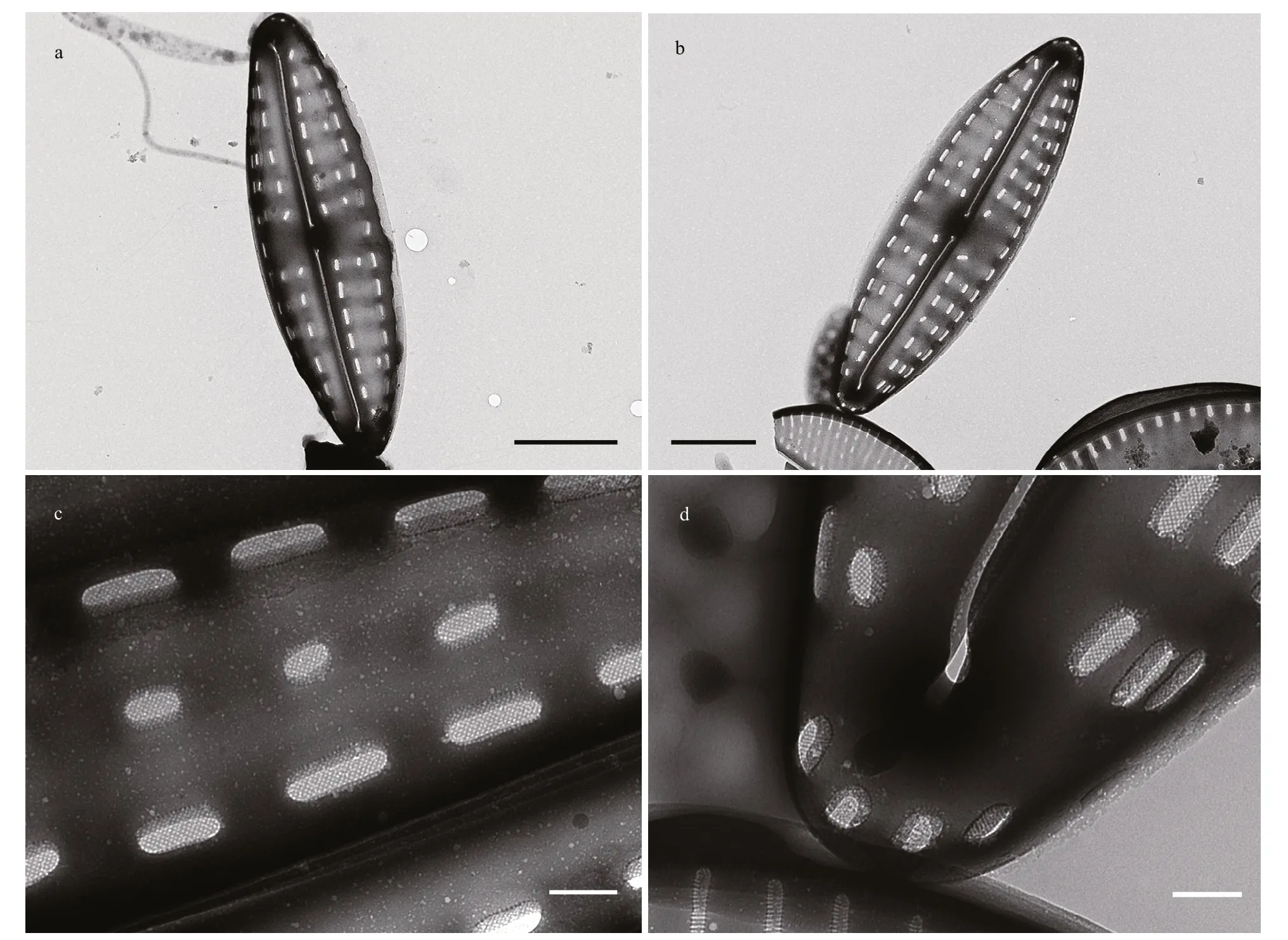
Fig.4 TEM micrographs of Hippodonta qingdaoensis
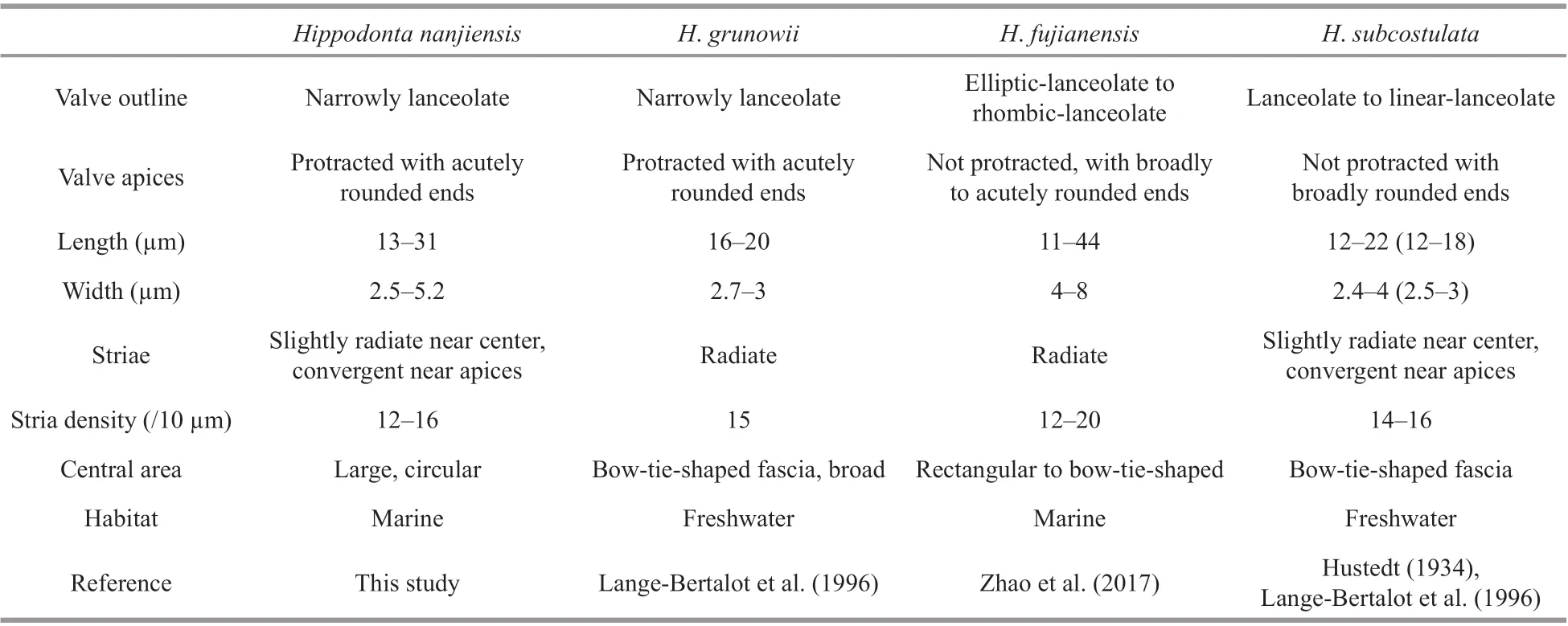
Table 1 Comparison of Hippodonta nanjiensis with H. grunowii, H. fujianensis, and H. subcostulata in morphological attributes

Table 2 Comparison of Hippodonta qingdaoensis with H. minuta, H. pseudoacceptata, H. dulcis, and H. sp. sensu Al-Handal,Compére et Riaux-Gobin ( 2016) in morphological attributes
Hippodontaqingdaoensisdiff ers from congeners in having a unique stria pattern: the areolae in the middle transapical striae are less developed or absent on the primary side of the valve, appearing two areolae per stria under LM (Table 2). One of the areolae is adjacent to the raphe and the other one is along the valve margin. This kind of stria pattern also appears inNaviculawasmundiiWitkowski, Metzeltin et Lange-Bertalot. However,Naviculawasmundiilacks the two diagnostic features ofHippodontamentioned above.Hippodontaqingdaoensismost closely resemblesHippodontaminutaPavlov,Levkov, D.M. Williams et Edlund; both species have linear-lanceolate valves with broadly rounded apices and bow-tie-shaped central areas. However, in addition to exhibiting the unique feature mentioned above,H.qingdaoensiscan be distinguished fromH.minutaby its smaller valve size (6.8–11.0 μm vs.12.5–17 μm) and higher stria density ((11–14)/10 μm vs. (8–12)/10 μm).Hippodontaqingdaoensisalso resemblesH.pseudoacceptata(Kobayasi) Lange-Bertalot andH.dulcisPotapova but diff ers from them in valve outline (linear-elliptic to linear rhombic inH.pseudoacceptataand lanceolate with acute apices inH.dulcis) and stria density, which is lower inH.qingdaoensis((11–14)/10 μm inH.qingdaoensisvs. (15–21)/10 μm inH.pseudoacceptataand (14–17)/10 μm inH.dulcis). Moreover,H.minuta,H.pseudoacceptataandH.dulcisare all originally reported from freshwater or brackish water rather than marine environments (Kobayasi and Mayama,1986; Pavlov et al., 2013; Potapova, 2013).Hippodontaqingdaoensisalso resemblesH. sp.sensuAl-Handal, Compére et Riaux-Gobin (2016). Both have a similar valve outline. However,H.qingdaoensisdiff ers from the latter species in having its unique stria pattern and a much lower stria density ((11–14)/10 μm vs. (18–20)/10 μm).
During our survey of marine benthic diatoms in Dasha’ao, Nanji Islands and Qingdao No.1 Bathing Beach, we observed fiveHippodontaspecies:H.nanjiensissp. nov.,H.qingdaoensissp. nov.,H.linearis(Østrup) Lange-Bertalot, Metzeltin, et Witkowski,H. cf.dulcisandH.fujianensis. These species represent all theHippodontaspecies recorded from the China coast of the Pacific. In contrast, among the 13Hippodontaspecies reported from coastal areas, nine were reported from the coastal areas of the Atlantic, viz.,H.caoticaWitkowski, Lange-Bertalot et Metzeltin;H.germainiiLange-Bertalot, Metzeltin et Witkowski;H.kaiseriLange-Bertalot, Metzeltin et Witkowski;H.kociolekiiLange-Bertalot, Metzeltin et Witkowski;H.lesmonenisLange-Bertalot,Metzeltin et Witkowski;H.linearis,H.ruthnielseniaeLange-Bertalot, Metzeltin et Witkowski; andH.umbilicatissima(Reichardt) Reichardt (Lange-Bertalot et al., 1996; Witkowski et al., 2000). Al-Handal et al. (2016) also mentioned sevenHippodontaspecies in the West Indian Ocean; unfortunately, they could not identify these species due to insuffi cient specimens. The data indicate thatHippodontaspecies commonly occur in coastal areas worldwide.However,Hippodontaspecies are usually low in abundance and easily confused withNaviculaspecies in routine LM examination because the diagnostic features cannot be revealed under light microscope.Thus, the diversity ofHippodontain coastal areas is presumably still underestimated. Extensive taxonomic works remain needed to reveal the diversity and distribution ofHippodonta.
5 CONCLUSION
Two new species of marine benthic diatoms,Hippodontananjiensissp. nov. andH.qingdaoensissp. nov., are described from the intertidal sandy sediments of Nanji islands in the East China Sea and Qingdao No. 1 Bathing Beach in the Yellow Sea,respectively.H.nanjiensiscan be easily distinguished by its unique large round central area.H.qingdaoensishas a unique stria pattern from that ofits congeners.This study expands our knowledge of the diversity and distribution of the genusHippodontaon the coast of China.
6 DATA AVAILABILITY STATEMENT
The authors declare that the data supporting the findings of this study are available within the article.The data will be available on request from the corresponding author.
7 ACKNOWLEDGMENT
The authors are grateful to three anonymous reviewers for providing constructive suggestions, and to Mrs. Houcai CAI, Wandong CHEN, and the staff of the Nanji Islands National Marine Natural Reserve Research Institute for their help in sample collection.
 Journal of Oceanology and Limnology2021年3期
Journal of Oceanology and Limnology2021年3期
- Journal of Oceanology and Limnology的其它文章
- Steady increase in water clarity in Jiaozhou Bay in the Yellow Sea from 2000 to 2018: Observations from MODIS*
- Phylogenetic diversity and bioactivity of culturable deepsea-derived fungi from Okinawa Trough*
- Allelopathic eff ects of mixotrophic dinoflagellate Akashiwo sanguinea on co-occurring phytoplankton: the significance of nutritional ecology*
- Investigation of the decline of Ulva prolifera in the Subei Shoal and Qingdao based on physiological changes*
- Effi ciency of phosphorus accumulation by plankton,periphyton developed on submerged artificial substrata and metaphyton: in-situ observation in two shallow ponds*
- Petroleum exploitation enriches the sulfonamide resistance gene sul2 in off shore sediments