
原代培养海马神经元成熟过程中PSD95与GluA2的分布
2021-06-15 04:17颜颖慧杨梦婕朱增燕
中国药理学通报 2021年6期
颜颖慧,杨梦婕,王 梅,杜 娆,朱增燕
(苏州大学附属儿童医院,江苏 苏州 215000)
中枢神经系统中的突触会经历突触强度的双向变化,这一过程被称为突触可塑性。这些变化在长时程增强(long-term potentiation,LTP)或长时程抑制(long-term depression,LTD)期间发生在个别突触的局部,统称为Hebbian可塑性[1]。目前普遍认为,通过Hebbian可塑性改变突触强度是学习和记忆的细胞基础,在可塑性过程中控制突触强度的一个主要机制是改变突触后膜上AMPA受体的数量、组成和生物物理性质[1]。
AMPA受体为离子型谷氨酸受体,介导快速兴奋性突触传递,正在成为诸多中枢神经系统药物研发的作用靶点[2],其有4种高度同源的亚基GluA1-4,由GluA1/2组成的四聚体是海马组织中最主要的AMPA受体亚型。GluA2决定了天然受体的生物物理属性,包含GluA2的离子通道是不通透Ca2+的。很多证据表明,在LTP初始阶段突触部位的Ca2+通透的AMPA受体(calcium permeable AMPARs,CP-AMPARs)是必需的,但在维持阶段则非必要。在记忆检索过程中,不通透Ca2+的AMPA受体(calcium impermeable AMPARs,CI-AMPARs)会被CP-AMPAR所替换[3];但未经调节的CP-AMPARs引起的Ca2+内流被认为是一些疾病中神经功能紊乱和死亡的基础[4-5]。可见,GluA2是否参与AMPA受体的构成与组装对受体功能发挥及突触可塑性进程至关重要。
突触后致密区(postsynaptic density,PSD)是兴奋性突触树突棘内致密的局部区域,可作为兴奋性突触的标记,其由突触可塑性相关的受体、激酶、结构蛋白和信号分子组成。而突触后密度蛋白-95(PSD95)是PSD最丰富的蛋白[6],它是膜相关鸟苷酸激酶 (membrane-associated guanylate kinase,MAGUK)家族的成员,是一种位于兴奋性突触的支架蛋白,参与N-甲基-D-天冬氨酸(N-methyl-D-aspartate,NMDA)受体和AMPA受体向突触后膜的稳定、募集和运输[6]。PSD95是神经发育过程中参与谷氨酸传递、突触可塑性和树突棘形态形成的重要成分[7]。
来自啮齿动物的原代神经元培养物被广泛用于研究神经元的基本生理特性。在体外神经元发育和成熟的过程中,研究兴奋性突触传递与突触可塑性的最佳培养时间如何确定;介导快速兴奋性突触传递的AMPA受体功能亚基GluA2的胞内分布以及与作为兴奋性突触标记的PSD95的定位之间是否存在联系,目前尚未见报道。因此,本研究旨在对这些问题做一个初步的探讨。
1 材料与方法
1.1 材料新生24 h Sprague-Dawley大鼠用于体外原代培养神经元,培养液采用含B27(17504044,Gibco,USA)和Neurobasal(21103049,Gibco,USA)的血清培养基。细胞免疫荧光实验中所用一抗为Anti-GluR2 (GluA2) (extracellular) Antibody(rabbit,alomone labs,Israel)和 PSD95 Antibody[7E3-1B8](mouse,abcam,USA),二抗为驴抗小鼠IgG H&L (Alexa Fluor®594)和驴抗兔IgG H&L (Alexa Fluor®488),均购买于Abcam公司。细胞免疫荧光实验的观察记录使用Olympus FV1000激光共聚焦显微镜(Olympus Microsystems,Japan)。
1.2 海马神经元原代培养新生24 h大鼠以酒精棉球消毒、断颈。取出半月形海马,仔细剥离血管膜和其他附带脑组织,加入含木瓜蛋白酶的消化液,37 ℃放置15 min,终止液终止消化,吹打15-20次使细胞分散。静置2 min,将上清移入离心管,1 200-1 500 r·min-1离心5 min。倾去上清,加入贴壁液将沉淀吹打松散混匀。显微镜下计数后将1×106细胞种于多聚赖氨酸包被的共聚焦小皿中,37 ℃,5% CO2培养。4-5 h后更换培养液仍为贴壁液,12 h后更换培养液为含B27的神经元培养液(含五氟尿嘧啶)。以后每隔5-7 d 1/4换液,维持五氟尿嘧啶的浓度。实验中涉及到动物的使用已经过苏州大学附属儿童医院伦理委员会审查通过。
1.3 细胞免疫荧光为测定GluA2与PSD95的共定位,原代培养海马神经元至4、7、14及20 d,在4%多聚甲醛中室温固定15 min,用PBS漂洗并以含0.3%Triton X-100及10%驴血清的PBS室温封闭通透1 h。在0.5%BSA中分别与兔抗GluA2抗体(1 ∶250)和小鼠抗PSD95抗体(1 ∶200) 4 ℃摇动过夜,然后分别与驴抗小鼠IgG H&L (Alexa Fluor®594)和驴抗兔IgG H&L (Alexa Fluor®488)室温孵育1 h。
1.4 荧光显微镜和数据分析图像由Olympus FV1000激光共聚焦显微镜以60倍的倍率拍摄。用ImageJ软件的Coloc功能定量分析GluA2和PSD95在原代海马神经元的树突部位共定位情况。Coloc函数通过比较两个标记的立体像素强度之间的相似性来提供Pearson相关系数。相似性越高,Pearson相关系数越高,则代表两种标记物的共定位程度越高[8]。Pearson相关系数>0.6被认为是强烈共定位。为了计算Pearson相关系数,创建了绿色标记的GluA2通道(通道A)用于定义感兴趣区域(ROI)。然后在COLOC模块中,获取通道A和红色标记的PSD95通道B的感兴趣区域的立体像素强度。对于≥3个条件的统计比较,在PRISM中使用了单因素方差分析和Tukey’s的后处理分析。
2 结果
2.1 在神经元培养的不同阶段,PSD95在神经元不同部位的表达培养至4、7、14及20 d的神经元,PSD95在神经元胞体、轴突、树突和树突棘显示出明显的分布差异(Fig 1):神经细胞核无PSD95表达,4DIV神经元PSD95分布较均一,胞体、轴突、树突均有表达;7DIV神经元胞体、轴突、树突均有PSD95表达,树突荧光较弱,其分布与神经元形态较一致,表达较为均一;14 DIV神经元已可见PSD95呈点状分布,树突密集处点状表达较多;20 DIV神经元PSD95在树突位置呈点状密集分布,且可见在树突棘的表达增多。

Fig 1 Cell morphology of primary hippocampal neurons (4DIV,7DIV,14DIV and 20DIV) and distribution of PSD95 and GluA2 (Scale bars,10 μm)
2.2 在神经元培养的不同阶段,胞内GluA2在神经元不同部位的表达培养至4 d、7 d、14 d及20 d的神经元,GluA2显示出在细胞内的均匀分布,其在神经元胞体、轴突、树突分布均一,20DIV神经元的树突棘也有GluA2表达,与神经元形态发生一致(Fig 1)。GluA2在细胞核无表达。
2.3 随神经细胞的成熟,PSD95与GluA2的共定位表达水平逐渐下降培养至4、7、14及20 d的神经元PSD95和GluA2在树突(棘)的共定位表达如Fig 2所示。通过Pearson相关系数对树突图像中的总GluA2/PSD95共定位进行量化,结果显示,4DIV和7DIV神经元树突部位Pearson系数分别为0.830±0.033和0.734±0.019,显示两种蛋白具有强烈的共定位;而14DIV和20DIV神经元树突部位Pearson相关系数则为0.547±0.021和0.574±0.024,呈现中等程度相关。4DIV神经元(n=21)树突共定位系数高于14DIV(P<0.01)和20DIV(P<0.01);7DIV神经元树突共定位系数高于14DIV(P<0.01)和20DIV(P<0.01)。
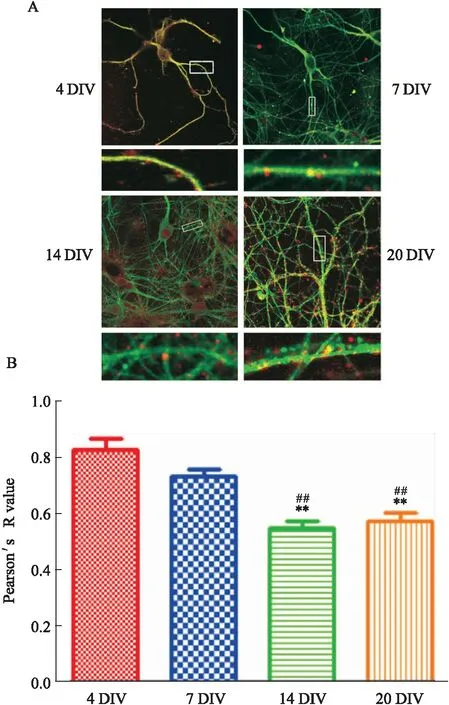
Fig 2 A. Colocalization of PSD95 and GluA2 on dendrites of different stages of primary hippocampal neurons;B. Pearson correlation of PSD95 and GluA2 on primary hippocampal neuron dendrites of different stages
3 讨论
3.1 PSD95的功能定位依赖于树突棘状结构的产生PSD95在成熟的神经细胞中主要表达于兴奋性突触后,为兴奋性突触上谷氨酸受体的聚集和稳定提供支架,在大鼠出生后早期发育过程中,棘突不稳定并且突触不成熟,此时PSD95的表达很低,只有在成年后才增加到最高水平[9]。表达PSD95荧光标记物的体外研究表明,大多数新的树突棘最初缺乏PSD95[10-11];在分离的神经元培养中,新生树突棘接收突触前信号传递的数分钟到数小时内,PSD95-GFP在树突棘中积累[10-12]。在这些研究中,已经证明PSD95-GFP的招募依赖于突触的活动[10-11]。体外培养海马神经元的成熟过程中,PSD95蛋白始终表达于神经元细胞内,其分布却不尽相同,且与神经元形态密切相关。4DIV 的神经元细胞形态单一、树突稀少,7DIV的神经元细胞可见较多无刺突树突,14DIV神经元细胞仍未见明显树突棘,至20DIV原代海马神经元细胞呈现出较多棘状结构,这与成年小鼠海马神经发生实验中对新生神经元进行的形态学分析结果相一致[13],与此相对应的,PSD95随着细胞成熟逐渐呈点状分布于树突及树突棘,棘状结构的产生为突触活动提供基础,PSD95在棘部的定位标志着成熟、稳定突触的存在。
3.2 GluA2与PSD95在树突棘的共定位提示该树突棘存在着包含AMPA受体的稳定突触成年小鼠海马的神经发生实验中对逆转录病毒标记的神经元进行的形态学分析结果表明,树突棘形成的起点时间为16 d[13],这也标志着谷氨酸能突触的发生,电生理结果也显示到第3周,新生神经元开始接受功能性谷氨酸能传入,并显示高频适应的重复动作电位。可见兴奋性谷氨酸受体的功能性表达依赖于神经元的成熟。那么作为谷氨酸受体支架的PSD95表达对AMPA受体与NMDA受体功能影响是否相同呢?已有研究表明,PSD95在切片培养中的过表达导致AMPA受体介导的传递增加,并且可以模仿和阻断LTP[14-15]。在体外培养神经元中敲除PSD95会导致AMPAR介导的兴奋性突触后电流减少,并阻止具有功能性AMPA受体突触的正常发育[16]。在PSD95基因敲除的小鼠中,兴奋性突触后电流的AMPA/NMDA介导比率也降低,这表明PSD95在兴奋性突触群体的成熟过程中发挥了作用,且在AMPA受体的功能发挥中扮演重要角色[17-18]。
本研究发现在体外培养原代海马神经元过程中,20DIV原代海马神经元细胞呈现出较多棘状结构,提示体外培养细胞表现为成熟神经元状态;随着神经元的成熟,PSD95与GluA2在树突的共定位水平下降,尤其20DIV神经元树突部位GluA2与PSD95仅呈现中等程度共定位。研究结果提示,在树突棘结构出现之前,GluA2与PSD95在神经元细胞的共定位意义不大,谷氨酸受体作为神经元的固有属性最初即表达于神经元细胞内,而PSD95依赖于突触活动的招募方能发挥功能,两种蛋白在神经元棘状结构产生后才依赖于突触活动发生有意义的功能定位。同时并非所有表达AMPA受体的棘状结构均表达PSD95,缺乏PSD95不一定能否定突触的存在,而是表明缺乏包含稳定的AMPA受体的成熟突触。因此,体外培养神经元至少应在16-19 d以上才视为成熟的神经元,能够开展突触可塑性等相关功能性研究。
综上所述,GluA2蛋白在原代海马神经元成熟过程中胞内分布无特异性,PSD95的胞内功能定位依赖于树突棘状结构的产生;而GluA2与PSD95在成熟神经元树突棘的共定位提示在该树突棘部位存在着包含AMPA受体的稳定突触。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
河南大学学报(医学版)(2021年1期)2021-11-26
海外星云(2021年6期)2021-10-14
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
教育教学论坛(2019年18期)2019-06-17
中西医结合心血管病电子杂志(2018年28期)2018-11-19
中国现代医学杂志(2015年26期)2015-12-23
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年3期)2015-06-10
