
基于叶绿体DNA的黄土高原特有植物蕤核的谱系地理学
2021-06-10 21:59刘家奇付孟娇吴海洋田斌
广西植物 2021年3期
关键词:黄土高原
刘家奇 付孟娇 吴海洋 田斌
摘 要:该文利用母系遗传的叶绿体DNA片段(psbA-trnH,psbI-psbK和psbJ-petA)对黄土高原地区的特有植物蕤核(Prinsepia uniflora)进行谱系地理学研究,以揭示其现有的遗传结构和群体历史动态。结果表明:(1)蕤核自然种群总的遗传多样性较高(HT
=0.796),而种群内的遗传多样性较低(HS =0.276)。(2)分子变异分析(AMOVA)表明,分布区域内蕤核具有较高的遗传分化(FST = 0.674),且遗传变异主要存在于种群间(67.4%)。(3)Mismatch分析和基于MaxEnt的不同历史时期生态位模拟结果表明,尽管蕤核的核心适生范围没有经历太大的变化,但其种群在最后一次大冰期结束后仍存在一定程度上的扩张。综上结果认为,与大多数分布在我国北方地区的许多植物类群一样,蕤核很可能是采取冰期在邻近或者就地避难,冰期过后局部回迁的方式应对第四纪的气候波动。
关键词: 蕤核, 黄土高原, 叶绿体DNA, 谱系地理学
中图分类号:Q948.2
文獻标识码:A
文章编号:1000-3142(2021)03-0396-08
收稿日期:2019-12-16
基金项目:国家自然科学基金(41861008) [Supported by National Natural Science Foundation of China(41861008)] 。
作者简介: 刘家奇(1994-),硕士研究生,研究方向为林木遗传育种、植物群体遗传,(E-mail)2047223903@qq.com。
通信作者:田斌,博士,副教授,研究方向为植物学、林木遗传育种、植物群体遗传学,(E-mail)tianbin@swfu.edu.cn。
Phylogeography of the Loess Plateau endemic plant Prinsepia uniflora inferred from the chloroplast DNA
LIU Jiaqi, FU Mengjiao, WU Haiyang, TIAN Bin
( Key Laboratory of National Forestry and Grassland Administration on Biodiversity Conservation in Southwest China (Southwest Forestry University), Kunming 650224, China )
Abstract:To reveal the historical dynamics of the population and existing genetic structure, maternal chloroplast DNA fragments (psbA-trnH, psbI-psbK and psbJ-petA) were used to study the phylogeography of Prinsepia uniflora, a kind of endemic plant distributed in the Loess Plateau. The results were as follows: (1) Total genetic diversity of P. uniflora was high (HT = 0.796), but genetic diversity within species was low (HS= 0.276); (2) The molecular variation analysis (AMOVA) showed that there was a high genetic differentiation within P. uniflora (FST = 0.674), and the genetic variation was mainly between populations (67.4%); (3) Mismatch analysis and MaxEnt based ecological niche simulation results in different historical periods showed that although the core suitable range of P. uniflora did not change much, population still expanded to some extent after the end of the Last Glacial Maximum (LGM). Our study indicated that, like most of the plant groups distributed in the northern regions of China, P. uniflora probably responded to the Quaternary climatic fluctuations by taking refuge nearby or in situ during the glacial period and locally recolonizing in following post-glacial period.
Key words: Prinsepia uniflora, Loess Plateau, chloroplast DNA, phylogeography
伴随地质历史事件的生物类群群体历史动态变化对理解现今的生物分布和多样性格局,以及预测未来气候变化条件下的物种响应有着非常重要的意义(Davis et al.,2005)。第四纪(约200万年前)以来,全球范围内出现了多次气候的周期性波动,其典型事件是冰期和间冰期交替出现,对植物类群现今的地理分布和种群遗传格局造成了深刻影响(Comes & Kadereit,1998;Provan & Bennett,2008)。
越来越多的证据表明,尽管以黄土高原为代表的中国北部没有像欧洲和北美那样大规模冰川发育的自然地理条件,但是第四纪气候的反复变化,也对分布在这一地区的植物地理分布格局和群体遗传结构产生了一定影响(Qiu et al.,2011;Liu et al.,2012)。其具体表现为种群内遗传多样性降低,以及分布范围的反复扩张、收缩或生境破碎化。并且多数研究表明中国北方现存的植物物种在冰期存在多个避难所,冰期结束后也仅仅是在局部范围内回迁(Chen et al.,2008;Tian et al.,2009;Fu et al.,2014;Zeng et al.,2015)。
蕤核(Prinsepia uniflora)隶属于蔷薇科李亚科扁核木属,为中国的特有植物类群。在山西、内蒙古、陕西、甘肃、宁夏等黄土高原地区广泛分布。该物种主要生于山坡、河谷等稀疏灌丛或干旱沙丘,海拔 900~1 100 m(Gu & Bartholomew,2003)。由于该物種具有一定的药用和食用价值,加之黄土高原地区频繁人类活动的影响,使其生存受到了严重的威胁。我们的野外调查发现,目前该物种自然群体片段化分布,并且种群内的植株数量非常有限。本研究拟利用3套叶绿体DNA片段去构建黄土高原特有种蕤核的谱系地理结构,结合生态位模拟的方法,试图为黄土高原地区植被的历史动态提供线索,并对该物种的有效保护提供理论支持。
1 材料与方法
1.1 材料
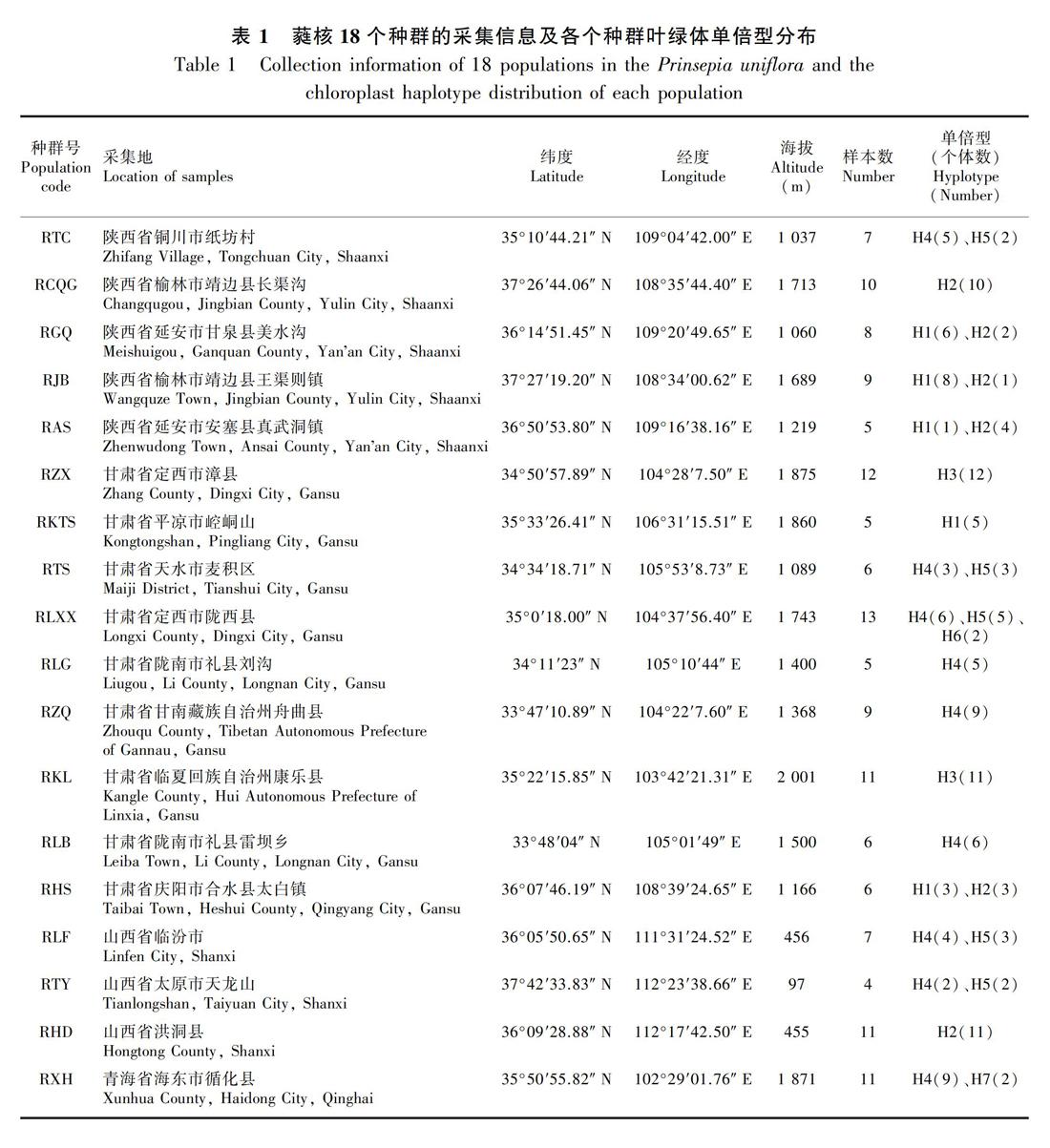
研究材料于2010—2017年采自陕西、甘肃、山西、青海,共采集蕤核18个种群144个个体(表1,图1)。每个种群随机采集4~13个个体,个体之间相隔至少100 m以上。采集生长良好植株的新鲜嫩叶后立即用硅胶干燥,带回实验室存放于-20 ℃冰箱备用。凭证标本贮藏于西南林业大学标本馆。
1.2 DNA提取与PCR扩增
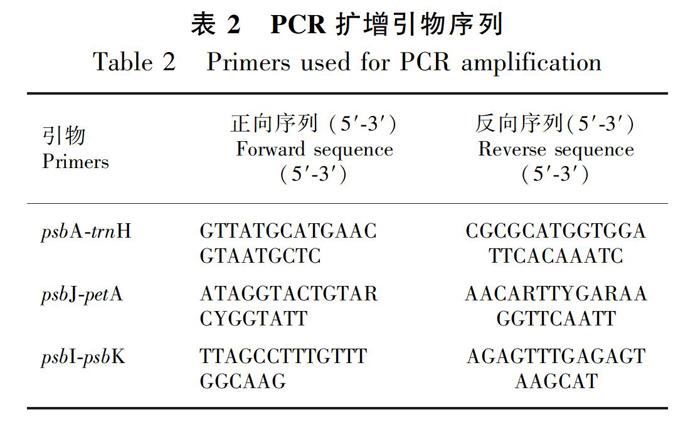
采用改良的CTAB法(Doyle JJ & Doyle JL,1990)提取所有采集样本叶片的总DNA。从候选的10对cpDNA通用引物筛选出具有多态性的引物,最终甄选出3对cpDNA引物(psbA-trnH、psbJ-petA、trnL-trnF)进行PCR扩增,引物序列见表2。PCR扩增反应体系为25 μL,内含2Taq MasterMix(Sangon Biotech)12.5 μL、正反向引物各1 μL(浓度为5 μmol·L-1)、植物总DNA模板1 μL以及双蒸水9.5 μL。PCR扩增反应程序为94 ℃预变性4 min;94 ℃变性30 s,52 ℃退火45 s,72 ℃延伸60 s,循环35次;72 ℃延伸7 min结束,4 ℃保存。多次实验筛选出3对引物最佳的退火温度均为52 ℃。PCR产物用1%琼脂糖凝胶电泳检测后送测序公司进行双向测序。
1.3 数据分析
经测序后得到的数据使用ClustalX 1.83 (Thompson et al.,1997)以及MEGA 7.0.26 (Kumar et al.,2016)进行序列的多重比对,同时将原始峰图作为参照进行手工校对。所有的序列完全校对正确后,再用DnaSP 5.10 (Librado & Rozas,2009)程序来统计叶绿体DNA单倍型。
叶绿体DNA单倍型的关系通过NETWORK 4.1.6.3 (Bandelt et al.,1999)软件来检测。我们选择最大简约法以最小生成树为基础构建单倍型中央连接网状图。在分析过程中点突变和缺失被平等处理,并且假设每个突变之间是相对独立的。种群内遗传多样性(HS)、总遗传多样性(HT)和物种层面上的种群分化水平GST和NST值均使用PERMUT (Pons & Petit,1996)软件来估测和观察叶绿体的变异情况。GST值计算只使用了单倍型频率,而NST值则把单倍型之间的差异也计算在内。比较这两个值可以看出单倍型变异在整个地理分布区的地理格局,如果NST值大于GST值,且达到显著水平(P < 0.05),则表明关系相近的单倍型在相同的种群出现比在不同种群出现几率大,说明存在着明显的谱系地理结构。采用Arlequin 3.5.2 (Weir & Cockerham,1984)软件的分子变异分析(analysis of molecular variance,AMOVA)对种群间和种群内的遗传差异进行估算,其中显著性检验使用随机参数进行1 000次的置换检验。
1.4 生态位模拟
本研究通过最大熵模型(Phillips et al.,2006) (Maximum entropy;Maxent)对蕤核进行生态位模型预测来识别该物种在第四纪时的避难所和避难模式,推测大冰期后至今的历史动态,以及未来在全球变暖的趋势下该物种的潜在分布。蕤核的当前分布数据主要来自我们的野外调查采样以及植物标本馆记录(www.cvh.ac.cn)。在去除重复的分布点后,得到35个当前的分布位置。基于4个时期(Last Interglacial,LIG、Last Glacial Maximum,LGM、Present and Future)背景下的各自19个气候变量从WorldClim数据库下载(www.worldclim.org),分辨率和大气环流模型分别统一为2.5 arc-min (大约5 km)和CCSM4。根据Pearsons相关系数(r>0.9),对当前19个气候变量中具有强相关性的变量排除,剩下的9个气候变量用于模型创建。生态位模型创建在Maxent 3.3.3中进行,设置Random test percentage为25%,Replicates为10。
2 结果与分析
2.1 蕤核的单倍型分布
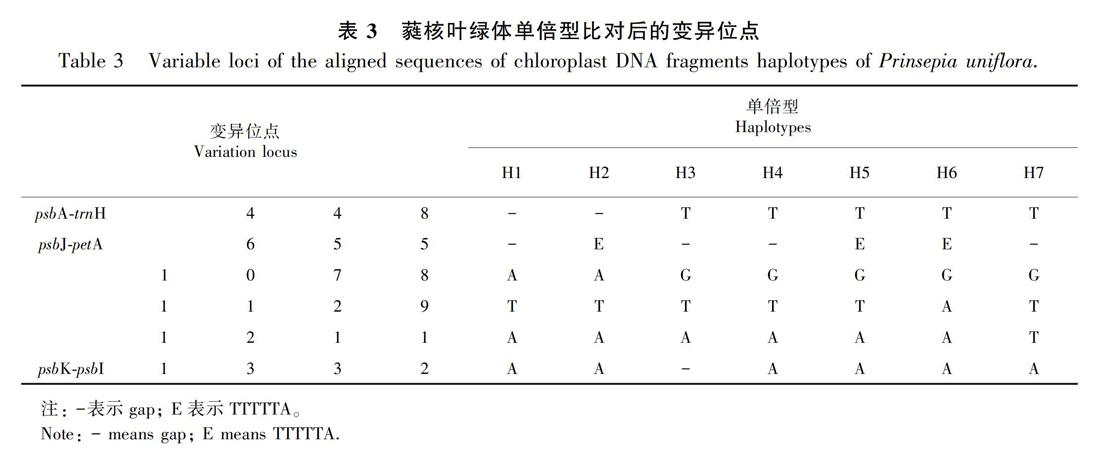
对蕤核cpDNA的3个片段psbA-trnH、psbJ-petA和psbK-psbI进行扩增测序的结果显示:3个片段各出现1个、4个、1个变异位点(表3)。经过序列比对后的psbA-trnH、psbJ-petA和psbK-psbI 3对引物片段长度分别为495、801、244 bp,这3套叶绿体DNA片段的序列拼接在一起可得到1 540 bp的长度,一共识别出7种叶绿体单倍型H(1-7),存在5个点突变和4处插入缺失。
从蕤核叶绿体单倍型的分布图(图1)可以看出,共计8个种群只存在1种单倍型,另外有9个种群有2种单倍型,仅在1个种群中存在3种单倍型RLXX(H4、H5、H6)。单倍型H4出现的频率最高。单倍型网状图显示蕤核的单倍型成多元扩张分布,主要是以单倍型H4和H5为中心,由单倍型H4和H5向四周发散,然后衍生出单倍型H2、 H3这2个大的分布类型,并且多数单倍型之间仅有一步的变异(图1)。
2.2 蕤核遗传多样性和遗传结构分析
从表4可以看出,蕤核种群内遗传变异百分比为32.59%,种群间遗传变异量占总变异量的67.41%,种群间发生显著的遗传分化,FST=0.674 12(表4,P<0.001)。通过PREMUT程序计算得出其自然种群间遗传多样性HT(0.796)高于种群内的遗传多样性HS(0.276)。从所有的18个种群地理单元间遗传变异性和分化系数显示出NST(0.834)显著高于GST(0.654)(P<0.01)。结果显示,蕤核叶绿体单倍型有明显的谱系地理学结构。
2.3 蕤核种群历史分析
叶绿体单倍型的种群错配的分布显示单峰型(图2),表明了蕤核种群经历了种群的急速扩张性事件。基于计算所得的τ值为0.516,根据τ=2ut计算扩张经历世代数t,其中u=2μkg。通过野外观察,我们设定世代长度g为3 a,μ代表DNA平均变异速率,采用木本植物的进化速率,取已发表的平均变异速率的较高值:3~9×10-9变异/位点/年(substitution/site/year)。k表示序列总长度为1 540。计算得到扩张时间为18 614~8 272年前。
2.4 生态位模拟分析
生态位模型创建结果获得了非常高的AUC(the Area Under the Curve)值(> 0.995)的支持。蕤核4个时期的潜在分布预测图中清晰地展现了该物种在不同气候背景下的分布格局。与现在(Present)的分布范围相比,LIG和LGM时种群范围并没有出现较大的变化,其分布的核心区均在黄土高原,只是在最后一次冰期后出现了小幅度的适生区扩大。然而在未来(2070年)蕤核的分布范围将会缩小(图3)。
气候因子Bio6(最冷月最低温)和Bio15(降水季节变化)在生态位建模的4个时期,都表现出较高的贡献率,尤其是LIG,Bio15的贡献百分比达到了37.4%(图4)。此外,Bio16(最湿季节降水量)、Bio13(最湿月降水量)和Bio14(最干月降水量)在现在和未来(Future)蕤核分布区模拟中占有较高的贡献百分比(图4)。
3 讨论与结论
本研究发现蕤核具有明显的谱系地理结构(NST = 0.834 >GST = 0.654, P < 0.01),并且有显著的遗传分化(FST=0.674 12, P<0.001)和较低的种群内遗传多样性(HS=0.276)。分子变異遗传方差分析(AMOVA)的结果显示,蕤核遗传变异主要以种群间的变异为主,占67.41%。总的来说,植物种群的遗传结构受到地质历史气候变迁、环境因素(如地势条件、生境异质性)以及种子基因交流程度等诸多因素的影响。已有的研究表明,我国北方分布的植物类群由于第四纪冰期与间冰期的反复交替造成不同避难所之间的种群产生强烈的遗传分化,并且降低了种群内的遗传多样性(Qian & Ricklefs,2000;Chen et al.,2008;Qiu et al.,2011;Zeng et al.,2015)。此外,我们在野外采样过程中发现,蕤核现存的自然种群大多零星分布在黄土高原的中高海拔地区,种群规模相对较小。黄土高原沟壑纵横的地貌和严重的水土流失可能加剧了蕤核不同种群之间的基因交流困难性,使得种群之间具有较大的遗传分化。黄土高原地区长期而剧烈的人类活动可能是造成上述结果的另一重要原因。
从蕤核单倍型的network分析和地理分布来看,两种古老单倍型H4和H5从东到西均有分布,其他衍生单倍型则是局限地分布于不同的地理区域,如单倍型H1、H3和H7仅分布在分布区西部的少数几个种群。生态位模拟的结果也表明,在最后一次大冰期蕤核并没有大规模的分布区范围变化。Mismatch分析(错配分析)表明蕤核种群经历了近期的扩张事件。经推算的扩张时间为1.86万~0.82万年前,这与第四纪最后一次大冰期结束的时间(1.8万~1.6万年前)相吻合。因此,我们推测该物种在最后一次大冰期时,并没有大规模的灭绝事件发生,而是在多个冰期避难所就地避难。不同地区避难的种群也很可能没有经历大范围的扩张,仅仅回迁到邻近地区。这一推测也和在该地区分布的其他重要树种如虎榛子(Ostryopsis davidiana)(Tian et al.,2009)、 连翘(Forsythia suspensa) (Fu et al.,2014)、文冠果(Xanthoceras sorbifolium)(Zhu et al.,2019)等的研究结果相类似。本研究同样支持了我国北方地区的许多植物类群并没有像同纬度欧洲分部的植物一样出现大规模的南迁避难,而是多采取邻近或者就地避难的方式应对第四纪的气候波动。
全球气候变化下,很多生态敏感地区的动植物都有遗传多样性丧失甚至灭绝的风险(Vellend & Geber,2005)。对于黄土高原这一生态脆弱地区来说,分布在这一地区的特有动植物类群应该备受关注。我们对蕤核生态位模拟的结果显示,不同的历史时期,降水相关的生态因子都表现出较高的贡献率。在未来全球气候将会出现的增温变干的趋势下,蕤核适生范围有可能急剧减小。加之其生存的黄土高原地区特殊的生境和强烈的人为干扰,这一黄土高原特有植物类群将面临灭绝风险。结合蕤核种群遗传结构结果,该物种具有较高的遗传分化水平,并且多个种群均出现了独有的叶绿体单倍型。因此,我们认为蕤核的多数种群均有保护价值,一方面应对现有的蕤核群体进行有效的保护,另一方面应加大对蕤核遗传资源的收集,进行入库保存。
參考文献:
BANDELT HJ, FORSTER P, ROHL, A, 1999. Median-joining networks for inferring intraspecific phylogenies [J]. Mol Biol Evol, 16(1): 37-48.
CHEN K, ABBOTT RJ,MILNE RI, et al., 2008. Phylogeography of Pinus tabulaeformis Carr. (Pinaceae), a dominant species of coniferous forest in northern China [J]. Mol Ecol, 17(19): 4276-4288.
COMES HP, KADEREIT JW, 1998. The effect of quaternary climatic changes on plant distribution and evolution [J]. Trends Plant Sci, 3(11): 432-438.
DAVIS MB, SHAW RG, ETTERSON JR, 2005. Evolutionary responses to changing climate [J]. Ecology, 86(7):1704-1714.
DOYLE JJ, DOYLE JL, 1990. A rapid total DNA preparation procedure for fresh plant tissue [J]. Focus, 12:13-15.
FU ZZ, LI YH, ZHANG KM, et al., 2014. Molecular data and ecological niche modeling reveal population dynamics of widespread shrub Forsythia suspensa (Oleaceae) in Chinas warm-temperate zone in response to climate change during the Pleistocene [J]. Bmc Evol Biol, 14(1): 1-13.
GU CZ, BARTHOLOMEW B, 2003. Prinsepia Royle [M]//WU ZY, RAVEN PH. Flora of China. Beijing: Science Press, 9; St. Louis: Missouri Botanical Garden Press: 389-391.
KUMAR S, STECHER G, TAMURA K, 2016. MEGA7:molecular evolutionary genetics analysis version 7.0 for bigger datasets [J]. Mol Biol Evol, 33(7): 1870-1874.
LIBRADO P, ROZAS J, 2009. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data [J]. Bioinformatics, 25(11): 1451-1452.
LIU JQ, SUN YS, GE XJ, et al., 2012. Phylogeographic studies of plants in China: advances in the past and directions in the future [J]. J Syst Evol, 50(4): 267-275.
PHILLIPS SJ,ANDERSON RP, SCHAPIRE RE, 2006. Maximum entropy modeling of species geographic distributions [J]. Ecol Model, 190(3-4): 231-259.
PONS O, PETIT RJ, 1996. Measuring and testing genetic differentiation with ordered versus unordered alleles [J]. Genetics, 144(3): 1237-1245.
PROVAN J, BENNETT KD, 2008. Phylogeographic insights into cryptic glacial refugia [J]. Trend Ecol Evol, 23(10): 564-571.
QIAN H, RICKLEFS RE, 2000. Large-scale processes and the Asian bias in species diversity of temperate plants [J]. Nature, 407(6801): 180-182.
QIU YX, FU CX, COMES HP, 2011. Plant molecular phylogeography inChina and adjacent regions: Tracing the genetic imprints of quaternary climate and environmental change in the worlds most diverse temperate flora [J]. Mol Phylogenet Evol, 59(1): 225-244.
THOMPSON JD, GIBSON TJ, PLEWNIAK F, et al., 1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools [J]. Nucl Acid Res, 25(24): 4876-4882.
TIAN B, LIU R, WANG L, et al., 2009. Phylogeographic analyses suggest that a deciduous species (Ostryopsis davidiana Decne., Betulaceae) survived in northern China during the Last Glacial Maximum [J]. J Biogeogr, 36(11): 2148-2155.
VELLEND M, GEBER MA, 2005. Connections between species diversity and genetic diversity [J]. Ecol Lett, 8(7):767-781.
WEIR BS, COCKERHAM CC, 1984. Estimating F-statistics for the analysis of population-structure [J]. Evolution, 38(6): 1358-1370.
ZENG YF, WANG WT, LIAO WJ, et al., 2015. Multiple glacial refugia for cool-temperate deciduous trees in northern East Asia: the Mongolian oak as a case study [J]. Mol Ecol, 24(22): 5676-5691.
ZHU RB, WANG Q, GUAN WB, et al., 2019. Conservation of genetic diversity hotspots of the high-valued relic yellowhorn (Xanthoceras sorbifolium) considering climate change predictions [J]. Ecol Evol, 9(6): 3251-3263.
(責任编辑 何永艳)
猜你喜欢
炎黄地理(2020年3期)2020-05-26
科学与财富(2017年24期)2017-09-06
山东工业技术(2017年16期)2017-09-05
新课程·下旬(2017年6期)2017-08-05
科教导刊·电子版(2016年35期)2017-04-20
绿色科技(2017年1期)2017-03-01
中外文摘(2016年7期)2016-12-20
江苏农业科学(2014年12期)2015-04-02
吉林农业·下半月(2014年10期)2014-11-06
雕塑(1998年2期)1998-06-28
