
tae-miR167d及靶基因ARF12在小麦中的表达模式分析
2021-06-09 03:45:14宋国琦李玮张淑娟张荣志李玉莲高洁李吉虎陈明丽李根英
山东农业科学 2021年5期
宋国琦,李玮,张淑娟,张荣志,李玉莲,高洁,李吉虎,陈明丽,李根英
(山东省农业科学院作物研究所/农业部黄淮北部小麦生物学与遗传育种重点实验室/小麦玉米国家工程实验室,山东 济南 250100)
小麦(Triticum aestivurn L.)是世界上最主要的粮食作物之一,在粮食安全中占有举足轻重的地位。低温是影响作物生长发育的重要环境因子之一。近来,许多研究阐述了植物miRNAs在冷胁迫响应中的作用。拟南芥中发现16个miRNAs参与冷胁迫响应[1-3]。在杨树中鉴定出19个冷胁迫响应miRNAs[4],其中6个与拟南芥中相同。Zhang等[5]在短柄草中发现了25个冷胁迫响应的特异性miRNAs。这些研究表明冷响应miRNAs既具有保守性又具有调控特异性。在普通小麦中,许多保守的、小麦属特异的及小麦中特异的miRNAs被鉴定出来,包括小麦A基因组的祖先、B基因组的祖先及AABBDD异源六倍体小麦[6-11]。小麦中许多miRNAs在胁迫响应中发挥着 重 要 的 作 用[8,12,13]。 在 冷 胁 迫 响 应 中,miR167c、tae-miR167d、tae-miR172a、tae-miR393、tae-miR396a和tae-miR444c.1这6个miRNAs在小麦温敏核不育系(TGMS)中响应冷胁迫[14];miR159、miR164、miR169、miR319、miR398、miR1029和miR1126这7个miRNAs在小麦苗期响应冷胁迫[8]。分析小麦中应答不同胁迫反应的miRNAs发现,对于多样性的生物胁迫和非生物胁迫,miRNAs表现出交叉应答反应,这表明小麦已经发展了复杂的miRNA介导途径应对不断变化的环境[15]。因此,研究小麦幼穗中miRNAs的冷胁迫响应机制,有助于在miRNAs层面上理解小麦生殖阶段的冷响应机制,为提高小麦春季抗冻性提供理论依据。
本课题组对处于药隔期的济麦22幼穗低温胁迫后进行了miRNA分析和降解组分析。共发现来自105个家族的192个保守miRNAs和9个新miRNAs,其中34个保守miRNAs和5个新miRNAs在冷胁迫前后表达差异显著。对这些miRNAs的靶基因预测和降解组验证表明,差异表达miRNAs的靶基因多数具有应答刺激、调控转录和离子运输的功能。其中有13个miRNAs的靶基因参与花发育过程,如ARF(auxin response factor),SPB(squamosa promoter binding like protein)和AP2(AP2-like ethylene-responsive transcription factor)等,其中miR160和miR167的靶基因均为ARF。前期结果显示,tae-miR167d在药隔期的幼穗中低温诱导后下调表达,而其靶基因ARF12则上调表达[15]。这一结果与Tang等[14]在小麦冷诱导温敏不育系过程中的发现一致,他们推测tae-miR167d和靶基因ARF在小麦温敏核不育系的穗发育中可能控制冷诱导的雄性不育。另有研究表明,miR167家族参与调控植物花和果实的发育[16-19],而其靶基因ARF则是一类响应生长素信号的转录因子,通过特异性的结合生长素初期响应基因启动子区域AuxRE元件(TGTCNC)调控生长素响应基因的表达,从而引发生长素介导的许多生理效应[20-22]。
我们前期的研究结果发现tae-miR167d和靶基因ARF12在小麦生殖发育阶段响应冷胁迫[15],但其在小麦中的时空表达情况仍不清楚。本研究通过分析tae-miR167d及其靶基因ARF12在小麦不同发育时期的表达模式和组织表达特异性,以及冷胁迫下不同小麦品种tae-miR167d和ARF12的表达水平,探讨了tae-miR167d及其靶基因ARF12在小麦中的时空表达模式及冷胁迫后二者的调控关系。
1 材料与方法
1.1 供试材料
供试材料为济麦22、临麦4号、京411、豫麦47、鲁原502、周麦22、西农889和濮麦053。将小麦种子浸水至萌发后,置于4℃冰箱春化30 d,将芽苗修剪后移栽至直径20 cm的花盆中,花盆基质为草炭土,每盆种植6株。济麦22和西农889每个材料种植21盆,其余6个材料每个种植9盆。在人工气候室的可控环境下生长,设置昼/夜温度为22℃/16℃,光/暗周期为16 h/8 h,光照强度300μmol·m-2·s-1,相对湿度为60%~70%。
1.2 试验方法
1.2.1 取样方法 济麦22和西农889每个时期取生长发育一致的材料9株,每3株为一个重复,共3个重复。分别取三叶期、二棱期(7叶)的叶片,取雌雄蕊分化期(拔节)、四分体期(挑旗)和开花期的叶片、叶鞘、茎和幼穗,用锡箔纸包好,放入液氮中速冻,-80℃冰箱保存备用。
小麦品种济麦22、西农889、鲁原502、临麦4号、周麦22、豫麦47、濮麦053和京411每6株为1个重复,共3个重复。在雌雄蕊分化期进行0℃处理48 h,分别取正常生长和0℃处理48 h的叶片和幼穗,用锡箔纸包好,放入液氮中速冻,-80℃冰箱保存备用。
1.2.2 小麦RNA提取及cDNA合成 分别取0.1 g样品于液氮研磨后,利用miRcute多糖多酚植物miRNA提取分离试剂盒(天根,北京)进行总RNA的提取,利用Nanodrop 2000/2000C分光光度计(Thermo,美国)进行RNA浓度的测定。利用Mir-X miRNA First-Strand Synthesis Kit(Clontech,美国)进行反转录用于tae-miR167d的qRT-PCR分析;利用PrimeScript RT Reagent Kit With gDNA Eraser(TaKaRa,大连)进行反转录用于ARF12的qRT-PCR分析。
1.2.3 荧光定量PCR tae-miR167d定量PCR引物为tae-miR167d-F和miRNA-specific Primer,内参为U6 Forward Primer和U6 Reverse Primer。tae-miR167d-F引物序列如下:CTGAAGCTGCCAGCATGATCTA,miRNA-specific Primer、U6 Forward Primer和U6 Reverse Primer为Mir-X miRNA First-Strand Synthesis Kit(Clontech,美国)提供。qRTPCR按 照Light Cycler 480 SYBR GreenⅠ(Roche,德国)说明书进行反应体系配制和反应程序设定,在Light Cycler 480实时定量PCR仪(Roche,德国)上进行反应,反应程序为:95℃预变性5 min;95℃5 s,60℃20 s,72℃20 s,共40个循环。
ARF12定量PCR引物为ARF12-F/R,以小麦Actin为内参,引物序列如下:ARF12-F/R,ATAGATCAGGCTGGCAGCTTGT/GCACATCCTCTGGTGAAAGTATCT;TaActin-F/R,GCCACACTGTTCCAATCTATGA/TGATGGAATTGTATGTCGCTTC(AB181991)。反应程序为:95℃预变性5 min;95℃变性5 s,56℃20 s,72℃20 s,共40个循环。
反应结束后分析荧光值变化曲线和熔解曲线。每个反应3个技术重复,采用2-ΔΔCt法分析试验结果。
2 结果与分析
2.1 tae-miR167d和ARF12在小麦相同发育时期不同器官中的表达分析
分析济麦22和西农889中相同发育时期叶片、叶鞘、茎和穗中tae-miR167d及靶基因ARF12的表达情况(图1),发现在雌雄蕊发育时期以叶片为参照,济麦22叶鞘和茎中tae-miR167d表达上升,幼穗中tae-miR167d表达下降,而叶鞘中ARF12表达下调,茎和幼穗中ARF12表达上调;西农889叶鞘、茎和幼穗中tae-miR167d表达均下降,ARF12表达均上升。在挑旗期和开花期以叶片为参照,济麦22和西农889叶鞘、茎和幼穗中tae-miR167d表达下调,靶基因ARF12的表达上调,二者呈负调控关系。比较济麦22和西农889中tae-miR167d和ARF12表达差异发现,在雌雄蕊发育时期,济麦22的叶鞘和茎中tae-miR167d的表达高于ARF12,而西农889中正好与济麦22相反。

图1 tae-miR167d及靶基因ARF12在济麦22(A)和西农889(B)相同时期不同器官中的表达分析
2.2 tae-miR167d和ARF12在小麦不同发育时期相同器官中的表达分析
分析济麦22和西农889不同发育时期叶片、叶鞘、茎和穗中tae-miR167d及靶基因ARF12的表达情况(图2),发现随着生育进程推进,与三叶期叶片相比,济麦22叶片中tae-miR167d表达下调,靶基因ARF12表达上调;在西农889叶片中,除开花期tae-miR167d表达上调,其他3个时期均下调,而靶基因ARF12表达上调。与雌雄蕊发育时期的叶鞘相比,济麦22挑旗期和开花期叶鞘中的tae-miR167d的表达下调,靶基因ARF12的表达上调;在西农889中,挑旗期叶鞘中的taemiR167d的表达下调,靶基因ARF12的表达上调,而开花期叶鞘中tae-miR167d和ARF12均下调。与雌雄蕊发育时期相比,济麦22在挑旗期和开花期的茎和幼穗中tae-miR167d的表达均下降,靶基因ARF12的表达均上升;西农889挑旗期茎中tae-miR167d的表达上调,靶基因ARF12的表达下调,在开花期茎中tae-miR167d和靶基因ARF12的表达均下降;西农889幼穗中挑旗和开花期tae-miR167d和靶基因ARF12的表达也呈负调控关系。比较济麦22和西农889相同时期不同器官中tae-miR167d和靶基因ARF12表达差异,发现在叶鞘中两个品种差异明显。
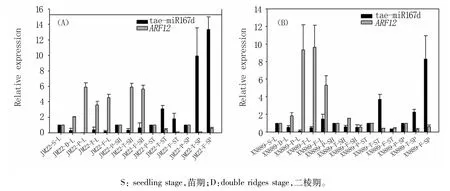
图2 tae-miR167d及靶基因ARF12在济麦22(A)和西农889(B)不同时期相同器官中的表达分析
2.3 冷胁迫前后不同小麦品种tae-miR167d及ARF12的表达分析
每个小麦品种冷胁迫后分别与自己的对照相比较,分析耐倒春寒不同8个小麦品种中taemiR167d及靶基因ARF12冷胁迫后的表达模式,结果(图3)发现,在叶片中,济麦22、京411、豫麦47、鲁原502和西农889冷胁迫后tae-miR167d表达下调而ARF12上调,临麦4号tae-miR167d上调而ARF12下调,在上述6个品种中,taemiR167d和ARF12二者呈负调控关系;周麦22和濮麦053中tae-miR167d和ARF12同时上调表达。在幼穗中,济麦22、京411、豫麦47、鲁原502、周麦22和西农889冷胁后tae-miR167d表达下调而ARF12上调,濮麦053中tae-miR167d上调而ARF12下调,在这7个品种中tae-miR167d和ARF12二者呈负调控关系;临麦4号taemiR167d和ARF12存在同时上调表达情况。通过分析发现冷胁迫前后tae-miR167d和ARF12的表达在多数品种中存在负调控关系。

图3 不同小麦品种冷胁迫后叶片和幼穗中tae-miR167d和ARF12相对表达量
与济麦22正常生长条件下的叶片相比较(图4A),正常生长条件下,除鲁原502中taemiR167d的表达量比济麦22高,其余6个品种tae-miR167d的表达量均比济麦22低;而ARF12的表达量除在京411、鲁原502和周麦22中比济麦22低,其余4个品种均比济麦22高。冷胁迫后,tae-miR167d的表达量在临麦4号、鲁原502和濮麦053中比济麦22中高,在其余4个品种中比济麦22的表达量低;而ARF12的表达量除在周麦22和西农889中比济麦22高,在其余5个品种中均比济麦22低。与济麦22正常生长条件下的幼穗相比较(图4B),发现正常生长条件下,7个品种中tae-miR167d和ARF12的表达量均比济麦22中高。冷胁迫后,tae-miR167d的表达量除在京411中比济麦22中低,在其余6个品种中均比济麦22中高;而ARF12的表达量除在鲁原502、周麦22和濮麦053中比济麦22中低,在其余4个品种中均比济麦22中高。比较叶片和幼穗中tae-miR167d和ARF12的表达趋势发现,多数情况下,在叶片中tae-miR167d的表达量低于ARF12,而幼穗中tae-miR167d的表达量高于ARF12。

图4 相同对照下不同品种小麦叶片(A)和幼穗中(B)tae-miR167d和ARF12的相对表达量
3 讨论与结论
本课题组在研究倒春寒胁迫响应miRNAs时,发现小麦幼穗中tae-miR167d参与冷响应,并发现其靶基因为ARF12。qRT-PCR结果显示,小麦幼穗中tae-miR167d在低温诱导后下调表达,而其靶基因ARF12则上调表达[15]。本研究的目的是验证上述结果,并明确耐倒春寒品种和不耐倒春寒品种中tae-miR167d及其靶基因ARF12的表达特异性,为小麦耐倒春寒品种的筛选提供依据。因此,在设计试验时,参考已经发表的文献选出耐倒春寒强的品种济麦22、临麦4号、京411和豫麦47,和耐倒春寒弱的品种鲁原502、周麦22、西农889和濮麦053,共8个品种参加试验[23,24]。首先分析了倒春寒抗性不同品种济麦22和西农889中不同器官和不同时期taemiR167d和靶基因ARF12的表达差异。结果显示,在多数情况下,tae-miR167d和靶基因ARF12呈负调控关系,这一结果验证了前期的研究结果[15]。另外,在雌雄蕊发育时期,济麦22和西农889的叶鞘和茎中tae-miR167d和ARF12表达存在明显差异,这是否是导致它们耐倒春寒能力不同的原因还需要进一步验证。同时,该结果也表明在小麦耐倒春寒过程中,叶鞘和茎发挥着重要作用。
分析8个小麦品种叶片和幼穗冷胁迫前后tae-miR167d和ARF12表达差异,发现受到冷胁迫后,多数品种中tae-miR167d和ARF12的表达呈负调控关系,这一结果与前期的研究结果一致[15]。分别以济麦22的叶片和幼穗为对照,分析不同抗性品种中tae-miR167d和ARF12表达差异发现,在叶片中,冷胁迫后除临麦4号中taemiR167d的表达水平高于ARF12,其余6个品种中tae-miR167d的表达水平均低于ARF12的表达水平,与济麦22中表达模式一致;在幼穗中,冷胁迫后济麦22、京411、豫麦47和西农889中taemiR167d的表达水平低于ARF12的表达水平,其余4个品种中tae-miR167d的表达水平高于ARF12的表达水平。这与最初耐倒春寒强的品种和耐倒春寒弱的品种的分类并不完全一致。分析可能的原因:一是上述小麦品种来自不同的生态区,对低温的耐受性不同;二是小麦耐倒春寒是一个复杂的生物学过程,单纯的从某个角度不能全面揭示耐倒春寒的机理[15,25-27]。
猜你喜欢
作物杂志(2023年3期)2023-08-07 01:04:08
华北农学报(2022年4期)2022-09-14 04:22:22
东北农业大学学报(2020年12期)2021-01-15 07:44:52
作文周刊·七年级版(2020年4期)2020-05-06 09:25:18
作文周刊·七年级读写版(2020年4期)2020-05-06 09:00:33
农业知识(2019年17期)2019-09-18 01:38:34
江苏农业科学(2019年10期)2019-07-08 03:30:59
江苏农业科学(2016年11期)2017-03-21 14:50:30
植物保护(2016年1期)2016-09-14 06:14:52
湖南农业(2014年9期)2014-04-02 07:03:37
