
我国七大流域大鳞副泥鳅形态学比较分析
2021-06-08 03:41陈鸿禄易少奎李军涛王卫民
华中农业大学学报(自然科学版) 2021年3期
陈鸿禄,易少奎,李军涛,王卫民
1.华中农业大学水产学院/农业动物遗传育种与繁育教育部重点实验室/农业农村部淡水生物繁育重点实验室,武汉430070;2.湖州师范学院生命科学学院,湖州 313000; 3.中国热带农业科学院热带生物技术研究所,海口 571101
形态学研究是分类学的最原始和最基础的方法,从形态学的水平对遗传变异与适应性进化进行检测和研究也是最直接和最快速的[1]。水产动物环境适应性最直接的外在表现是形态表型差异,不同地理分布的群体,会随着环境出现适应性进化[2]。
大鳞副泥鳅(Paramisgurnusdabryanus),隶属于鲤形目(Cypriniformes)、鳅科(Cobitinae)、花鳅亚科(Cobitinae)、副泥鳅属(Paramisgurnus),除青藏高原地区外,在我国的各大水域均有自然分布[3],因而是研究种间遗传变异与适应性进化的理想材料[4]。之前也有关于大鳞副泥鳅在各流域分布的报道[5],但只是调查了大鳞副泥鳅在某一个或两个流域的资源分布情况,对我国整个水域(七大流域:长江流域、海河流域、淮河流域、珠江流域、海河流域、辽河流域、松花江流域)大鳞副泥鳅缺乏系统的分析。近年来,研究者开始调查不同地理群体大鳞副泥鳅的种内群体形态差异情况[6-7]。也有学者根据线粒体DNA的Cytb基因序列、D-Loop序列、线粒体全基因组以及微卫星[8-11]对其群体进行遗传变异分析。尽管目前分子标记提供了较为丰富的群体遗传学数据,但是群体间形态学差异仍没有得到很好的结果,同时国内对于大鳞副泥鳅的研究只局限于流域水平,因此,对全国各流域的大鳞副泥鳅种内形态变异特点及划分规律具有重大意义。本研究比较大鳞副泥鳅不同地理群体间的形态差异,旨在为大鳞副泥鳅种群的遗传学研究提供一些基础材料,同时也为大鳞副泥鳅种质资源的保护及开发利用补充部分材料。
1 材料与方法
1.1 材 料
本研究所用样品于2013-2016年的5月至9月份采集,为确保所采集的样品为大鳞副泥鳅自然群体,一是采集大鳞副泥鳅样品所选取的地点避开大鳞副泥鳅的人工养殖区。二是大鳞副泥鳅样本由研究人员使用地笼捕获,或直接雇用当地农民捕捞。本研究在我国七大水系的各个水体及附属水体共选取了19个采样点(表1),采样进行鱼类形态学的观察、测量并记录。

表1 大鳞副泥鳅形态采样信息表Table 1 Sample details for P. dabryanus in China
1.2 方 法
形态学的测量参考周小云[12]方法。选择健康有活力的个体,观察体型、体色,舍弃腹部膨大的个体,然后使用氨甲酸乙酯(100 mg/L)麻醉,用电子游标卡尺(精确度为0.01 cm)测量外部可量性状,包括吻长(LS)、体长(LB)、尾柄高(HCP)、头长(LH)、体高(HB)、全长(LT)、前躯长(DPR)、眼间距(WL)、尾柄长(LCP)、体宽(WB)和腹鳍臀鳍间距(DAC)。本研究以秦岭淮河为分界线,将我国七大流域分为南北水系,其中南方水系包括长江流域、珠江流域和淮河流域,北方水系包括黄河流域、海河流域、辽河流域和松花江流域。
1.3 数据分析

2 结果与分析
2.1 前驱长/体长、尾柄长/体长与纬度的线性回归分析
大鳞副泥鳅前驱长/体长与纬度呈强负相关(r=-0.782),尾柄长/体长与纬度呈极强正相关(r=0.834)。其结果显示,大鳞副泥鳅前驱长/体长随着纬度上升而下降,而尾柄长/体长则随着纬度的上升而上升(表2)。
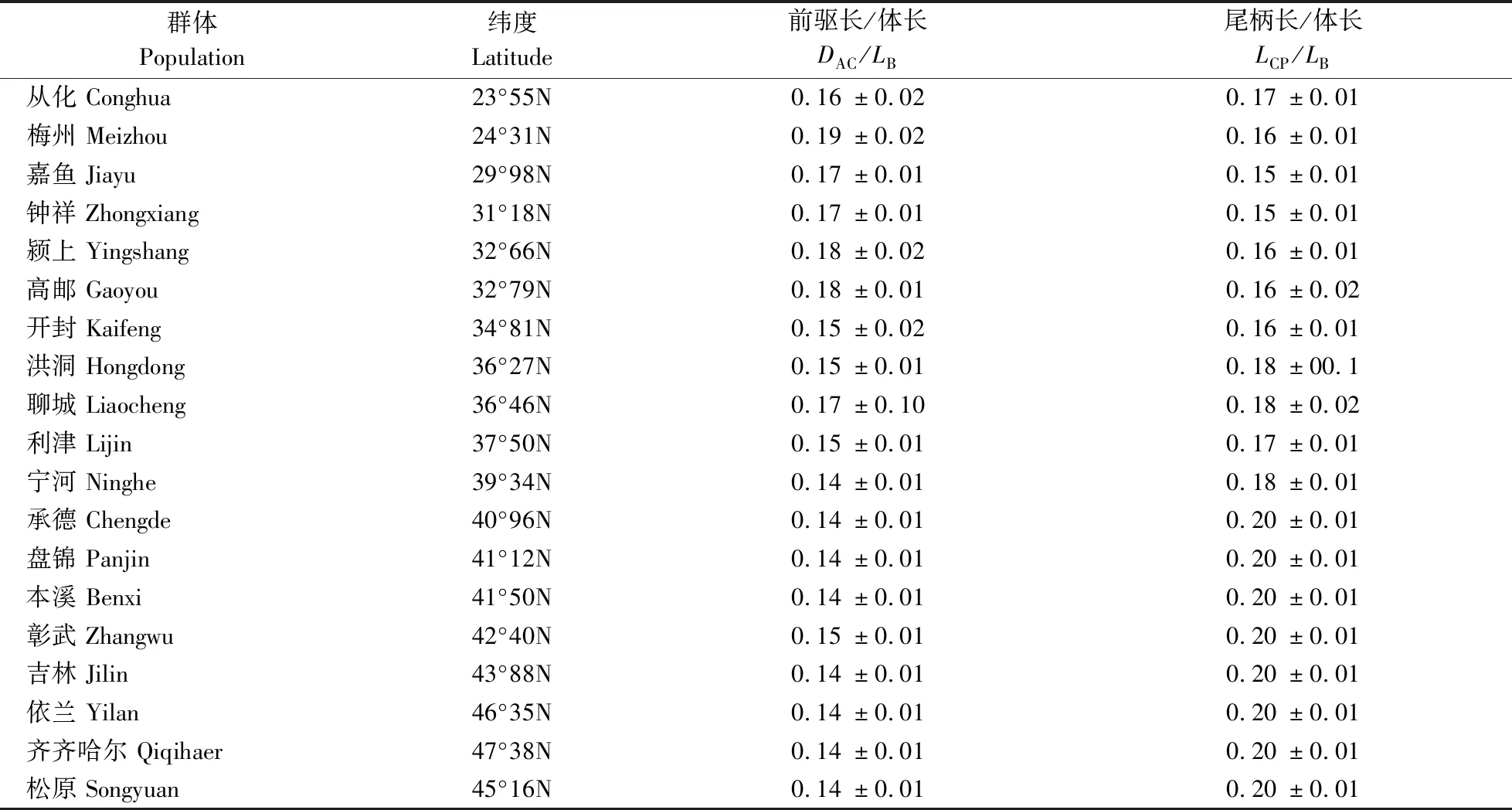
表2 大鳞副泥鳅群体前驱长/体长、尾柄长/体长与纬度关系表Table 2 Latitude and DAC/LB,LCP/LB of P. dabryanus in China
2.2 单因素方差分析
对我国七大流域的19个大鳞副泥鳅采集点样本的10项形态特征进行单因素方差分析。辽河流域和松花江流域的大鳞副泥鳅除体宽有差异之外,其他形态特征均不显著;长江流域、珠江流域、淮河流域、黄河流域和海河流域相互之间的体宽以及眼间距均存在显著差异;若将流域以南北水系进行划分,南方大鳞副泥鳅和北方大鳞副泥鳅群体在尾柄长、尾柄高、前驱长、体高、体宽、眼间距以及腹鳍臀鳍间距均有显著性差异。对不同流域大鳞副泥鳅群体的形态特征进行差异分析,差异系数比较结果:不同流域大鳞副泥鳅群体的全长、体高、吻长、前驱长、体高、体宽、腹鳍臀鳍间距的最大差异系数均小于1.28;尾柄长、尾柄高和眼间距的差异系数大于1.28,主要表现在南方水系和北方水系之间(表3)。

表3 大鳞副泥鳅群体单因素方差分析结果及最大变异系数Table 3 One-way ANOVA results among P. dabryanus population and maximum coefficient of variance
2.3 判别分析
对10项比例参数进行判别分析,得到7个流域大鳞副泥鳅的费歇尔判别函数。
代入判别函数进行判别,预测分析结果见表4,其中海河流域和辽河流域的大鳞副泥鳅判别准确率低于50%;将流域以南北方水系进行划分,松花江流域和辽河流域与南方水系各水域大鳞副泥鳅群体的判别准确率为100%,表明松花江流域和辽河流域与南方水域大鳞副泥鳅群体差异显著;同时,南方水系和北方水系的判别率均大于90%;通过判别函数对全国七大流域大鳞副泥鳅的综合判别率为65.9%。

表4 大鳞副泥鳅群体判别分析结果Table 4 Discriminant results of P. dabryanus populations
长江流域:Y=-2015.478+2453.538LT/LB+254.227LH/LB+2488.774LCP/LB+345.699HCP/
LCP+694.901DPR/LB+261.898WL/LH+1640.100WB/LB+202.881LS/LH-1483.621HB/LB
珠江流域:Y=-2015.206+2479.015LT/LB+209.659LH/LB+2499.845LCP/LB+323.578HCP/
LCP+713.319DPR/LB+122.028WL/LH+1521.919WB/LB+200.010LS/LH-1296.674HB/LB
淮河流域:Y=-2006.154+2472.211LT/LB+182.305LH/LB+2496.173LCP/LB+333.758HCP/
LCP+714.931DPR/LB+100.048WL/LH+1505.311WB/LB+178.730LS/LH-1243.829HB/LB
黄河流域:Y=-1975.034+2446.421LT/LB+243.013LH/LB+2475.907LCP/LB+316.938HCP/
LCP+687.470DPR/LB+219.091WL/LH+ 1499.637WB/LB+210.381LS/LH-1346.577HB/LB
海河流域:Y=-1972.251+2442.333LT/LB+249.978LH/LB+249.978LCP/LB+313.963HCP/
LCP+686.139DPR/LB+202.578WL/LH+1453.029WB/LB+200.841LS/LH-1321.218HB/LB
辽河流域:Y=-1941.886+2415.293LT/LB+235.749LH/LB+2590.695LCP/LB+313.432HCP/
LCP+681.997DPR/LB+210.370WL/LH+1455.352WB/LB+202.035LS/LH-1374.705HB/LB
松花江流域:Y=-1936.624+2417.562LT/LB+228.267LH/LB+2593.277LCP/LB+313.309
HCP/LCP+678.614DPR/LB+190.050WL/LH+1390.216WB/LB+203.691LS/LH-1322.986HB/LB
2.4 主成分分析
对大鳞副泥鳅10组形态性状比例参数进行主成分分析,结果显示,前3个主成分的累计贡献率达到62.9%,其第一主成分贡献率为38.1%,其他2个主成分的贡献率分别为13.8%、11.0%。第一主成分主要反映在尾柄高、尾柄长、体高和体宽等指标;第二主成分主要反映在体高和吻长等指标;第三主成分主要反映在全长、吻长和眼间距等指标。根据3个主成分绘制散点图(图1),我国七大流域明显分为两大支,一支为南方水系,另一支为北方水系,两大支间存在部分个体交叉现象,其中辽河流域和松花江流域形成一个相对集中的组,和南方水系没有交叉现象;海河流域和黄河流域与南方水系存在个体交叉混合。

图1 大鳞副泥鳅主成分散点图Fig.1 Scatter plots of P. dabryanus populations
2.5 聚类分析
对七大流域19个大鳞副泥鳅采集点所有样本校正值的平均值进行聚类,聚类结果和主成分分析的结果相似。其结果显示七大流域大鳞副泥鳅大致聚为两大类,南方水系大鳞副泥鳅除从化群体之外聚为一类,而北方水系聚为一类;对于北方水系大鳞副泥鳅,大致聚为两大类,辽河流域、松花江流域和海河流域的大鳞副泥鳅聚为一支,黄河流域的聊城群体和开封群体为一支;南方水系大鳞副泥鳅中,位于珠江流域的梅州群体与淮河流域大鳞副泥鳅先聚为一类,再与长江流域大鳞副泥鳅聚为一类。
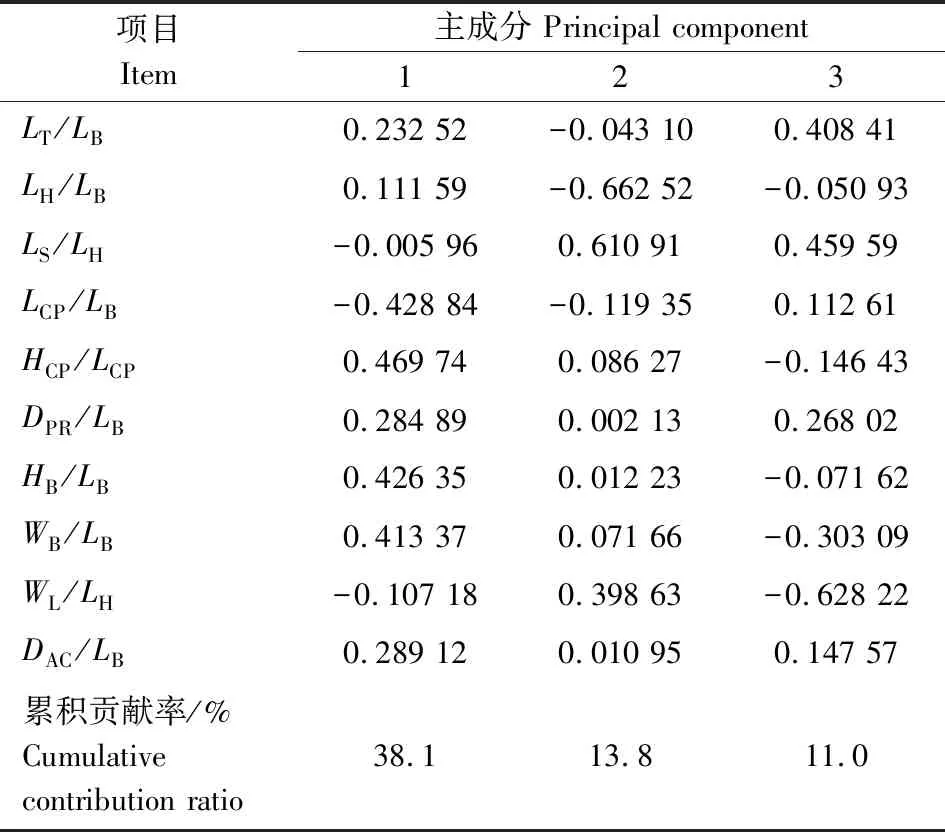
表5 大鳞副泥鳅群体主成分分析结果Table 5 Results of principal component analysis ofP. dabryanus populations

图2 大鳞副泥鳅群体聚类关系树Fig.2 Dendrogram showing the relationship of P. dabryanus populations
3 讨 论
形态学比较分析是研究物种遗传结构的有效方法之一,它能够提供与物种的生态和功能角色有关的信息[14]。本研究利用多种统计方法对我国七大流域大鳞副泥鳅进行形态学研究。单因素方差分析、判别分析、主成分分析和聚类分析结果均表明松花江流域和辽河流域的大鳞副泥鳅形态差异较小,且松辽流域和其他流域之间的大鳞副泥鳅形态差异较大。松辽流域位于我国的东北部,由南向北跨越了暖温带-中温带-寒温带3个不同的自然温度地带,据统计,该流域1980-2014年的气温平均值为5.11 ℃[15]。温度作为鱼类生长发育的重要影响因子,会直接影响鱼类整个形态特征[16]。Yi等[17]对我国泥鳅属3种鱼类进行转录组序列分析,发现生活在我国北方地区的北方泥鳅和黑龙江泥鳅其机体抵抗冬季极限温度有关的许多跨膜蛋白基因呈现快速进化与正向选择。据此推测,松辽流域大鳞副泥鳅在低温环境下,可能出现了环境适应性变化。在本研究中,松辽流域与其他流域的大鳞副泥鳅在全长、尾柄长以及前驱长表现出显著性差异,相较其他流域,松辽流域大鳞副泥鳅其体型尾部较长,头部较短。李雅娟等[18]通过对泥鳅属鱼类的形态差异比较分析,其研究结果表明泥鳅属鱼类的形态差异表现最为明显的部位是头部和尾部。本研究发现,大鳞副泥鳅不同地理群体间的形态差异主要集中在头部和尾部,和其结论是一致的,因此推测头部和尾部是花鳅亚科物种进化过程中变化较为迅速的表型。
鱼类在特定的栖息环境条件,最直接的适应性进化就是形态变异[19]。从线性回归分析结果可以得知,大鳞副泥鳅的体型随着纬度的上升而更细长,这可能与群体对环境的长期适应性、自身有限的迁移能力和地理隔离等因素有关。从单因素方差分析结果得知,我国南北水系的大鳞副泥鳅形态上存在较大差异,从体型上看,南方水系的大鳞副泥鳅较为粗短,而北方水系的大鳞副泥鳅较为细长。据此推测,大鳞副泥鳅可能由于南北地理隔离,出现栖息环境适应性进化。从判别分析结果得知,海河流域和辽河流域的大鳞副泥鳅判别准确率偏低。辽河流域大鳞副泥鳅个体错判的主要集中在松花江流域,其结果表明松花江流域和辽河流域的大鳞副泥鳅形态差异不明显;海河流域大鳞副泥鳅的判别准确率仅为24.9%,推测其可能原因有3个:一是海河流域与松辽流域及黄河流域地理位置较近,大鳞副泥鳅存在着丰富的基因交流。二是南水北调工程以及京杭大运河,为大鳞副泥鳅从南方水系向海河流域地迁移提供了渠道[20]。三是洪水促进了黄河流域和海河流域之间大鳞副泥鳅的基因流动[21]。松辽流域和南方水系大鳞副泥鳅之间的判别准确率为100%,其可能原因一是松辽流域位于我国东北部,和南方距离较远,流域之间存在一定程度的地理隔离;二是大鳞副泥鳅迁徙能力较弱。对七大流域大鳞副泥鳅进行判别分析,综合判别率为65.9%。其综合判别准确率较低,推测其有以下两个原因:一是本研究中的形态判别为种内判别;二是大鳞副泥鳅作为我国重要的淡水养殖鱼类,水产养殖运输频繁,使其在不同流域扩散。从主成分分析结果可以得知,七大流域大鳞副泥鳅的差异主要集中在尾部,体型肥瘦程度以及头部等方面。聚类分析结果中,七大流域的大鳞副泥鳅大致分为两大支,南方水系除从化群体之外为一支,北方水系为一支。聚类分析结果和主成分分析结果大体上是一致的,表明不同地理群体的大鳞副泥鳅在形态上既相似,同时又有一定程度的差异[22]。此研究结果和之前学者对我国甘肃省裂腹鱼的地理种群分化研究结果类似[23]。不同地理条件下的鱼类会产生与其栖息环境相适应的形态变异[24],如嘉陵江中游饵料资源丰富,中游的蛇鮈群体通过增加头长等方式来提高摄食频率[25]。推测其造成大鳞副泥鳅南北分化的原因可能有两个:一是南北方在气候、气温、降水量等自然因素存在差异[26];二是大鳞副泥鳅迁徙能力较弱。
综上所述,基于外部形态性状对我国七大流域大鳞副泥鳅进行区分具有一定的局限性,在今后的研究中,仍需结合更敏感的基因标记(如线粒体基因标记、核基因标记)对我国七大流域的大鳞副泥鳅的种群结构做进一步研究。
猜你喜欢
海河水利(2021年6期)2021-12-28
江苏农业科学(2021年19期)2021-11-18
建材发展导向(2021年23期)2021-03-08
经济研究导刊(2019年19期)2019-08-24
南水北调与水利科技(2018年6期)2018-02-01
环境保护与循环经济(2017年8期)2017-03-22
环境保护与循环经济(2017年11期)2017-03-16
安徽农业科学(2016年4期)2016-10-21
水科学与工程技术(2016年3期)2016-07-10
支点(2016年3期)2016-03-21
