
繁殖期黄喉雉鹑在景观尺度上的生境选择
2021-06-03 09:03:10冉江洪
生态学报 2021年8期
徐 雨,张 凯,窦 亮,杨 楠,王 彬,冉江洪,*
1 贵州师范大学生命科学学院,贵阳 550025 2 四川大学生命科学学院生物资源与生态环境教育部重点实验室,成都 610065 3 西南民族大学青藏高原研究院,成都 610041 4 西华师范大学生态研究院,南充 637002
生境是动物赖以生存和繁殖的场所,对种群的存活与延续具有重要的意义[1]。在长期进化过程中,动物对生境的选择表现出特定的环境适应性,同时也具有一定的尺度依耐性[2]。根据Johnson[3],动物的生境选择遵循4个基本层次:第1级是对地理区域的选择(区域尺度),第2级是在特定区域内对家域的选择(景观尺度),第3级是在家域内对不同生境斑块的选择(家域尺度),第4级是在特定生境斑块内对觅食、筑巢、夜栖等位点的选择(斑块尺度)。研究物种的生境选择,对探讨生物对环境的适应,以及开展物种保护和管理工作,具有重要的价值[4- 6]。
黄喉雉鹑(Tetraophasisszechenyii)隶属鸡形目雉科,是我国特有鸟种,国家I级重点保护动物[7-8]。分布于四川西部、青海南部、云南西北和西藏南部,栖息在海拔3350—4600 m之间的针叶林、高山灌丛、草甸及林线以上的岩石苔原地带[8-9]。对于黄喉雉鹑的生境选择和利用,早期刘少初和次仁、吴毅等、格玛嘉措、Potapov、文陇英等进行过简单的描述性报道[10-14]。就定量研究而言,Xu等、Zhang 等、徐雨等、Wang 等 从觅食地、夜栖地、巢址和沙浴地等角度对植被偏好及其环境影响因素进行了一定分析[15-22]。但是,先前研究很少考虑到生境选择的层次性,尤其对植被类型选择的研究而言,如果缺乏对生境选择层次性的考虑,可能会使理解出现偏差[1,23-24]。
本研究依据Johnson[3]对生境选择层次性的定义,从景观尺度上确定黄喉雉鹑在繁殖期的占域情况,然后通过建立生境模型分析占域与植被、地形等环境因子之间关系,以此分析黄喉雉鹑在繁殖期的生境选择形式,探讨该物种对环境的适应性,为物种的生境保护和管理提供一定理论基础。
1 研究方法
1.1 研究地区
研究地位于四川省甘孜藏族自治州雅江县帕姆岭(30°06′N, 101°11′E),海拔3350—4300 m,面积50.9 km2。该区域气候属青藏高原亚湿润气候区,分布有6种典型植被类型(图1),即冷红杉林(FF)、针阔混交林(DBCF)、高山松林(PPF)、高山栎丛(OK)、杜鹃灌丛(RS)及草地(AM)。各植被的特征描述如下(均值表示为±标准误):
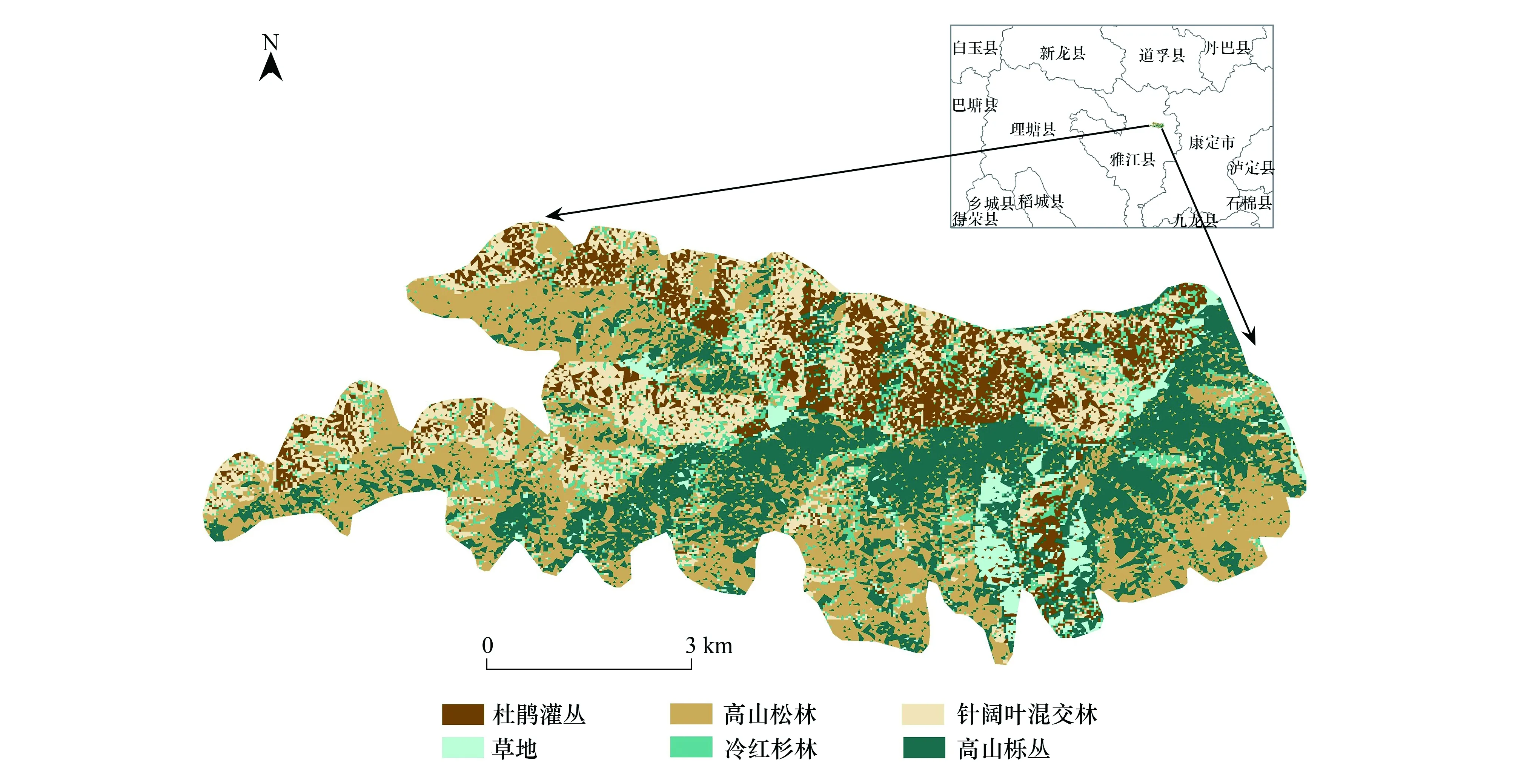
图1 图示研究区的位置及植被组成Fig.1 Map showing the location of the study site and vegetation structure
(1)冷红杉林(n=16):主要分布于海拔3350—4100 m,面积占7.7%。上层乔木盖度(44.0±7.4)%,优势树种为鳞皮冷杉(Abiessquamata)、大果红杉(Larixpotaniniivar.macrocarpa)。林下灌木盖度(33.0±5.1)%,草本盖度(12.5±3.8)%,落叶盖度(50.5±6.7)%。
(2)针阔混交林(n=13):分布于海拔3350—3600 m,面积17.2% 。乔木盖度(39.4±4.8)%,优势树种为大果红杉、桦木(Betulaspp.)、川西云杉(Picealikiangensisvar.balfouriana)等。林下灌木盖度(28.9±4.6)%,草本盖度(14.4±2.9)%,落叶盖度(48.5±4.5)%。
(3)高山松林(n=20):分布于海拔3350—3850 m,面积占29.6%。乔木层盖度(38.3±3.7)%,高山松(Pinusdensata)为绝对优势种,林下灌木盖度(32.1±4.4)%。由于凋落的松针难以分解,落叶层盖度高达(77.8±5.0)%,草本盖度很低, 仅为(5.4±2.2)%。
(4)高山栎丛(n=27):分布于海拔3700—4300 m,面积占27.3%。上层盖度(36.2±6.0)%,川滇高山栎(Quercusaquifolioides)是绝对优势种,其他灌木稀少,草本盖度(15.8±3.4)%,落叶盖度(49.4±3.9)%。
(5)杜鹃灌丛(n=8):分布于海拔3700—4200 m,面积占14.1%。盖度(55.1±4.9)%,包含多种杜鹃,比如光亮杜鹃(Rhododendronnitidulum)、淡黄杜鹃(Rh.flavoflorum)等,其他常见灌木有柳(Salixspp.)、金露梅(Dasiphorafruticosa)、小檗(Berberisspp.)等。草本盖度(39.1±4.4)%,落叶盖度(24.6±2.7)%。
(6)草地:零星分布于上述各种植被之间,面积很小,草本盖度>80%,以禾本科草本为主。
除黄喉雉鹑之外,研究区还分布有血雉(Ithaginiscruentus)、白马鸡(Crossoptiloncrossoptilon)、高原山鹑(Perdixhodgsoniae)等鸡形目鸟类。主要的鸟类天敌有胡兀鹫(Gypaetusbarbatus)、高山兀鹫(Gypshimalayensis)、大鵟(Buteohemilasius)。兽类天敌有豹猫(Prionailurusbengalensis)、藏狐(Vulpesferrilata)、黄鼬(Mustelasibirica)及香鼬(M.altaica)等。
1.2 野外调查
首先,从研究区中心沿各个主要方位设置调查样带(共设置调查样带10条,每条样带长度为0.3—5 km,总长度22.5 km)。然后,依据鸟类调查的相关准则[5]以及黄喉雉鹑的家域值大小[25],在每条样带上每隔250 m设置1个调查样点(共计设置了84个样点,但由于4个样点的缓冲半径超出研究区范围,实际纳入分析的样点为80个)。
使用录音回放技术开展调查(录音为一段40 s高质量领域鸣叫声;设备型号:京华DVR- 892)。调查通常在晴朗天气进行,调查时间为每日07:30—11:00和14:00—17:40。在调查时,研究人员(2人组成)沿样带以大约2 km/h速度行进。当到达某一样点,首先播放录音;然后,记录半径250 m范围内是否有黄喉雉鹑的应答声。若有应答,则立即停止回放;否则,2 min间隔后,重复回放1次。研究者在每个样点的停留时间为10 min。
调查工作于2009年4—6月开展,调查每月进行1次,共开展了3次调查。
1.3 数据处理和分析
首先,下载研究区的Landsat 7 TM影像图(2009年3月,分辨率 30 m × 30 m),使用ERDAS 8.6进行监督分类。随后,导入ArcGIS 3.2,使用野外调研中实际获得的182个植被经纬度点对分类图进行校正;以1∶100000地形图为底图,提取等高线及赋高程值(等高线间距是40m),生成DEM图(分辨率30 m),求出以样点为中心250 m缓冲半径内的平均高度(ALT)、平均坡度(SLP)、平均坡向(ASP)及各种植被的面积占比。然后,对所有变量进行Spearman 相关性分析。若发现两变量间的相关系数绝对值大于或者等于0.5,则认为变量之间存在一定相关性[26],在随后模型中仅纳入生物学意义较强或之前研究[16,18-19]认为重要的变量进入分析。
使用逻辑斯蒂回归模型预测影响黄喉雉鹑占域的重要因素。对某一监测样点,倘若在3次调查中至少探测到1次黄喉雉鹑的鸣声应答,则认为黄喉雉鹑有占域;否则,视为未占域。在分析中仅纳入单级因子(不考虑因子间二级以上的交互作用),建立不同组合的备选模型(包括零模型)。使用修正AIC(AICc)评判和选择模型。首先,计算出即各模型的AICc值与最小AICc的差,即ΔAICc。通过比较各模型的 ΔAICc值和 Akaike 权重值(wi),确定各模型的优劣程度。若最优模型的ΔAICc> 2,则使用该模型计算参数估计值[27];否则,对95%置信模型(wi由高到低排序后累加值 ≤ 0.95的模型)进行模型平均[27-28],然后比较每个变量的相对重要性(Σwi)和95%置信区间。如果某一变量相对重要性较大且95%置信区间不包含0值,则认为该变量对模型的贡献较大[29]。所有分析在R 3.5.1[30]上进行,模型拟合和模型平均借助glmulti[31]和MuMIn[32]两个插件完成。
2 结果
在纳入分析的80个监测样点中,250 m缓冲半径内发现有黄喉雉鹑出现的样点是41个,即,51.3%的监测样点是黄喉雉鹑的占域点。
Spearman 相关性分析显示,样点250 m缓冲半径内高山松林占比与海拔、杜鹃灌丛占比的相关性较高,高山栎丛占比与杜鹃灌丛占比、针阔混交林占比相关性较高(表1)。基于变量的相对重要性,在随后分析中,剔除海拔、杜鹃灌丛占比和针阔混交林占比3个变量。即,最终进入模型分析的变量是坡度、坡向、高山松林占比、草地占比、冷红杉林占比及高山栎丛占比等6个。

表1 生境变量的相关分析
共建立了64(26)个逻辑斯蒂回归模型。其中,有多个模型的ΔAICc≤ 2,即没有1个最优拟合模型(表2)。 因此,选择95%置信备选模型(wi由高到低排序后累加值 ≤ 0.95的模型)进行模型平均。模型平均的结果显示,3个变量,即高山松林占比、冷红杉林占比和高山栎丛占比的相对重要性(Σwi)较大,且参数估计值的95%置信区间不包含0值(表3)。因此,这3个变量是决定黄喉雉鹑生境选择的最重要因子。根据模型平均的预测,无论是阳坡或是阴坡,黄喉雉鹑都偏爱冷红杉林和高山栎丛占比较高的地方,而不喜好高山松林占比较高的地方(图2)。

表2 黄喉雉鹑生境选择预测的95%置信模型
表3 基于模型平均的生境变量参数估计和相对重要性值
Table 3 Model-averaged coefcients and relative importance calculated for variables predicting habitat selection of Buff-throated Partridges
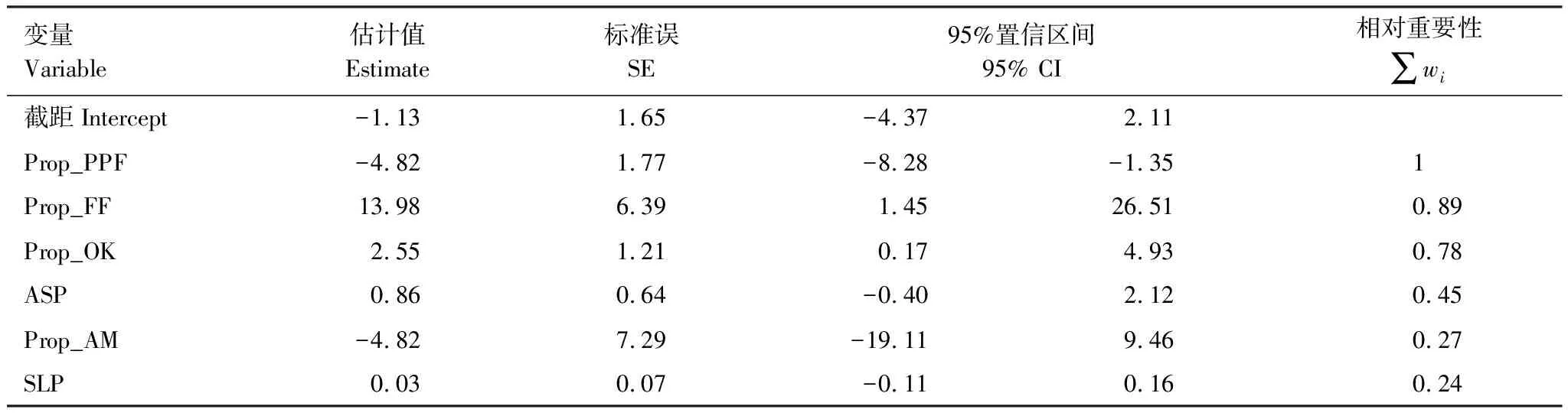
表3 基于模型平均的生境变量参数估计和相对重要性值
变量Variable估计值Estimate标准误SE95%置信区间95% CI相对重要性 ∑wi截距Intercept-1.131.65-4.372.11Prop_PPF-4.821.77-8.28-1.351Prop_FF13.986.391.4526.510.89Prop_OK2.551.210.174.930.78ASP0.860.64-0.402.120.45Prop_AM-4.827.29-19.119.460.27SLP0.030.07-0.110.160.24
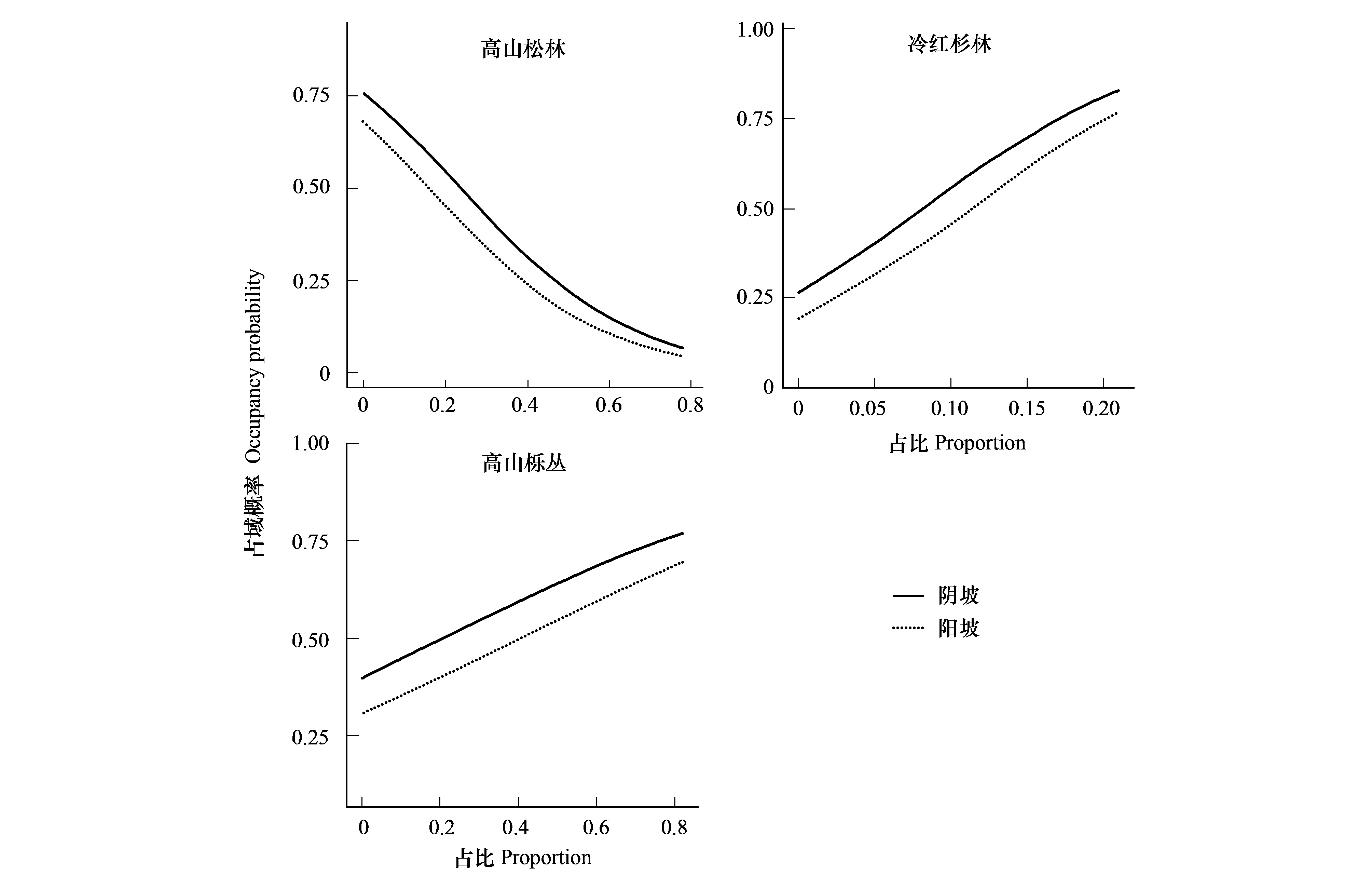
图2 黄喉雉鹑占域概率及高山松林、冷红杉林、高山栎丛占比的影响Fig.2 Occupancy probability of Buff-throated Partridges Proportion of pine forests, fir-larch forests and oak thickets
3 结论与讨论
先前研究报道黄喉雉鹑在繁殖期倾向于选择冷红杉林和高山栎丛,而拒绝使用高山松林[16,18-19]。本研究依据Johnson[3]对生境选择层次性的定义,从景观尺度上调查黄喉雉鹑在繁殖期的占域情况,进一步严格地证实黄喉雉鹑偏爱冷红杉林和高山栎丛占比高的地方,而不喜好高山松林占比高的地方。
鸟类的生境选择通常与逃避天敌和觅食相关[5,23,33]。研究区的冷红杉林和高山栎丛同时具有较大乔木郁闭度和林下灌木盖度,选择这两种植被可能有助于躲避天敌。但是,高山松林也具有较大乔木郁闭度和林下灌木盖度,而黄喉雉鹑却不喜好,推测可能是对捕食风险和食物资源权衡的结果。与同域分布的白马鸡等鸡形目鸟类相比较,黄喉雉鹑棕灰色的羽毛本身能降低捕食风险。在这种情况下,黄喉雉鹑在选择生境时可能会重点考虑食物资源的可获得性[22]。黄喉雉鹑以草食性为主(主要啄食草本的茎叶、果实、种子及根),且啄刨能力不甚强[8,10,22,34]。高山松林下落叶主要以高山松松针为主,因松针不易分解,致使林下落叶盖度很大,落叶厚度很深,而草本盖度很低,不利于黄喉雉鹑获取食物。比如,果实或种子被落叶层深深地埋藏,将难以被取食。进一步研究有待从植被结构和地被层特征开展定量分析,以此阐述其生境选择的机制。
尽管当前研究揭示冷红杉林和高山栎丛是黄喉雉鹑在繁殖期的重要生境。但是,在同一研究位点,先前研究报道黄喉雉鹑成体夜栖时完全使用冷红杉林,而不选择其他生境;幼体除了使用冷红杉林外,还使用杜鹃灌丛[15,17]。因此,推测黄喉雉鹑对生境的需求可能存在时间尺度(季节、日)的变化。
就保护管理而言,冷红杉林和高山栎丛作为黄喉雉鹑的重要生境,应该重点受到保护。高山栎是当地居民薪材砍伐的主要对象,尤其应受到管控。考虑到地处高寒地区以及特殊的社会经济状况,当前不可能完全杜绝当地人取材,因此有必要对伐取薪材的时间、地点和规模进行规范,杜绝滥砍滥伐[18]。同时,鉴于黄喉雉鹑的生境选择具有时间变异性,建议在保护管理和规划工作中维持多样性化或镶嵌式的植被结构。在下一步工作中,需对生境面临的主要威胁进行调查,以制定更具针对性的保护策略。
致谢:感谢四川省格西沟自然保护区对野外工作提供的帮助与支持,感谢帕姆岭寺对野外生活给予的照顾和帮助,感谢岳汉秋和廖立欢对植被解译和GIS作图提供的帮助。
猜你喜欢
Plasma Science and Technology(2022年9期)2022-08-29 00:43:22
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
科学导报(2019年73期)2019-12-20 08:46:53
水土保持研究(2019年6期)2019-10-19 03:33:32
语文世界(小学版)(2018年9期)2018-09-18 20:01:50
科学之谜(2018年3期)2018-04-09 06:37:46
连环画报(2016年10期)2016-12-16 05:13:34
心潮诗词评论(2016年1期)2016-12-01 05:48:31
乡村地理(2016年2期)2016-06-15 20:29:26
防护林科技(2015年5期)2015-06-10 11:42:52
