
藏北高寒草甸根系生物量与碳氮分布格局及关联特征
2021-06-03 04:28张豪睿
生态学报 2021年9期
张豪睿,付 刚
1 中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室 拉萨高原生态系统研究站, 北京 100101 2 中国科学院大学, 北京 100049
根系作为植物地下部分最敏感的器官,探究其在全球变化背景下的响应有重要理论及应用价值。一方面,根可以固定植物体,抵抗外界的自然灾害(大风、洪水等);另一方面,根系作为植物的营养器官吸收土壤中的营养物质,促进生态系统能量流动与物质循环[1-2]。土壤中的氮通过有机物的分解作用形成植物可利用的氮元素被根系吸收,而植物死亡后被微生物分解成为土壤中氮元素的主要来源[3],植物根系中碳氮元素的含量主要受外界环境的影响,而土壤作为植物生长的载体与植物根系碳氮元素的吸收密不可分。根系中的养分浓度不仅反映了植物本身的生物学特征,也反映了不同生长环境下植物根系对土壤中营养物质的吸收和利用情况[4]。由根系生物量与根系碳氮含量决定的根系碳氮库能反映出土壤养分库的大小。植物根系的生长发育与植物根系、土壤中的碳氮元素息息相关。由于草地生态系统的根冠比高于其他生态系统[5-6],植物根系是土壤中有机碳的主要来源,根系生物量可以用来衡量土壤的碳输入[7]。因此根系与土壤连接成为“根土系统”直接影响着整个植被系统[8],研究草地中根系生物量与碳氮元素的分布至关重要。
有关根系生物量随土壤深度的研究在不同草地系统已开展,内蒙古的草甸草原[9]、新疆天山高寒草地[10]、祁连山北坡天然草地[11]和西藏那曲高寒草甸[12]等。有关草地生态系统根系生物量沿土壤深度变化的研究发现,随深度的增加,根系生物量的分布格局呈现不同的形态,例如,呈现“T”型分布[13- 16],有些在随深度的变化过程中存在两个峰值[17]。尽管如此由于不同类型草地生态系统的环境差异较大,根系生物量总量及其在不同土层中的分布不同,还需要进一步的探究。
高寒草甸作为青藏高原独特的环境条件影响下的代表性草地系统,研究高寒草甸根系生物量的分布对草地保护与退化草地恢复有重要意义。虽然已经有对西藏地区高寒草甸的探究[16-18],但并没有研究海拔的影响,不同海拔下根系在土壤中的垂直分布是否相同,不同海拔下总根系生物量是否会有差异等并未进行探究。青藏高原独特的环境主要是由于其高海拔的影响造成,因此研究根系生物量与碳氮含量沿海拔的分布尤为重要。且相关研究在探究根系生物量在土层的分布仅结合土壤理化性质进行探讨,并未与植物根系碳氮相结合。
本研究以西藏自治区拉萨市当雄县草原站的高寒草甸为研究对象,对2011年群落根系生物量、根系碳氮和土壤碳氮含量等进行了测定,探讨了藏北高寒草甸根系生物量与根系碳氮、土壤碳氮的分布格局及其关联特征。
1 材料与方法
1.1 研究区域概况
研究区位于西藏自治区拉萨当雄草原站,气候类型为高原性季风气候,年均气温为1.87 ℃,年降水量为474.9 mm,降水主要集中在生长季(6—8月)。本研究样地沿海拔分布,三个海拔分别为:低海拔(4300 m,30°30′N, 91°04′E)、中海拔(4500 m,30°31′N,91°04′E)、高海拔(4700 m,30°32′N, 91°03′E),随海拔的升高土壤温度降低,降水与土壤湿度增加[19]。低海拔和中海拔为草原化高寒草甸,其优势物种为小嵩草(Kobresiapygmaean)、黒褐苔草(Carexatrofusca)和丝颖针茅(Stipacapillacea),而高海拔为典型的高寒嵩草草甸,其优势种为小嵩草(Kobresiapygmaean)[20]。
1.2 样品的采集和分析
2011年8 月,在三个海拔样地中,垂直地面分层(0—10 cm,10—20 cm,20—30 cm,30—40 cm和40—50 cm)采集土壤样品,每个海拔设置3个重复,样方大小为25 cm×25 cm。采集的土样(包含根系)封装于自封袋中。用2 mm孔径的筛子将自封袋中的新鲜土样过筛,挑出肉眼可见的根系,并用水冲洗。将洗后的根系在65℃的烘箱中烘干至恒重,称重得到根系生物量。然后粉碎样品,用碳氮元素分析仪(Elementar Variomax CN)测定根系碳氮含量。
将筛后的部分鲜土用于测定水溶性有机碳(dissolved organic carbon, DOC)、微生物量碳(Microbial biomass carbon, MBC)、溶解性总氮(dissolved total nitrogen, DTN)、微生物量氮(Microbial biomass nitrogen, MBN)、pH和电导率。将筛后的部分土壤鲜样风干后用于测量土壤有机碳(soil organic carbon, SOC)和全氮(total nitrogen, TN)。pH值采用酸度计测量;电导率用标准DDS- 11AT型电导率仪测定;DOC、MBC、DTN、MBN、SOC和TN的测量方法与Fu等[21]的测量方法一致。
1.3 数据分析
统计方法使用了两因素方差分析、多重比较(LSD)、非线性拟合和相关分析(pearson相关)。通过构建结构方程模型(Structural Equation Model,SEM)分析了根系生物量与根系碳氮以及土壤理化性质的关系。所有的统计分析均由R 3.5.2 完成,画图由Sigmaplot 12.5与R 3.5.2 完成。统计分析涉及的包有stats、agricolae、psych、FactoMineR、factoextr、dplyr和lavaan等,画图由ggplot2包完成,结构方程模型图由Visio 2019完成。
2 结果与分析
通过两因素方差分析发现,除了微生物量碳氮比外,海拔对所有指标均有显著性影响;除了根系氮含量和微生物量碳氮比外,土壤深度对所有指标均有显著性影响;除了根系碳氮含量、土壤碳氮比、微生物量碳氮比、pH和电导率外,两因子交互作用对其他指标均有显著性影响(表1)。
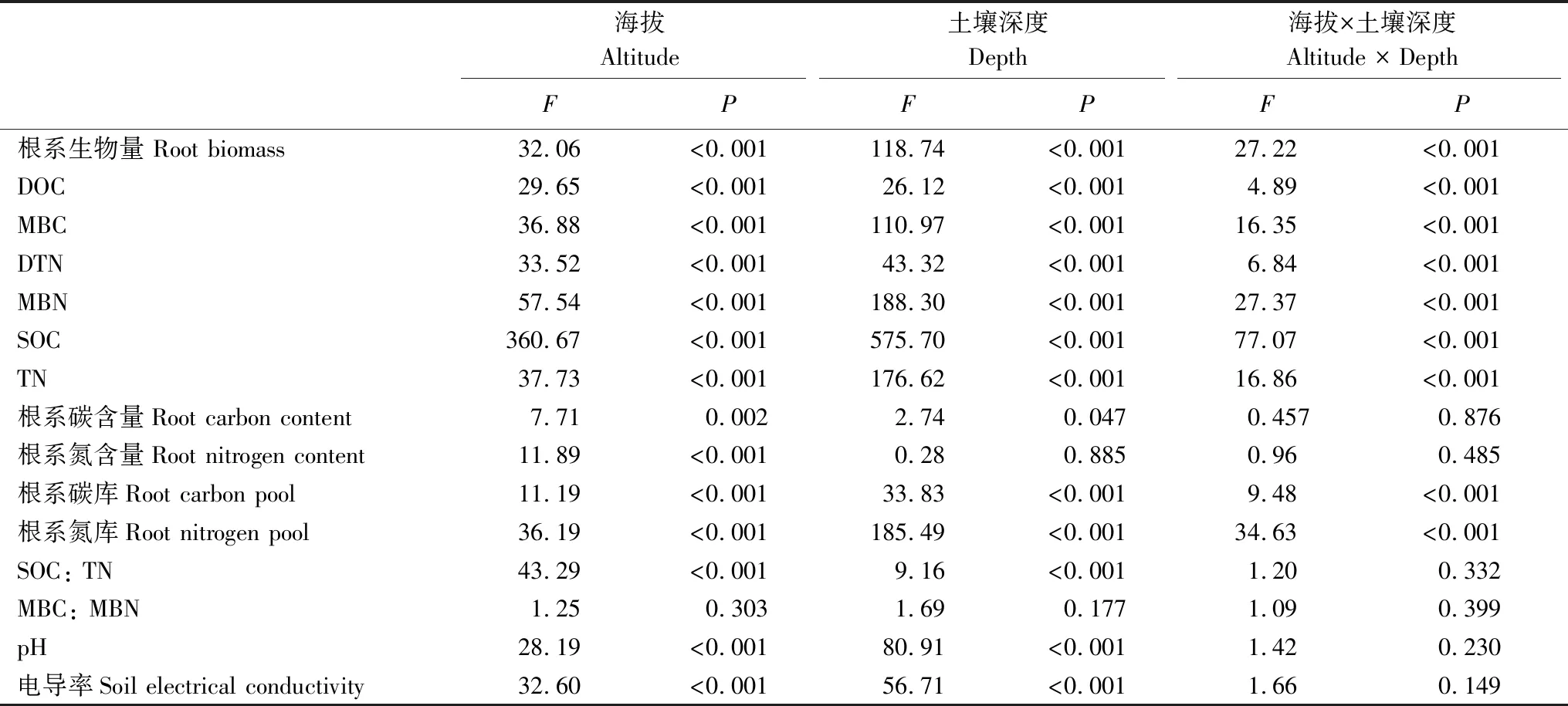
表1 海拔和土壤深度对根系生物量、根系碳氮以及土壤理化性质的影响
2.1 海拔与土壤深度对碳氮、pH和电导率的影响
2.1.1海拔对碳氮、pH和电导率的影响
0—50 cm深度,4300 m的DOC最大;4700 m的MBC、SOC、DTN、MBN和TN最大,且SOC随海拔的升高逐渐升高(图1)。0—20 cm深度,4700 m的DOC、MBC、SOC、DTN、MBN和TN含量均显著大于4300 m与4500 m(图1)。4300 m的MBC/MBN在每个土壤深度均大于4500 m,SOC/TN在每个土壤深度均随海拔逐渐升高(图1)。4700 m在每个土壤深度的pH均最低,4300 m的电导率均最高(图2)。在每个土壤深度,4300 m的根系氮含量均显著高于4700 m;4500 m的根系碳含量均为三个海拔最低,4700 m根系碳氮比均为三个海拔最高(图3)。

图1 不同海拔下土壤碳氮组分随土壤深度的变化规律Fig.1 Variation of soil carbon and nitrogen components with soil depth at different altitudesDOC:水溶性有机碳 Dissolved organic carbon;MBC:微生物量碳 Microbial biomass carbon;DTN:溶解性总氮 Dissolved total nitrogen;MBN:微生物量氮 Microbial biomass nitrogen;SOC:土壤有机碳 Soil organic carbon;TN:全氮 Total nitrogen
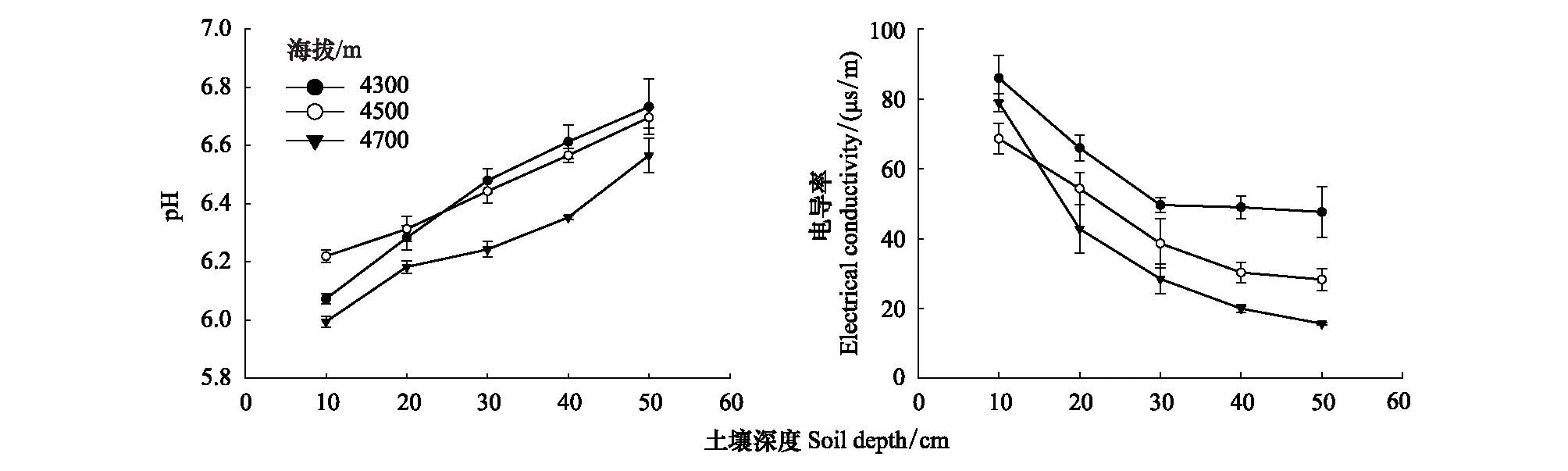
图2 不同海拔下土壤pH和电导率随土壤深度的变化规律Fig.2 Variation of soil pH and conductivity with soil depth at different altitudes
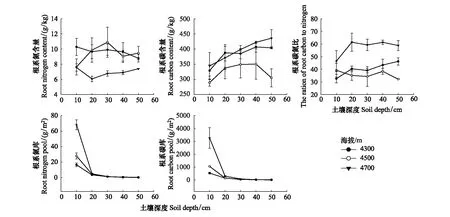
图3 不同海拔下根系碳氮组分随土壤深度的变化规律Fig.3 Variation of carbon and nitrogen components in roots with soil depth at different altitudes
2.1.2土壤深度对碳氮、pH和电导率的影响
三个海拔的DOC、MBC、SOC、DTN、MBN、TN和SOC/TN都随土壤深度的增加呈现逐渐减少的趋势,且4700 m土壤碳氮含量随土壤深度的变化最为显著(图1)。4300 m与4500 m的MBC/MBN随土壤深度的变化趋势相似,随土壤深度的增加,0—20 cm的MBC/MBN逐渐升高,20—40 cm逐渐降低,40—50 cm逐渐升高;4700 m 的MBC/MBN随土壤深度的增加呈现上升趋势(图1)。三个海拔的pH随土壤深度的增加逐渐升高,而电导率随土壤深度的增加逐渐降低(图2)。
三个海拔的根系碳、氮含量随土壤深度的变化趋势不一致,4300 m根系氮含量呈逐渐下降的趋势,4500 m根系氮含量随土壤深度先增高后降低,4700 m根系氮含量随土壤深度先降低后升高(图3)。4300 m与4700 m根系碳含量随土壤深度的增加逐渐增高,而4500 m根系碳含量随土壤深度的增加先增高后降低(图3)。三个海拔的根系碳、氮库均随土壤深度的增加逐渐降低(图3)。
2.2 海拔与土壤深度对根系生物量的影响
0—50 cm深度的根系生物量随着海拔的升高逐渐增加。三海拔间根系生物量多重比较(LSD)结果表明:0—30 cm深度,根系生物量4700 m显著大于4300 m与4500 m;30—40 cm深度,三个海拔间无显著性差异;40—50 cm深度,4300 m显著大于4500 m与4700 m。随土壤深度的增加,三个海拔根系生物量逐渐减少(图4)。表2为三个海拔下,不同土壤深度根系生物量占0—50 cm总根系生物量的比例。由此可以看出,根系生物量主要分布在上层土壤中(0—10 cm),三个海拔在0—10 cm 深度所占的比例分别为72.94 %、85.61 %和89.44 %。对不同海拔间根系生物量百分比进行多重比较(LSD)发现:0—10 cm深度,三海拔根系生物量百分比无显著性差异;10—20 cm深度,4300 m根系生物量百分比显著大于4700 m,4500 m与4300 m、4700 m无显著性差异;20—50 cm深度,4300 m根系生物量百分比显著大于4500 m与4700 m,4500 m与4700 m无显著性差异。由此可以看出,随海拔的增加,根系生物量越集中分布于上层土壤,下层土壤根系生物量分布越少。

表2 不同土壤深度土壤中根系生物量的分异
图4为三个海拔的根系生物量及其平均值随土壤深度的变化特征,对三个海拔根系生物量及三海拔均值与土壤深度进行回归分析,所有方程均显著,且R2趋近于1。随着土壤深度的增加,根系生物量呈指数下降。
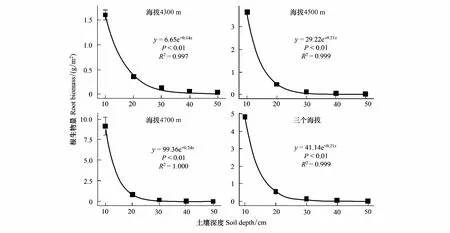
图4 根系生物量与土壤深度的指数回归Fig.4 Exponential regression of root biomass and soil depth
2.3 根系生物量与根系碳氮以及土壤理化性质的关系
Pearson相关分析(图5)发现:根系生物量与DOC、MBC、DTN、MBN、SOC、TN和电导率呈现出显著的正相关关系,与pH呈现显著的负相关关系。结构方程模型的χ2值不显著(χ2=21.66,P=0.753)(图6),表明此模型能较好的反映根系生物量与解释因子的关系。该模型解释了99.5 %的根系生物量的变化。根系氮库直接影响根系生物量且与根系生物量呈显著正相关,根系碳库通过影响根系氮库间接影响根系生物量。微生物量碳通过影响根系碳、氮库间接影响根系生物量。pH通过影响电导率间接影响根系生物量。

图5 根系生物量与根系碳氮及土壤因子的相关分析Fig.5 Correlation analysis between root biomass, root carbon, nitrogen and soil factors

图6 根系生物量与根系碳氮及土壤因子的结构方程模型Fig.6 Structural equation model of root biomass and root carbon and nitrogen and soil factors* P<0.05, ** P<0.01, *** P<0.001;实线表示P值显著,虚线表示P值不显著
3 讨论
3.1 海拔与土壤深度对根系碳氮与土壤碳氮的影响
与深层土壤相比,0—20 cm深度,4700 m与较低海拔处土壤碳氮含量差异最为显著(图1),这可能是由于微生物多存在于上层土壤,使得上层土壤中的碳氮含量分布较多;4700 m海拔高、温度低、生物活性受到抑制,从而更多的碳氮元素得以储存在土壤中。与之前的研究相似[21-22],本研究发现土壤碳氮比与海拔呈现正相关(图1),这可能是由于随海拔的升高,温度降低,微生物活性下降,氧化分解产生的有效氮元素较少[23]。微生物量碳氮比多集中于5—10,由于真菌碳氮比大于10,细菌碳氮比小于5[24],从而可以推断出4300 m真菌与细菌的比值大于4500 m,这与之前研究的实测数据一致[20],由此可以看出微生物量碳氮比可以反映出土壤中真菌与细菌的比例。三个海拔的根系碳含量、氮含量、碳氮比中,根系氮含量在不同海拔下差异最显著,这符合“温度-植物生理假说”[25]。三海拔中,4700 m处温度最低[26-27],在这种营养物质的分配方式下,4700 m根系中的氮含量最低。
土壤中碳氮含量均随土壤深度的增加逐渐减少,这与之前的研究结果一致[28- 30]。土壤有机质是土壤中碳元素与氮元素的主要来源[31],有机质含量主要来源于腐殖质中微生物对凋落物的氧化分解,并通过淋溶作用从土壤表层逐渐向下层输入,且有机质的含量取决于微生物氧化分解的程度与淋溶作用的强度[32]。随土壤深度的增加,MBC减少(图1),MBC一定程度上可以反映微生物量[33],微生物量减少导致氧化分解的速率降低;随土壤深度的增加,土壤中凋落物含量减少,微生物用来氧化分解的底物减少;随土壤深度的增加淋溶减弱[34],土壤的通气性也会随之降低,从而使得土壤中碳氮含量随土壤深度的增加逐渐减少(图1)。
3.2 海拔与土壤深度对根系生物量的影响
与之前的研究结果一致[35-36],本研究发现随海拔的升高根系生物量增加。根系生物量受到植物群落环境的影响,降水是影响青藏高原根系生物量动态的重要因素[18]。随海拔的升高,土壤湿度与空气湿度增加。与低海拔相比,高海拔较为充足的水分条件更利于根系生物量的积累。且随海拔的升高,土壤中营养元素较多(图1),从而为植物根系的生长提供充足的养分。
与之前的研究结果一致[12-13,16,37], 本研究发现随着土壤深度的增加,根系生物量呈指数下降,且随海拔的增加根系生物量越集中分布于上层土壤,这与土壤营养元素沿土壤深度的分布有关(图1)。土壤质地的差异也会影响根系生物量在土壤中的分布[38],随着土壤深度的增加,土壤颗粒变大,不利于根系的呼吸及对营养物质的吸收[16]。此外,根系生物量随土壤深度的变化趋势还可能与土层的含水量有关[39]。
3.3 根系生物量与根系碳氮、土壤碳氮等环境因子的关系
根系氮库能最好的解释根系生物量的变化,这可能与氮元素活跃的地方根系生物量大有关[40]。根系氮库较大表明植物根系对营养物质的吸收较好,有利于根的生长发育。根系生物量与土壤理化性质息息相关[41]。本研究中所有土壤pH值均呈酸性,这说明偏酸性的土壤更利于本研究区域植物的生长。电导率可以表征土壤的含盐量[42]。土壤深度越深,含盐量越低。土壤中盐分的积累是由于盐分随水分运输的结果,而植物根系对水分的吸收会影响土壤中盐分的分布,根生长旺盛的地方对水分的吸收更强,电导率更大。同时,土壤中盐分的分布会影响根系的生长[43],铵盐和硝酸盐等是促进根系生长的重要营养元素。青藏高原海拔高,垂直分异明显,不同海拔下土壤温湿度有显著差异[26,44]。因此,土壤的水热条件也是影响根系生物量生长的重要条件[18]。
4 结论
通过对海拔4300 m、4500 m和4700 m 的0—50 cm土壤深度的土壤碳氮及根系生物量的研究发现:随土壤深度的增加,根系生物量呈指数下降;随海拔的增加,根系生物量越集中分布于上层土壤,下层土壤根系生物量分布越少且变化趋于平缓。
猜你喜欢
石油库与加油站(2022年3期)2022-08-30
散文诗(2021年22期)2022-01-12
矿冶工程(2021年4期)2021-09-15
世界有色金属(2020年15期)2020-10-10
环球时报(2019-05-23)2019-05-23
浙江工业大学学报(2017年5期)2018-01-22
中国食用菌(2017年2期)2017-03-28
江苏农业科学(2016年11期)2017-03-21
科技视界(2016年7期)2016-04-01
郑州大学学报(工学版)(2014年6期)2014-03-01
