我国辣椒资源pvr2-eIF4E基因的多样性分析
2021-05-26 13:09曹亚从顾晓振张正海于海龙冯锡刚张南南张伟丽景雅欣王海平李锡香BenoMOURY张宝玺王立浩
辣椒杂志 2021年1期
曹亚从 顾晓振 张正海 于海龙 冯锡刚 张南南 张伟丽景雅欣 王海平 李锡香 Benoît MOURY 张宝玺 王立浩*
(1. 中国农业科学院蔬菜花卉研究所, 北京 100081; 2. 法国农业科学院植物病理研究所,阿维尼翁 84143)
辣椒(Capsicum spp.)属于茄科辣椒属,起源于南美洲。1493年,哥伦布首次将辣椒引入欧洲,并于15至16世纪后期引入东亚[1],由于复杂的地理环境以及人们的驯化等活动,形成了丰富的辣椒种质资源。遗传多样性使育种者可以利用种质资源中不同的等位基因来培育品种,等位基因挖掘已经被用来在已知基因座上发现未探索的等位基因多样性,是利用资源多样性育种的基础。
我国是世界上辣椒种植面积最大的国家(http://www.FAO.org)。自2015年以来,辣椒已成为我国播种面积最大的蔬菜作物,年播种面积150万 ~200万hm2[2]。马铃薯Y病毒属包括多个种,是一种世界性的病害。1956年Simons首次报道在美国加州有PVY侵染辣椒[3]。在中国,20世纪80年代马铃薯Y病毒属对辣椒的危害就有广泛报道[4-6]。近年来报道危害辣椒的马铃薯Y病毒属病毒有马铃薯Y病毒(Potato Virus Y,PVY)、烟草蚀纹病毒(Tobacco Etch Virus,TEV)、辣椒斑驳病毒(Pepper Mottle Virus,PepMoV)、辣椒脉斑病(Pepper Veinal Mottle Virus,PVMV)等[7-14],严重影响辣椒的生产效益,尤其在高温地区,是辣椒的重要病害,其危害仅次于CMV和TMV[15-16]。
eIF4E是植物对抗病毒的重要决定因子[17-20],酵母双杂交及ELISA实验表明病毒基因组连接蛋白(VPg)或其前体(NIa)可以与eIF4E或其同源异构体Eif(iso)4E相互作用,而互作缺失的病毒则丧失侵染性[21-22]。在辣椒中同样发现了与马铃薯Y病毒属病毒PVY和TEV抗性相关的eIF4E[23-25],根据发现的抗性及发现时间先后顺序,Kyle和Palloix[26]将其命名为pvr2,后证明pvr1与pvr2为等位基因。pvr2-eIF4E序列多样性丰富,感病等位基因为pvr2+,继pvr2 1和pvr2 2被发现后[24,27-29],Charron等[30]对25份辣椒材料 (Capsicum annuumL.)的研究发现了pvr2的10个等位基因,在此10个等位基因中,有7个前人未报道,分别命名为pvr2 3-9。Ibiza等[31]采用EcoTILLING的方法,在233个辣椒栽培品种中发现了16个pvr2的等位基因,其中有13个等位基因前人未报道,分别命名为pvr2 10-22。pvr2-eIF4E序列的多样性导致其功能差异,体现在对不同PVY小种(PVY0、PVY0,1、PVY0,1,2)[29]和TEV的抗性差异。
对pvr2-eIF4E的多态性研究发现,其外显子的多态性比例高于内含子[30],因此pvr2-eIF4E的研究重点位于其外显子。pvr2-eIF4E基因具有5个外显子,研究表明外显子1的多态性最丰富[30-31],因此本课题组前期对外显子1的多态性进行了研究[32],在1 904份材料中共发现17个单倍型,14个有义突变位点。
我国有着丰富的辣椒种质资源,但是pvr2-eIF4E作为辣椒抗PVY和TEV育种工作中有效而稳定的抗源,并未在丰富的资源中得到广泛而深入的研究。本研究通过全面鉴定pvr2-eIF4E的所有外显子,挖掘出我国辣椒资源中的PVY和TEV抗性材料,为PVY和TEV抗性材料的挖掘和利用提供研究基础。
1 材料与方法
1.1 材料
我国种质资源中期库1 863份辣椒材料,其中187份材料引种自国外,引种地区包括美洲、欧洲、非洲、亚洲的26个国家;1 676份材料来自于中国33个省、市、自治区。
1.2 基因组DNA提取
辣椒材料播种于穴盘,待长到约6片真叶时,取嫩叶片,采用改良CTAB方法提取叶片基因组DNA[33]。采用Biospec-nano显微分光光度计(Shimadzu Corporation)测定DNA的浓度及质量。基因组DNA稀释到25 ng/L备用。
1.3 扩增引物设计
通过美国国家生物技术信息中心NCBI网站(http://www.ncbi.nlm.nih.gov/)公布的Yolo Wonder的pvr2-eIF4E的编码序列确定目的基因序列(AY122052.1)。将序列比对到CM334 v1.55版 本 基 因 组[34](http://passport.pepper.snu.ac.kr/?t=PGENOME),得到pvr2-eIF4E在CM334基因组中的基因为CA04g00860。
利用在线工具Primer 3(http://primer3.ut.ee/)设计引物扩增外显子,扩增引物见表1。
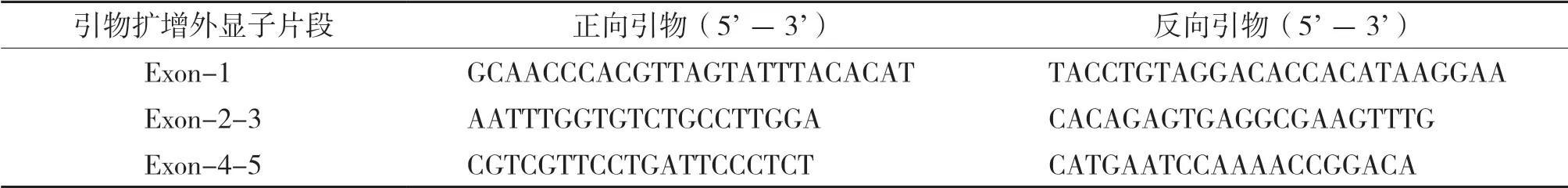
表1 pvr2-eIF4E外显子测序引物序列Table 1 Sequences of pvr2-eIF4E exon sequencing primers
1.4 产物扩增及测序
以叶片基因组DNA为模板,分别对外显子1、外显子2-3和外显子4-5进行扩增,采用北京全式金生物技术有限公司的DNA高保真聚合酶试剂盒(TransStart ® Top taq DNA Polymerase),扩增体系为30 μL:2 μL 基因组DNA,0.6 μL高保真聚合酶,3 μL 缓冲液(10 X),2.4 μL dNTP,上下游引物各1.2 μL,19.6 μL ddH2O。PCR反应程序为:94 ℃ 3min;94 ℃ 30 S,58 ℃ 30 S,72 ℃ 60 S,35 个循环;72 ℃ 10 min。PCR产物送上海生工生物工程有限公司进行Sanger测序,测序引物采用扩增正向引物。对含有新发现等位基因的材料进行3次重复测序。
1.5 测序结果分析
将测序结果在Cexpress软件中打开,序列对齐后根据Yolo Wonder的编码序列,确定各材料的碱基突变位点,用MEGA5.0对测序序列进行多重比对,统计各材料间氨基酸差异位点。
1.6 PVY抗性接种鉴定
对含有新发现等位基因的材料进行PVY抗性接种鉴定。鉴定工作委托法国农业科学院完成,接种PVY病毒株系为LYE84.2和SON41,其中LYE84.2为PVY0,SON41为PVY0,1,2。
1.7 PVY抗性材料遗传关系分析及核心种质构建
利用Gu等[35]SSR数据,提取含有抗性等位基因材料的数据,将其转化为矩阵,通过DARWin软件(https://darwin.cirad.fr/)对含有抗性等位基因的材料构建进化树。
根据进化树将材料进行分组,对每种pvr2等位基因在每个组中提取遗传距离最远的两份材料,分组中少于2份的,则提取实际材料数。
2 结果与分析
2.1 pvr2-eIF4E基因序列多样性分析
在PCR扩增测序的1 863份辣椒材料中,发现的变异均为SNP,共检测到10个有义氨基酸突变位点(表2),共检测到10种等位基因,其中有1 117份材料与感病材料Yolo Wonder有相同的氨基酸序列,为pvr2+;有8种等位基因与前人已报道等位基 因pvr2 1、pvr2 2、pvr2 3、pvr2 4、pvr2 6、pvr2 7、pvr2 9、pvr1 +编码序列一致,共744份材料;有1份材料为新发现的等位基因,命名为pvr223,在第1外显子的第185位碱基发生突变,导致编码苯丙氨酸的密码子(UGG)突变为终止密码子(UAG)(图1)。以感病材料Yolo Wonder编码区序列为对照,1 863份材料中共有746份发生有义氨基酸突变,约占总数的41%。
突变位点的统计结果显示(表2),突变位点主要位于第1外显子,有7个位点发生有义突变,在第2、3、4外显子分别有1个有义突变,在第5外显子未发现有义突变。突变位点最多的位点位于第1外显子的第67位氨基酸,有644份材料发生有义突变;其次为第205、71、79位氨基酸,分别有232、97、20份材料发生有义突变,其余位点发生突变的材料很少。
2.2 pvr2-eIF4E等位基因抗性分析
在1 863份辣椒材料发现的10种pvr2-eIF4E等位基因中,已知有2种等位基因为感病,7种等位基因对PVY和TEV部分小种具有抗性(表2),1种等位基因的抗性未知。

表2 pvr2-eIF4E基因多样性分析Table 2 Analysis of pvr2-eIF4E gene diversity
采用PVY株系LYE84.2(PVY0)和SON41(PVY0,1,2),对含有新发现等位基因pvr223的辣椒材料进行PVY抗性接种鉴定。结果显示,含有pvr223的辣椒材料易感此2种PVY株系(表3)。

表3 人工接种鉴定pvr223等位基因材料对PVY抗性的统计数据Table 3 Summary on the resistances of materials with pvr223 allele to PVY measured by artificial inoculation
在研究的1 863份辣椒材料中,共有649份材料含有pvr2抗性等位基因,占总材料数的34.84%。649份材料均对PVY0具有抗性,占总材料数的34.84%;其中有6份材料含有高抗PVY等位基因(pvr2 6、pvr2 7、pvr2 9),对PVY0,1,2具有抗性,占总材料数的0.32%;有1份材料含有已报道抗TEV的等位基因pvr2 2。
2.3 含有抗PVY的pvr2-eIF4E等位基因材料地理分布
抗性材料主要来源于我国国内,有586份,占抗性材料的90.29%;有63份抗性材料来源于国外,占抗性材料的9.71%(图2)。国内的抗性材料主要来源于湖北,有134份,占国内抗性材料的22.87%,其次来自于湖南、云南和四川,分别有52、49和42份,占国内抗性材料的8.87%、8.36%和7.17%。收集自国外的抗性材料主要来源于美国,有25份,占国外抗性材料的39.68%,其次来自于泰国、印度和荷兰,分别有9、5和4份,占国外抗性材料的14.29%、7.94%和6.35%。
含有高抗PVY等位基因(pvr2 6、pvr2 7、pvr2 9)的材料分别来自于中国山西(1份)、美国(2份)、墨西哥(1份)、西班牙(1份),另有1份引自国外,引进地不详。含有已报道抗TEV的pvr22材料来自英国。

图2 含有抗PVY等位基因材料的地理分布图 A,在不同国家之间的分布;B,在中国各地的分布Figure 2 Geographical distribution of materials containing PVY-resistant alleles. A, distribution among diあerent countries;B, distribution throughout China.
2.4 含有抗PVY的pvr2-eIF4E等位基因材料遗传关系分析
基于SSR标记的分型数据,对649份辣椒抗性材料的遗传关系进行了分析(图3)。根据遗传关系可以将含有pvr2抗性等位基因的材料分为3个群,如图3所示,其中群I包含本研究中鉴定到的所有抗性等位基因,又分为3个亚群;群II包含4种抗性等位基因(pvr2 1、pvr2 3、pvr2 4、pvr2 9),又分为3个亚群;群III包含3种抗性等位基因(pvr2 1、pvr2 3、pvr2 4),又分为4个亚群。
2.5 PVY抗性核心种质构建
结合SSR标记分型数据构建的遗传关系和pvr2等位基因,抽取PVY抗性核心种质,每个亚群中抽取每个pvr2等位基因2份,抽取在进化树上遗传距离最远的2份,少于2份的则抽取实际数据材料。PVY抗性核心种质材料共抽取53份,具体见表4。
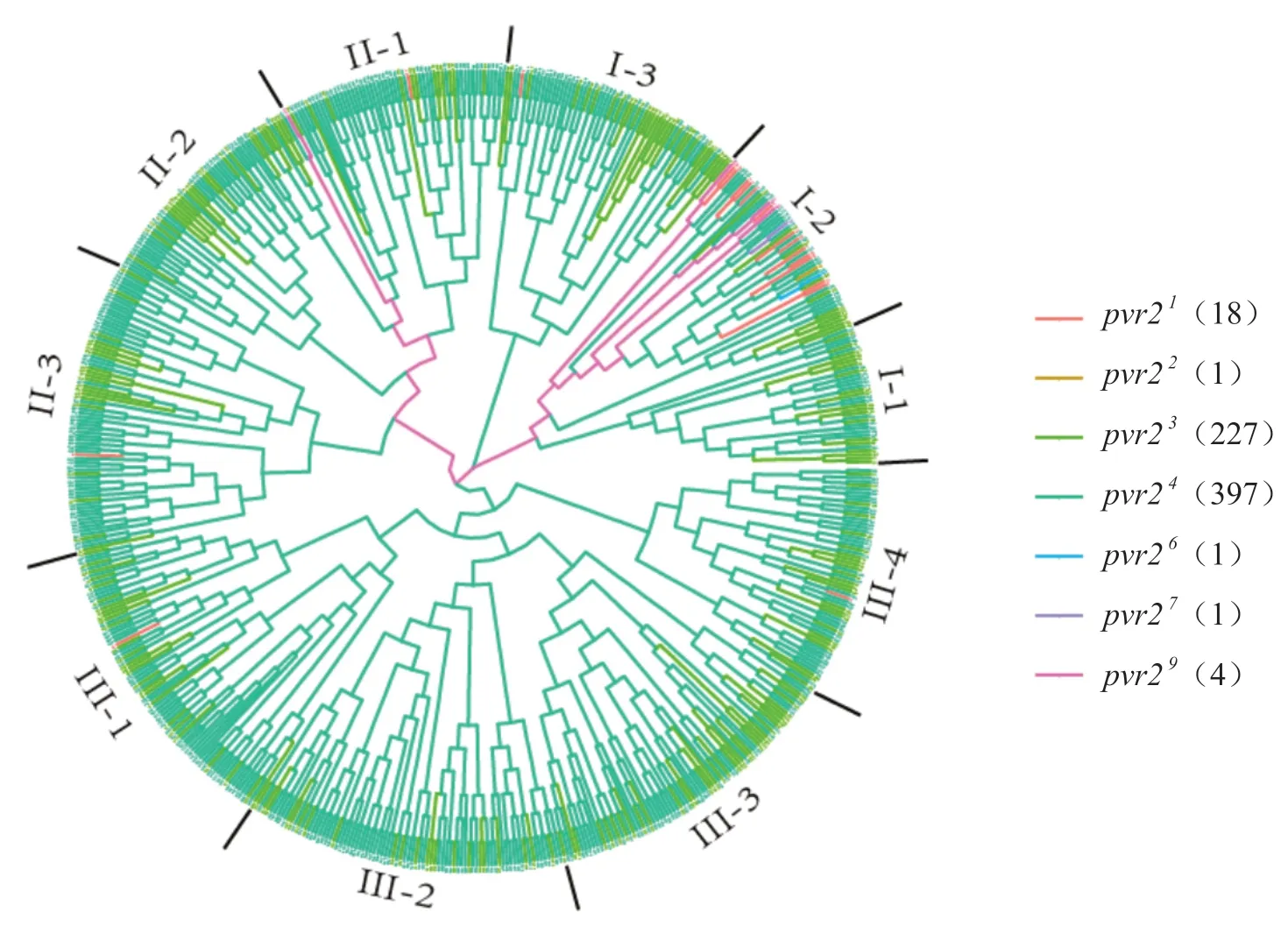
图3 含有抗性pvr2等位基因材料的遗传关系图Fig. 3 Genetic relationship of accessions containing resistant pvr2 alleles
3 讨论
pvr2-eIF4E基因产生抗性的确切机制仍有待阐明,对pvr2基因表达数据的分析表明,植物感病与否的表型差异是由pvr2氨基酸变化决定的,而不是因为其他调节eIF4E表达的因子决定[40]。已有报道的蛋白互作研究认为,pvr2-eIF4E基因发挥抗性作用,是由于病毒Vpg蛋白和植物eIF4E蛋白的互作被破坏,病毒Vpg不能结合到eIF4E,从而影响病毒的复制、移动等[36,40]。通过结构模型预测,认为pvr2的突变位点位于蛋白的表面,第62~79位氨基酸被定义为蛋白结构域I,第106~109位氨基酸被定义为蛋白结构域II,这两个区域可能是pvr2-eIF4E与PVY和TEV互作的关键区域,参与对PVY的抗性[41-42]。本研究发现的新等位基因pvr223的突变发生在第1外显子的第62位氨基酸,由苯丙氨酸突变为终止密码子,使原本含228个氨基酸的蛋白突变为仅有62位氨基酸的蛋白。虽然新发现等位基因pvr223仅保留了很短的蛋白序列,保留的序列不存在前人报道的与Vpg互作的候选关键区域,但仍然感病,因此pvr2-eIF4E可能存在其他抗病相关作用位点及作用机制,或含有pvr2 23-eIF4E的材料存在其他基因编码与PVY病毒Vpg蛋白互作的蛋白。
辣椒传入我国较晚,遗传背景比较狭窄,本研究鉴定的辣椒资源多收集于“七五”、“八五”期间,多数未利用多抗性材料经过抗性转育,因此,虽然我国有大量含PVY抗性等位基因的材料,但是多样性不够丰富,多抗性差:含有抗多个PVY小种的基因型等位基因pvr2 6、pvr2 7、pvr2 9材料仅有6份,且有5份是引种自国外;含有已报道抗TEV基因型等位基因pvr2 2的材料仅有1份。近年来,各地多有报道PVY和TEV病毒在辣椒上引起的危害[7-9,11,13,15,43-44],我国现存种质资源中缺少PVY和TEV高抗材料,需要引起重视,今后要加强引入丰富的pvr2抗性等位基因材料,用以培育丰富类型的多抗PVY和TEV辣椒品种。
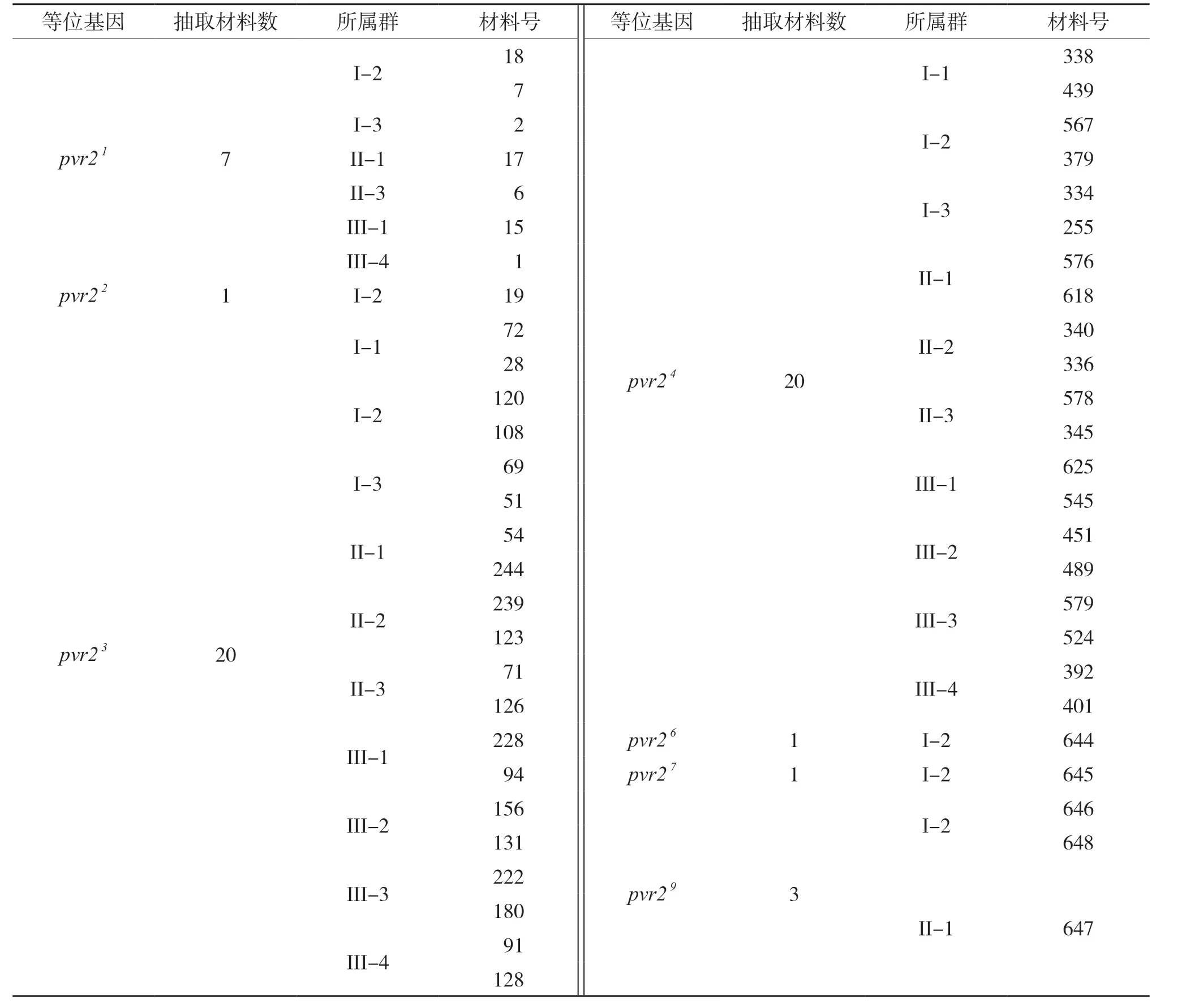
表4 PVY抗性核心种质材料抽取情况Table 4 Extraction of PVY resistant core germplasm
猜你喜欢
分子诊断与治疗杂志(2022年9期)2022-10-09
当代水产(2022年7期)2022-09-20
西南农业学报(2022年3期)2022-04-25
植物保护(2019年2期)2019-07-23
新课程·下旬(2018年9期)2018-11-14
中国医药导报(2018年14期)2018-08-30
青少年科技博览(中学版)(2015年10期)2015-01-11
湖北农业科学(2014年11期)2014-09-10
现代农业科技(2009年19期)2009-03-20
中学生物学(2008年6期)2008-08-29