
豌豆蚜翅分化相关miRNA及其预测靶基因对蜕皮激素的应答及miR-92a-1-p5靶基因的验证
2021-05-25 04:07马田田杨宗霖常美玲霍春月阚云超李丹丹
昆虫学报 2021年4期
马田田, 杨宗霖, 常美玲, 霍春月, 阚云超, 李丹丹
(南阳师范学院, 河南省伏牛山昆虫生物学重点实验室, 河南南阳 473061)
蚜虫是农林植物常见害虫类群之一,在世界各地广泛分布(Blackman and Eastop, 2000)。蚜虫具复杂生活周期,在春季和夏季以卵胎生方式孤雌生殖,秋季随着光周期变化和温度下降,群体中出现两性生殖个体,其交配产卵后以卵形式越冬,至第二年春季孵化,继续营孤雌生殖(Margaritopoulos and Tsitsipis, 2002; Braendleetal., 2006)。孤雌蚜在发育过程中具翅多型性,包括非遗传多型性和遗传多型性(Roff, 1986)。翅多型性是蚜虫适应环境变化的结果,在某个发育阶段环境中出现的敏感因素可造成翅原基的形成。如增加蚜虫密度或降低营养供应,会造成孤雌生殖后代出现更多的有翅个体(Lees, 1967; Muller, 2001; Ishikawaetal., 2012)。其他环境因素,如种间竞争及激素也会对蚜虫的翅型分化产生影响(Ishikawaetal., 2013),由回避捕食者效应触发的触觉刺激增加也会诱导更多有翅后代的产生(Kunert and Weisser, 2005)。而共生取食者蚂蚁(取食蚜虫蜜露)增加则会抑制有翅个体的产生(Kleinjan and Mittler, 1975; Pringleetal., 2014)。最近研究表明,Wnt2,Fng,Uba1,Hh,Foxo,Dpp,Brk,Ap,Dll,Hth,Tsh,Nub,Scr,Antp,Ubx,Asc,Srf和F1等基因在禾谷缢管蚜Rhopalosiphumpadi的翅多态性分化中发挥重要调控作用(Zhang RJetal., 2019)。同时,研究表明miRNA在蚜虫的翅二型性分化中也发挥重要作用,通过RNA-Seq在麦长管蚜Sitobionavenae中鉴定出345个miRNA,其中16个miRNA在有翅孤雌个体中的表达显著上调,而12个miRNA在无翅孤雌个体中积累更多;差异表达的miRNA靶基因可归结在124种代谢通路上,包括Wnt, Notch, Hedgehog和TGF-beta信号途径等(Lietal., 2016)。同时,Vellichirammal等(2017)发现在母代注射蜕皮激素或其类似物会导致后代有翅比例下降,表明蜕皮激素在蚜虫翅型分化中也发挥重要作用。但是蜕皮激素是否影响miRNA表达,是否通过miRNA参与蚜虫翅型分化目前还不清楚。
因此,为进一步探索蜕皮激素及miRNA在豌豆蚜Acyrthosiphonpisum翅型分化中是否存在互作,我们选择在前期工作中发现的5个参与孤雌生殖豌豆蚜翅型分化的关键miRNA,使用蜕皮激素类似物20-羟基蜕皮酮(20E)处理其若虫,研究蜕皮激素对miRNA及其靶基因表达的影响。同时通过荧光素酶活性实验及个体RNA干涉实验,对miRNA及靶基因的互作情况进行深入探索,为从蜕皮激素与miRNA互作角度探索孤雌蚜翅型分化的分子机制,为蚜虫综合防治提供理论依据。
1 材料与方法
1.1 试虫
所用豌豆蚜由中国科学院动物研究所崔峰研究员惠赠。孤雌生殖豌豆蚜培养方式参照杨宗霖等(2019)的方法:将蚕豆种子置于25℃浸泡12 h;将东北黑土与蛭石以1∶2(v/v)混合后播种,在温度28℃,相对湿度60%,光周期14L∶10D,光照强度15 000 lx条件下培养。待蚕豆展开两片真叶时移入塑料杯(直径8 cm,高14 cm)中,用毛笔挑取豌豆蚜1龄若蚜5头置于恒温气候培养箱(温度20℃,相对湿度60%,光周期16L∶8D,光照强度15 000 lx)中培养,低密度连续筛选两代至后代均为孤雌无翅蚜后用于实验。
1.2 蜕皮激素处理
将25头孤雌生殖豌豆蚜2龄若虫分别浸入0.1 mol/L 20E和4.5%的乙醇(20E溶剂,用作阴性对照)中10 min和30 min,然后转移至种植有蚕豆苗的塑料杯(直径8 cm,高度为14 cm)中培养48 h(对照与处理从发育阶段上无显著差异),收集蚜虫整虫在液氮中迅速冷冻,并在-80℃下保存,TRIzol法提取总RNA。
1.3 qPCR检测miRNA及靶基因的表达谱
选择在前期工作中发现的5个参与孤雌生殖豌豆蚜翅型分化的关键miRNA(Let-7, miR-92a, miR-92b, miR-92a-1-p5和miR-277),使用miRanda (Enrightetal., 2003)和RNAhybrid (Rehmsmeieretal., 2004)软件预测这些miRNA的靶基因分别为abrupt,wingless,Foxo,flightin和Uba1(杨宗霖等, 2019)。miRNA及靶基因表达谱检测同杨宗霖等(2019)方法:分别取1.2节提取的各样品总RNA 2 μg,使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)和All-in-OneTMmiRNA qRT-PCR Detection Kit(GeneCopoeia)合成cDNA第1链。内参基因为actin。靶基因PCR反应体系(10 μL): cDNA 0.5 μL, 上下游引物终浓度为0.5 μmol/L, FastStart Universal SYBR Green Mix (Roche) 5 μL。PCR反应条件: 95℃变性30 s, 55℃退火30 s, 72℃延伸30 s, 40个循环。miRNA PCR反应体系(10 μL): 5倍稀释的cDNA 1 μL, miRNA上游特异引物终浓度0.5 μmol/L, 下游引物为试剂盒通用引物Universal Adaptor PCR Primer终浓度0.5 μmol/L, 2×All in one qPCR Mix 5 μL。PCR反应条件: 95℃变性10 s, 55℃退火20 s, 72℃延伸30 s, 40个循环。内参基因为U6。所用引物见表1。qPCR实验设置生物学重复3次,每样品技术重复3次。
1.4 miR-92a-1-p5靶基因flightin的体外验证
1.4.1靶基因荧光素酶报告基因载体构建:以杨宗霖等(2019)有翅豌豆蚜4龄若虫RNA为模板,使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA第1链,使用表1引物扩增flightin基因CDS 593 bp序列。 PCR反应体系(20 μL): cDNA 1 μL, 上下游引物终浓度为0.5 μmol/L,Premix Ex TaqTMHot Start Version (TaKaRa) 10 μL。PCR反应条件: 95℃变性30 s; 55℃退火30 s, 72℃延伸1 min, 30个循环。使用琼脂糖凝胶DNA回收试剂盒(Axygen)回收目的条带, 连接pMD19-T Simple载体(TaKaRa)。转化后,菌液PCR检测连接效果,送菌液至苏州泓迅生物科技股份有限公司测序。测序正确的菌液提质粒,用XhoⅠ和XbaⅠ双酶切后连接在pmirGLO双荧光素酶报告基因载体(Promega)上。经转化、菌液PCR检测、测序等步骤确定载体是否构建成功。测序正确的菌液重新摇菌,用去内毒素质粒提取试剂盒(Omega)提取质粒。

表1 引物信息Table 1 Primers used in this study
1.4.2miRNA mimics合成:依据miR-92a-1-p5成熟miRNA序列,由上海吉玛制药技术有限公司合成miR-92a-1-p5模拟物(miR-92a-1-p5 mimics)、阴性对照(NC mimics)和miR-92a-1-p5突变模拟物(miR-92a-1-p5 mutant mimics)(引物序列见表1)。
1.4.3细胞转染及荧光素酶活性测定:HEK293T细胞(杨宗霖等, 2019)在含有10%胎牛血清(Gibco)的DMEM培养基(Biological Industry)中25 cm2培养瓶传代培养,温度37℃,CO2浓度5%。按每孔2×105个细胞铺24孔板,使用X-tremeGENE siRNA Transfection Reagent(Roche)转染miRNA mimics和靶基因荧光素酶报告基因载体。miRNA mimics转染浓度为100 nm/孔,质粒转染浓度为100 ng/孔。实验设置3组:共转染pmirGLO[flightin]与NC mimics;共转染pmirGLO[flightin]与miR-92a-1-p5 mimics;共转染pmirGLO[flightin]与miR-92a-1-p5 mutant mimics。每组设样品重复3个,实验重复3次。转染48 h后,用细胞裂解液裂解细胞,取20 μL细胞裂解液,按照双荧光素酶报告基因检测试剂盒(Promega)步骤,检测样品荧光值。靶基因位于Firefly荧光素酶基因下游,因此用获得的Firefly荧光素酶活性(F)/内参Renilla荧光素酶活性(R),获得每组3个样品及3个重复的R/F值,均一化后用Excel软件进行作图(杨宗霖等, 2019)。
1.5 miR-92a-1-p5靶基因flightin的体内验证
1.5.1miR-92a-1-p5 antagomir合成:依据miR-92a-1-p5成熟miRNA序列,由上海吉玛制药技术有限公司合成miR-92a-1-p5 antagomir(引物序列见表1)。
1.5.2纳米载体/miR-92a-1-p5 antagomir/变性剂复合物的制备: Zheng等(2019)和Yan等(2021)开发的纳米载体/变性剂复合物可以渗透蚜虫体壁,使编码基因在蚜虫个体中实现很好的RNA干涉效果。因此,使用该方法在蚜虫个体中尝试miRNA干涉效果。将miR-92a-1-p5的antagomir与纳米载体以1∶1体积比混合,加入0.1%体积的变性剂,将混合物在室温放置15 min,形成纳米载体/miR-92a-1-p5 antagomir/变性剂复合物。使用相同量的antagomir NC(上海吉玛制药技术有限公司设计合成)作为阴性对照。
1.5.3纳米载体/miR-92a-1-p5 antagomir/变性剂复合物在蚜虫个体中的使用: 选择孤雌生殖豌豆蚜4龄若虫,将50 nL纳米载体/miR-92a-1-p5 antagomir/变性剂复合物通过微量注射器点在蚜虫背板上,将蚜虫分开放置,直到液滴完全吸收为止。将蚜虫转移至种植有蚕豆苗的塑料杯中,1.1节条件下正常培养48 h,收集整虫液氮快速冷冻,TRIzol试剂提取RNA,使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA第1链,qPCR检测蚜虫个体中miRNA干涉效果和靶基因表达量变化,反应体系及条件同1.3节。
1.6 数据分析
数据分析同杨宗霖等(2019)方法。PCR结果采用2-△△Ct法计算相对表达量高低(Ct为循环阈值,表示每一个反应管内的荧光信号到达设定的阈值时所经历的循环数,△Ct=目的基因的平均Ct值-内参基因的平均Ct值,△△Ct=△Ct样本1-△Ct样本2(样本1为处理样本,样本2为对照样本)。实验独立生物学重复3次,每样品测定技术重复3次,数据为3次独立生物学实验平均值分析结果。数据采用SPSS Statistics 22.0软件进行处理,以平均值±标准误表示,并采用单因素方差分析和t检验进行检测不同处理间的差异显著性。
2 结果
2.1 miRNA对20E处理的表达响应
qPCR检测结果表明,20E处理30 min可极显著诱导5个miRNA的表达(P<0.01) (图1)。其中20E处理10 min和30 min后蚜虫中Let-7和miR-277表达量均极显著上升(P<0.01) (图1: A, C)。但处理10 min情况下,miR-92a-1-p5和miR-92b表达量较对照极显著下降(P<0.01) (图1: D, E),仅在处理30 min样品中有极显著上升(P<0.01) (图1: D, E),表明不同miRNA响应蜕皮激素的时间点也存在差别。

图1 qPCR检测5个miRNA在0.1 mol/L 20E处理后的孤雌生殖豌豆蚜2龄若虫中的表达谱Fig. 1 Expression profiles of five miRNAs in the 2nd instar nymphs of the parthenogeneticAcyrthosiphon pisum after exposed to 0.1 mol/L 20E detected by qPCRA: Let-7; B: miR-92a; C: miR-277; D: miR-92a-1-p5; E: miR-92b. Mock: 阴性对照(4.5%的乙醇)Negative control (4.5% ethyl alcohol); 20E: 20E处理20E treatment. 以U6作内参基因。图中数据为平均值±标准误;柱上双星号表示处理与对照间差异极显著(P<0.01, 单因素方差分析, 双尾配对t检验)。U6 was used as the reference gene. Data in the figure are mean±SE. Double asterisk above bars represents statistically extremely significant difference (P<0.01, one-way ANOVA, two-tailed, paired t-test) between the treatment and the control. 下同The same below.

图2 qPCR检测miRNA靶基因在0.1 mol/L 20E处理后的孤雌生殖豌豆蚜2龄若虫中的表达谱Fig. 2 Expression profiles of miRNA target genes in the 2nd instar nymphs of the parthenogeneticAcyrthosiphon pisum after exposed to 0.1 mol/L 20E detected by qPCRA: abrupt; B: wingless; C: Uba1; D: flightin; E: Foxo. 以actin作内参基因。actin was used as the reference gene.
2.2 miRNA预测靶基因对20E的表达响应
qPCR检测结果表明,在20E处理后,Let-7的预测靶基因abrupt在不同处理时间下表达量均降低,与Let-7呈相反的表达模式(图2: A)。wingless和Uba1在20E处理10 min时表达量极显著下降(P<0.01),与miR-92a和miR-277表达趋势(图1: B, C)相反,而在30 min处理下miRNA与靶基因表达趋势一致,均被20E极显著诱导(P<0.01) (图2: B, C)。miR-92a-1-p5的预测靶基因flightin在20E处理30 min后表达量极显著下降(P<0.01),与miRNA表达趋势相反,而在10 min处理情况下二者均下降(图2: D)。miR-92b与预测靶基因Foxo在20E诱导后并不存在表达模式上的明显相关性(图2: E)。表明不同的miRNA与预测的靶基因在不同的处理时间点并不具有完全相反的表达趋势。
2.3 miR-92a-1-p5与靶基因flightin互作的体外验证
蚜虫翅的发生关键是飞行肌的产生,为了进一步验证flightin基因与miR-92a-1-p5是否存在互作,我们利用双荧光素酶报告基因系统检测miRNA与预测的靶基因之间的互作关系。首先构建靶基因flightinCDS区荧光素酶报告基因载体pmirGlO [flightin],flightinCDS扩增结果如图3(A)所示,PCR产物电泳条带单一,593 bp位置正确。pmirGlO [flightin]双酶切结果如图3(B),能够酶切出正确的flightinCDS条带,测序结果正确。共转染miR-92a-1-p5模拟物和flightinCDS过表达载体pmirGlO [flightin]后,细胞荧光素酶活性比共转染miR-92a-1-p5突变模拟物和pmirGlO [flightin]细胞的下降了32%,与转染对照NC mimics相比下降40%,达极显著水平(P<0.01) (图3: C),表明miR-92a-1-p5对flightin基因的转录存在调控。
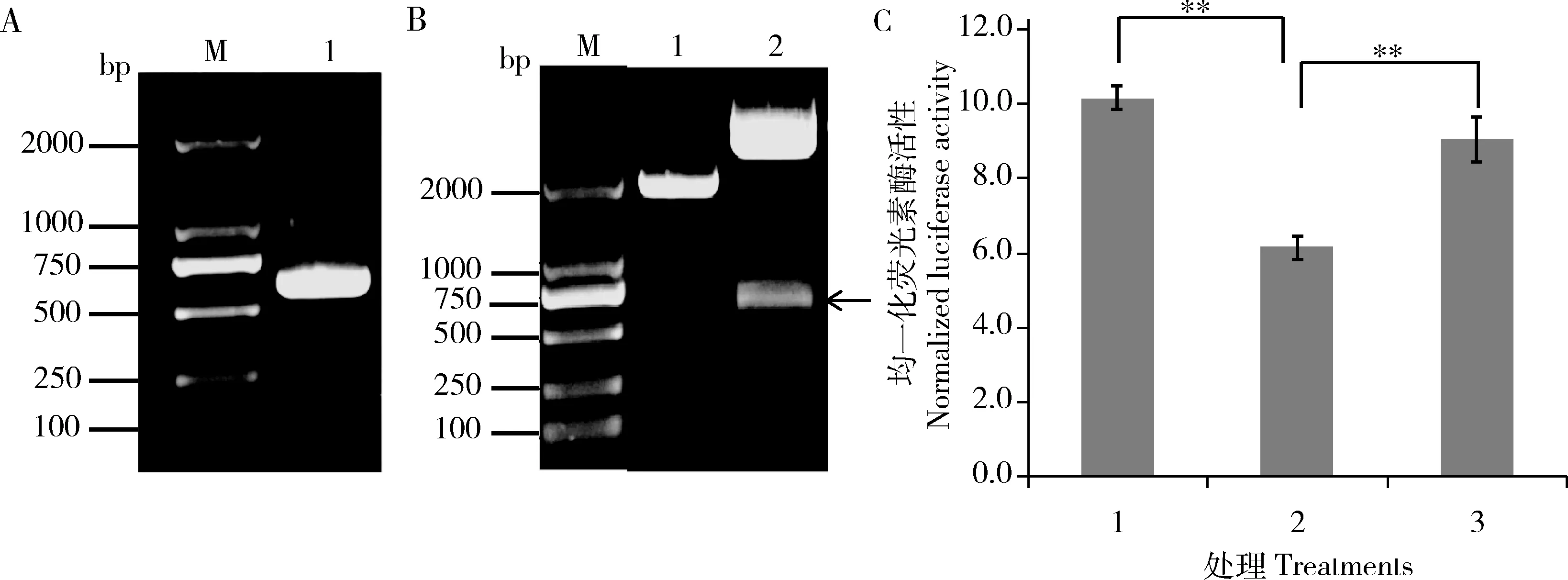
图3 荧光素酶活性试验检测miR-92a-1-p5与靶基因flightin互作Fig. 3 Verification of target gene flightin of miRNA miR-92a-1-p5 by luciferase activity analysisA: flightin CDS扩增结果Amplification of flightin CDS. M: DL2000 marker; 1: flightin CDS扩增产物Amplification product of flightin CDS. B: 载体pmirGlO [flightin]双酶切鉴定Double digestion result of the vector pmirGlO[flightin]. 1: pmirGlO双酶切产物Double digestion product of pmirGlO; 2: pmirGlO [flightin]双酶切产物Double digestion product of pmirGlO [flightin]. 箭头示酶切获得的flightin CDS 593 bp序列。Arrow shows the digested 593 bp sequence of flightin CDS. C: HEK293细胞系中荧光素酶活性试验检测miR-92a-1-p5的靶基因flightin (Verification of target gene flightin of miRNA miR-92a-1-p5 by luciferase activity analysis in HEK293 cells). 1: pmirGlO [flightin]+NC mimics; 2: pmirGlO [flightin]+miR-92a-1-p5 mimics; 3: pmirGlO [flightin]+miR-92a-1-p5 mutant mimics.
2.4 miR-92a-1-p5靶基因flightin的体内验证
miR-92a-1-p5 qPCR检测结果表明,纳米载体/miR-92a-1-p5 antagomir/变性剂复合物渗透豌豆蚜4龄若蚜后,miR-92a-1-p5的表达水平受到极显著抑制(P<0.01),表达量下降了83%(图4: A);而flightin的表达量却增加了48%(P<0.01) (图4: B),与miR-92a-1-p5呈现相反的表达模式。结果进一步证明了miR-92a-1-p5对flightin基因的转录存在调控,flightin是miR-92a-1-p5的靶标。
3 讨论

图4 孤雌生殖豌豆蚜4龄若虫中敲低miR-92a-1-p5后miR-92a-1-p5 (A)与flightin (B)的表达量变化Fig. 4 Changes in the expression levels of miR-92a-1-p5 (A) and flightin (B) after knocking down miR-92a-1-p5in the 4th instar nymphs of parthenogenetic Acyrthosiphon pisumNC: 阴性对照Negative control (antagomir NC); Antagomir: miR-92a-1-p5抑制物Inhibitor of miR-92a-1-p5.
昆虫的翅多态性是理解自然群体中扩散的适应性最有吸引力的模型,有翅个体具有完整的翅和飞行肌,可以长距离迁移以适应恶劣环境并寻找新的栖息地,但无翅个体则缺乏翅和飞行肌,并在相对受限的区域生存(Briceno and Eberhard, 1987; Ogawa and Miura, 2013; Zhang CXetal., 2019)。从环境因素来讲,饲养密度,寄主植物营养状况,温度和光周期等环境因素触发了蚜虫翅型的转换(White, 1946; Kunert and Weisser, 2003; Kunert and Weisser, 2005; Brisson, 2010; Castanedaetal., 2010; Zhangetal., 2015)。从分子水平上,蜕皮激素、胰岛素信号通路及miRNA等在蚜虫翅多型性演化中都发挥了至关重要的作用(Guoetal., 2016; Lietal., 2016; Vellichirammaletal., 2017; Zhang CXetal., 2019; Zhang RJetal., 2019)。蜕皮激素是一类具有昆虫蜕皮活性的天然类固醇,孤雌蚜母代注射蜕皮激素或其类似物会导致其所产有翅后代比例降低(Vellichirammaletal., 2017)。但是蜕皮激素如何在孤雌蚜翅型分化中发挥作用目前还不清楚。在这里,我们发现蜕皮激素类似物20E可以诱导Let-7, miR-92a, miR-277, miR-92a-1-p5和miR-92b的表达,可抑制飞行肌基因flightin的表达。同时miR-92a-1-p5可调控flightin基因的转录,表明flightin为miR-92a-1-p5的真实靶标。由于蚜虫对农作物的主要损害是由于有翅孤雌蚜的迁徙,因此了解孤雌蚜的翅分化调控机制将为蚜虫防治提供新的线索,而miR-92a-1-p5与flightin之间的互作也为孤雌蚜的防治提供了新的农药靶点。
在昆虫个体中抑制基因表达可采用直接注射或饲喂双链RNA的方法进行,个体直接注射法在黑腹果蝇Drosophilamelanogaster(Kennerdell and Carthew, 1998)、家蚕Bombyxmori(Lietal., 2015; Zhouetal., 2017)和桃蚜Myzuspersicae(Cheng and Li, 2018)等很多物种中都成功实现。但蚜虫个体小、体壁薄,采用显微注射法死亡率较高。Cao等(2018)和Pitino等(2011)用饲喂法也在蚜虫中实现了部分基因的RNA干涉,但操作步骤繁琐,效率有限。最近Zheng等(2019)和Yan等(2021)用纳米载体/变性剂在大豆蚜Aphisglycines中导入dsRNA实现了编码基因的RNA干涉,干涉效率可达到95%以上,取得了较好的效果,但是该方法未在非编码基因上进行尝试。在本研究中我们利用纳米载体/变性剂在豌豆孤雌蚜中成功导入了miRNA miR-92a-1-p5抑制物,干涉效率达83%,表明该法不仅适用于编码基因,对非编码基因同样可以实现较好的干涉效果。
猜你喜欢
小天使·一年级语数英综合(2022年5期)2022-05-25
发明与创新·小学生(2021年5期)2021-08-23
天津医科大学学报(2021年4期)2021-08-21
发明与创新(2021年19期)2021-05-07
广东蚕业(2021年1期)2021-03-18
江苏农业科学(2019年23期)2019-03-03
小学生导刊(2018年31期)2018-12-06
课外生活(小学1-3年级)(2017年5期)2017-06-10
医学信息(2016年29期)2016-11-28
故事作文·高年级(2016年9期)2016-10-25
