
田间全日照对蔓越橘光合特性的影响*
2021-05-23 09:01汤海昆姜闯道王振兴李亚东
中国果树 2021年4期
汤海昆,姜闯道,王振兴,陈 丽,李亚东
(1吉林省小浆果遗传育种与创新利用工程研究中心,吉林农业大学园艺学院,长春130118)(2中国科学院植物研究所,中国科学院北方资源植物重点实验室)
光不仅能够推动光合作用,而且是影响植物生长发育重要的因素之一。众多研究认为,植物按照需光特性的不同可以分为阳生植物、阴生植物和中间型植物,阳生植物的需光范围为1 000~1 500 μmol·m-2·s-1,而阴生植物达到光饱和仅需要200~500 μmol·m-2·s-1的光强。然而,我国大部分地区晴天条件下的午间最大光强为1 500~2 000 μmol·m-2·s-1[1-2]。虽然光照可以为植物提供能量来源,但过强的光照亦会对植物造成伤害;对于高等植物而言,光合电子传递链包括光系统Ⅱ(PSⅡ)和光系统Ⅰ(PSⅠ),二者对不同环境胁迫的敏感性不同[3]。多数研究认为,PSⅡ对环境胁迫很敏感,强光等胁迫会导致PSⅡ的结构和功能发生变化,尤其是夏季强光条件下,多数植物吸收的光能将会超过自身光合作用所能利用的能量,若不能及时耗散过剩的激发能,将会产生活性氧破坏叶绿素和D1蛋白等,导致PSⅡ发生伤害[4-7]。相反,一般认为,强光条件下植物PSⅠ相对稳定。然而,刘超等对牡丹的研究表明,在强光(1 400 μmol·m-2·s-1)处理下,随着处理时间的延长,牡丹叶片PSⅠ的活性也在逐步下降[7]。秦立琴等在花生上的研究也表明,花生叶片在室温和光强为1 200 μmol·m-2·s-1的条件下PSⅠ活性也显著降低[8]。此外,植物在波动光条件下会使植物的PSⅠ受到伤害。有学者认为其原因是波动光使线性电子传递受阻,电子在PSⅠ传递给氧分子形成活性氧,过量的活性氧生成导致PSⅠ活性受到伤害[9-11]。因此,强光对光合机构的影响还存在争议。另外,大多数强光对植物光合活性影响的研究多在实验室内使用人工光源开展,很少使用田间自然光照。生长光照和处理光照的巨大差异,以及研究条件的不同往往导致结论的差异,但这并不能充分反应田间情况。前人研究表明,田间条件下除非逆境交互胁迫,否则很难发生严重的光抑制现象[12-13]。
蔓越橘属于杜鹃花科(Ericales)越橘属(Vaccinium)红莓苔子亚属(Oxycoccos)植物,俗称蔓越莓、酸果莓及鹤莓等。因其果实有独特的风味和一定的医疗价值,所以在市场上备受瞩目。在我国,蔓越橘主要分布于东北大兴安岭和长白山脉地区,属于林下小灌木[14-16]。其原生境林下光强较弱,但在蔓越橘大面积推广种植时,其栽培环境将不再局限于林下,而会直接暴露在全日照条件下。我们观察到部分蔓越橘新生叶片在全日照条件下会出现变褐、焦枯和停长的现象。与原生环境相比,推测全日照的光强成为影响其生长发育的重要因素。当蔓越橘由原来的林下遮阴或半遮阴环境转移到大田全日照环境时,其叶片是否能适应这种光照条件的改变?是否会发生光抑制并影响其生长?针对这些问题,相关研究尚属空白。因此,本研究将比较蔓越橘叶片生长状况及光系统活性对林下弱光和露地强光的响应,理解蔓越橘叶片在自然光照下的光合机构运转,并为完善我国蔓越橘优质栽培技术提供试验依据。
1 材料与方法
1.1 试验材料与设计
试验于2018年在中国科学院植物研究所进行,试验材料为吉林农业大学选育的蔓越橘优良品系贝恩2年生扦插苗,选取生长状态相似、无病虫害的强壮植株,于新芽萌动时进行处理。采用盆栽,盆高10 cm、内径12 cm。栽培基质和肥水管理在试验期间均相同。
试验设2个处理,分别为林下弱光栽培处理组(L)和露地全日照栽培处理组(H),每组处理设12个重复。林下弱光栽培处理组在午间的最大光强为400~500 μmol·m-2·s-1,在弱光条件下的成熟叶记为LM,新叶记为LN;露地全日照栽培处理组在午间最大光强为1 500~1 600 μmol·m-2·s-1,在全日照条件下的成熟叶记为HM,新叶记为HN。成熟叶为已越冬的2年生叶片,新叶为当年生已完全展开的叶片。成熟叶选取2年生枝条顶端、无病虫害的叶片;新叶选取当年生新梢顶端、无病虫害并已完全展开的叶片。试验各指标测定及采样均选择此类叶片,各指标测定及采样均在处理8周后进行。
1.2 试验方法
1.2.1 叶面积及比叶重的测定
叶面积的测定首先通过随机取样并做标记,然后使用叶面积扫描仪测量叶面积,每个处理6次重复。比叶重的测定参照赵世伟等的方法[17]。
1.2.2 叶绿素含量的测定
随机取样后进行称量,每次称取0.1 g叶片,将称量后的叶片放入80%丙酮溶液中,置于暗处48 h,期间每隔2 h进行1次振荡,保证色素均匀地分布在丙酮溶液中。利用UV-8000型双光束紫外/可见分光光度计(上海元析仪器有限公司)分别在663、646 nm测定OD值,计算出叶绿素a、叶绿素b以及叶绿素的总含量[18]。每个处理6次重复。
1.2.3 快速叶绿素荧光诱导动力学曲线和PSⅠ对820 nm光吸收动力学曲线的同时测定
选一晴天下午,将处理后的植株放入黑暗条件下充分暗适应,利用M-PEA非调制式荧光仪进行测定,分别测量不同处理下蔓越橘快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线)和820 nm光吸收动力学曲线[19-20],PSⅠ的活性指标以820 nm光吸收的最大值与最小值的差值(△I/Io)表示[21]。O-J-IP曲线的测定用3 000 μmol·m-2·s-1的脉冲光诱导,而后进行JIP-test分析[22-23]。每个处理6次重复。
1.2.4 叶绿素荧光猝灭分析
在试验材料充分暗适应0.5 h之后,利用FMS-2脉冲调制式荧光仪(Hansatech,英国),具体光强设定参照乔美玉等的方法[24]。根据测定的荧光参数,计算光下PSⅡ天线转化效率(Fv′/Fm′)、非光化学猝灭(NPQ)。

1.3 数据处理
利用SPSS 19.0数据处理系统对试验所得数据进行统计分析,图表中数值为平均值±标准误,不同小写字母表示在5%水平差异显著。统计结果采用Excel 2013进行作图。
2 结果与分析
2.1 光照环境对蔓越橘叶面积和比叶重的影响
如图1-A所示,全日照条件下生长的蔓越橘新叶的叶面积为1.49 cm2,林下弱光条件下生长的蔓越橘新叶的叶面积为2.06 cm2,差异显著,证明全日照条件显著降低了蔓越橘新叶的叶面积。而光照环境对蔓越橘成熟叶的叶面积影响不显著。
如图1-B所示,相较于林下弱光环境,全日照光环境提高了叶片的比叶重,对成熟叶的提高幅度比对新叶的提高幅度大,但差异均不显著。
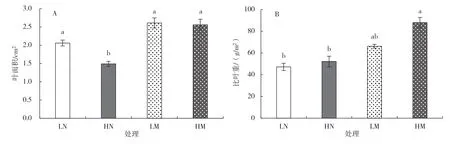
图1 不同光照环境对蔓越橘叶面积和比叶重的影响
2.2 光照环境对叶绿素含量的影响
在光合作用过程中,叶绿素是光能吸收、转化和传递的重要媒介。由表1可以看出,不同光照条件下生长的蔓越橘,新叶和成熟叶的叶绿素a、叶绿素b和总叶绿素变化趋势一致。与林下弱光环境相比,全日照环境显著降低了蔓越橘叶片的叶绿素含量。由此我们推测蔓越橘可以通过调节自身叶绿素含量来适应光环境的改变。
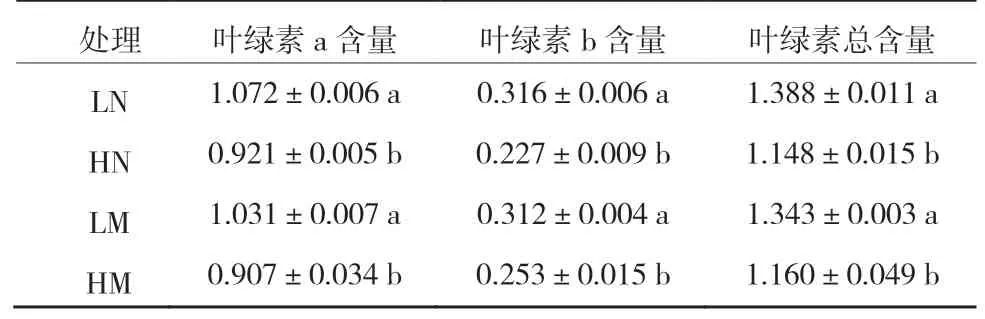
表1 不同光照环境对蔓越橘叶绿素含量的影响 mg/g
2.3 光照环境对叶片荧光诱导动力学曲线的影响
不同光照环境下蔓越橘叶片的快速荧光动力学曲线的变化趋势如图2-A、图2-B所示,新叶和成熟叶在2种光照环境下均表现出全日照条件下具有较低的荧光初始值(Fo)和P点荧光峰值。进一步分析光照环境对蔓越橘快速荧光动力学曲线的变化,我们以P点荧光值标准化诱导曲线。标准化后的曲线表现为全日照条件显著提高了蔓越橘叶片O~I点的相对荧光产量,在J点的时候提高幅度最大,并且新叶的提高幅度比成熟叶的提高幅度大(图2-C、图2-D)。
为了更好地量化不同光照环境对蔓越橘叶片PSⅡ的影响,计算如下参数见表2。由表2可以看出,相比于林下弱光环境,全日照环境下的蔓越橘天线转化效率Fv/Fm显著降低;同样与林下弱光环境相比,全日照环境显著提高了蔓越橘的Vj值,并且对新叶的提高幅度比成熟叶的提高幅度大,其中新叶提高了42.1%,成熟叶提高了25.0%;2种光环境下蔓越橘叶片的Vi值差异并不显著;在分析OJ-I-P曲线过程中,并没有观察到K点的出现(图2),但通过计算Wk,可以看出,相对于林下弱光条件,全日照条件下的成熟叶Wk差异并不显著,但新叶则表现为全日照条件下较高,比林下弱光条件下高出14.8%。

图2 不同光照环境对蔓越橘新叶和成熟叶快速荧光诱导动力学曲线的影响

表2 不同光照环境对蔓越橘叶片Fo、Fm、Fv/Fm、Vj、Vi、Wk的影响
2.4 光照环境对叶绿素荧光猝灭的影响
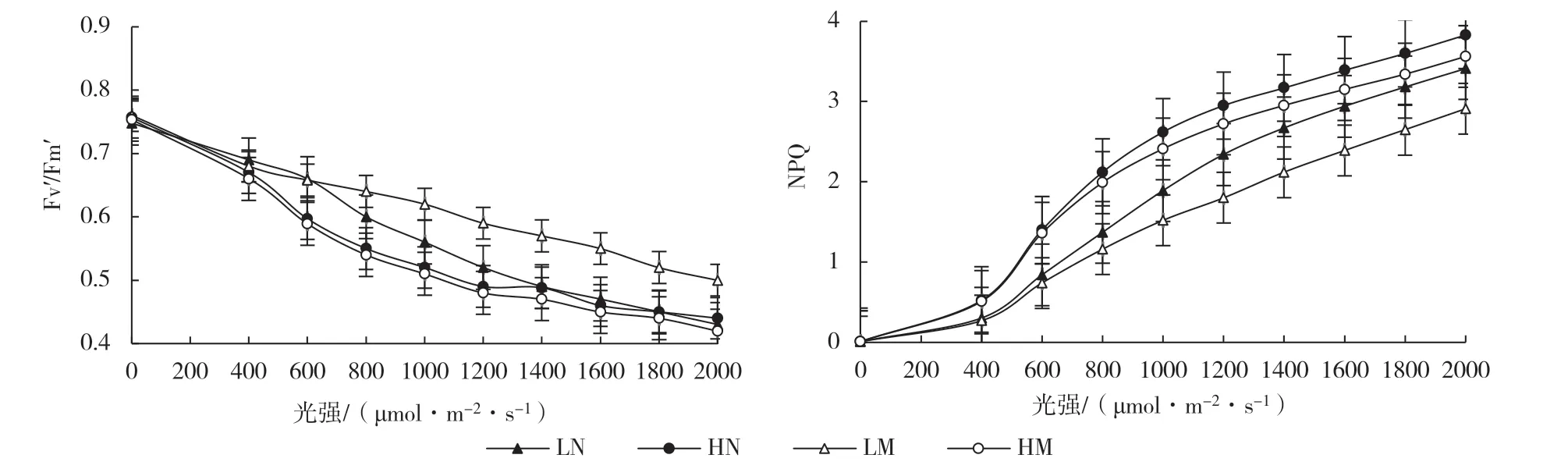
图3 蔓越橘叶片的Fv′/Fm′和NPQ对不同光强的响应
为了更好地分析蔓越橘PSⅡ对光环境的响应,利用FMS测定了林下弱光和全日照条件下蔓越橘叶片PSⅡ的天线转化效率(Fv′/Fm′)和非光化学猝灭(NPQ)对不同光强的响应(图3)。结果表明:林下弱光处理的Fv′/Fm′较全日照处理高,全日照处理新叶的Fv′/Fm′高于成熟叶;在不同光强下,全日照处理叶片的NPQ皆高于林下弱光处理。
2.5 光照环境对叶片820 nm光吸收动力学曲线的影响
图4是光系统Ⅰ的光吸收曲线,可以通过氧化还原衡量其活性[11]。结果表明,光环境对蔓越橘新叶影响较大,主要表现在全日照处理导致PSⅠ的氧化速率和幅度略有下降(图4-A),而对成熟叶PSⅠ的影响不大(图4-B)。这表明光环境能够影响PSⅠ的活性。
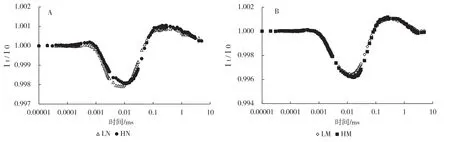
图4 光强对蔓越橘新叶(A)和成熟叶(B)对820 nm光吸收的影响
3 讨论与结论
在本研究中,蔓越橘在林下弱光条件下午间最大光强不超过500 μmol·m-2·s-1,而全日照条件下生长的蔓越橘其午间光强约1 600 μmol·m-2·s-1,二者差异巨大。全日照的强光对蔓越橘新叶的叶面积影响较大,而对成熟叶的叶面积影响不显著。这是由于蔓越橘属于常绿小灌木,成熟叶在上一年入冬前叶面积等形态发育已经基本完成,因此,强光对其叶面积的影响并不显著。而当年生新叶在还未展叶时就开始处理,此时2个处理光环境差异较大,蔓越橘新叶为适应全日照环境,通过减小叶面积来减少光能的捕获。除此之外,全日照条件下的蔓越橘叶片也具有较低的叶绿素含量。不过,无论新生叶片还是成熟叶片,其比叶重均变化较小。所以,从形态角度看,蔓越橘能够通过调节叶片的面积和叶绿素含量来适应光环境的改变。
为优化光合作用适应复杂的光环境,调节天线捕获效率是植物的重要光破坏防御机制之一[25]。通过对荧光猝灭分析可以看到,随着光强的增加,Fv′/Fm′下降,而NPQ逐渐增加;在2种光环境下成长的蔓越橘,新叶的天线下调和热耗散比成熟叶的幅度更大。因此,我们推测全日照强光使蔓越橘叶片产生过剩激发能,而且新叶和成熟叶同时受到强光照胁迫时,新叶的天线调节和热耗散要高于成熟叶。这可能是蔓越橘适应光环境的重要策略。
虽然蔓越橘能够一定程度上适应全日照环境,但是,本研究中2种光环境对蔓越橘叶片的PSⅡ天线转化效率影响较为明显,与林下弱光条件相比,全日照条件下蔓越橘叶片的Fv/Fm降低,但新叶和成熟叶在下降程度上略有不同。这些数据证明无论新叶还是成熟叶其PSⅡ的光化学活性均略有下降。通过JIP-test分析发现,相对于林下弱光条件,全日照条件增加了新叶的Vj和Wk值,增加幅度分别为42.1%和14.8%。而Vj表示QA向QB电子传递的情况[23],可以推测全日照条件导致蔓越橘叶片PSⅡ反应中心复合物的QA-大量积累。因此,相对于林下弱光条件,全日照条件抑制了蔓越橘叶片PSⅡ中QA向QB的电子传递效率,其中新叶电子传递速率受抑制程度更大。前人研究证明,K点的荧光产量(W相)迅速增加的原因是PSⅡ供体侧放氧复合体(OEC)受到伤害[17,26-27]。我们推测全日照强光对蔓越橘新叶的PSⅡ供体侧造成了一定的伤害。而新叶PSⅡ供体侧受到伤害、成熟叶则没有受到伤害的原因可能是幼叶的OEC未完全或刚刚发育成熟,所以更容易受到强光伤害。这与Jiang等[19]的研究结果一致。除PSⅡ外,全日照强光也轻微降低了蔓越橘新叶PSⅠ的活性,但对成熟叶无显著影响。因此,强光会在一定程度上影响蔓越橘新叶2个光系统的活性,从而影响蔓越橘的生长发育。适度遮阴可以作为蔓越橘栽培过程中的常规手段,既可以保证其光合活性,又不会伤害其新叶的生长。
猜你喜欢
中国林副特产(2021年1期)2021-02-25
现代园艺(2017年23期)2018-01-18
创新作文(小学版)(2017年29期)2017-04-04
农业与技术(2016年23期)2016-08-15
动物营养学报(2015年10期)2015-12-01
苏州科技大学学报(工程技术版)(2015年3期)2015-02-28
音乐生活(2014年3期)2014-04-25
植物营养与肥料学报(2013年1期)2013-10-26
食品科学(2013年19期)2013-03-11
食品科学(2010年23期)2010-03-23
