
广东始兴南山木本植物群落特征研究
2021-05-20 12:13肖家亮吴林芳李荣生黄萧洒覃俏梅
亚热带植物科学 2021年1期
肖家亮,吴林芳,李荣生,黄萧洒,覃俏梅
(1.广东始兴南山省级自然保护区管理处,广东 始兴 512521;2.广州林芳生态科技有限公司,广东 广州 510520;3.广东生态工程职业学院,广东 广州 510520)
群落是生态系统的重要组成部分,植物群落是植物之间、植物与环境之间经过长期相互作用形成的较为稳定的结构[1]。植物群落的特征能够综合反映生态系统的基本组成、空间结构、功能特性,对于维持生态系统稳定性具有重要的作用[2]。植物群落中木本植物的组成、生物多样性等是构成群落的基本要素,不同物种所具备的生态功能的差异决定其所在生态系统功能特性的差别[3]。生物量作为森林生态系统研究的重要指标,也与植物群落特征密切相关。生物量可以间接表征植物群落结构的特征,反映了群落总体的物质积累,可用于评价森林的生产力以及固碳潜力[4—5]。因此,探究植物群落的物种组成、生物多样性、生物量等基本特征对于了解群落结构具有积极意义。
已有学者对不同的植被类型、生态系统的植物群落特征进行了深入研究。研究发现,在海南岛的热带季雨林植被类型当中,海南榄仁(Terminalia hainanensis)季雨林较枫香(Liquidambar formosana)季雨林在物种组成、多样性、季相变化等方面具有更强的代表性,是该地区最典型的季雨林群落[6]。此外,在西北干旱区的相关研究也取得了许多成果。雷隆举等[7]研究发现,祁连山东部的高山灌丛群落与草地群落,由于不同植物群落受气候、环境以及物种分布的共同影响,从而形成差异显著的植物群落特征及生物量分配结构。此外,关于群落特征与生态因子(海拔梯度等)间的关系研究也较多,例如林瑜彤等[8]对广东莲花山木本植物群落研究发现,海拔对种类组成、种类多样性、生物量和径级结构有明显影响,物种多样性以及径级结构也对生物量有显著影响。广东始兴南山自然保护区作为始兴地区的生态走廊,对其周边生态环境起着非常重要的调节作用[9]。该生态系统的健康发展有利于维护区域生态环境的稳定性,提升区域生态系统生产力。基于植物群落结构的重要性,相关研究在其他植被类型以及区域均有开展,但关于广东始兴南山的木本植物群落特征的研究未见报道。
广东始兴南山保护区内植物种类丰富,大部分地段植被保持良好,处在植物群落演替的顶级或近顶级,属于典型的亚热带森林生态系统,地带性植被为亚热带常绿阔叶林。本研究基于在南山保护区内设置的3个位于不同海拔区域的1 hm2典型植物群落监测样地,通过调查样地内物种组成、树木生长特征等,分析该区域典型植物群落物种组成、分布、空间结构、多样性、径级结构以及生物量等群落特征,探究海拔对群落特征的影响。研究结果可为该区域实施高效的森林经营管理、监测群落动态变化以及预测群落生产力提供科学依据。
1 材料与方法
1.1 研究区概况
研究区域位于广东始兴南山省级自然保护区,该保护区位于粤北地区始兴县西南面,地理坐标为113°53′58″~114°01′29″E,24°49′37″~24°56′25″N。属南亚热带向中亚热带过渡的地带,全年热量充足,冷暖交替明显,区域内年均温度 19.6℃,年降雨量1537.8 mm[10]。保护区内地形变化显著,以中山地貌为主,平均海拔约500 m,其中黄坑顶海拔1094 m,为保护区最高峰。研究区为典型的亚热带森林生态系统,野生植物资源丰富,包括苏铁(Cycas revoluta)、银杏(Ginkgo biloba)、半枫荷(Semiliquidambar cathayensis)等多种国家保护植物[11]。
1.2 样地设置
2019年12月,在研究区内代表性群落类型中设置3个样地(表1),样地主要在海拔200~800 m之间选取,选择较为平坦地区作为样地布设区域,经现场踏勘核查在原始森林或成熟的次生林中选取结构完整、生物多样性高和人为干扰较少的成片森林进行样地布设。样地大小均为1 hm2,为长100 m、宽 100 m的矩形样地。用全站仪精确测定样地地形、确定样地范围并进行网格化设置,以 20 m×20 m 为基本样方单元,每个样地中共有25个样方,每个样方再细分为16个5 m×5 m 小样方。然后,对样地内所有胸径≥1 cm 的木本植物(含分枝个体)进行每木检尺,测量其胸径(DBH)、树高(H)和冠幅,记录种类、株数、分枝情况和生长状态。样地建设参照《林业系统自然保护区生态因子和生物多样性监测手册》及CTFS/CForBio样地建设规范要求[12]。

表1 样地基本信息Table 1 Basic information of quadrats
1.3 数据分析
依据各样地的重要值确定其优势树种,重要值(IV)=(相对密度+相对频度+相对优势度)/3。采用Shannon-Wiener多样性指数(H)和Simpson多样性指数(D)分析各样地的生物多样性[13],其公式如下:
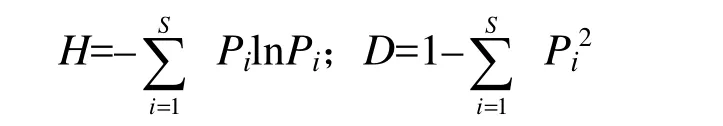
其中,Pi= ni/N,是指第i个物种的个体在样地物种中所占比例,N表示所有种的个体总数,S为样地的物种数量,ni为第i种物种的个体数量[14]。
生物量计算公式为根生物量(W根)、树干生物量(W干)、枝条生物量(W枝)、叶生物量(W叶)之和除以样地面积。各径级的个体百分数为样地中各径级的个体数量占该样地个体总数的百分比。样地树木根据胸径(DBH)大小分为 5个径级,Ⅰ:1.00 cm≤DBH≤10.00 cm;Ⅱ:10.00 cm<DBH≤20.00 cm;Ⅲ:20.00 cm<DBH≤30.00 cm;Ⅳ:30.00 cm<DBH≤40.00 cm;V:40.00 cm<DBH。
采用EXCEL 2013软件对数据进行处理和分析。
2 结果与分析
2.1 南山自然保护区木本植物的物种组成特征
调查表明,3个样地共分布有木本植物60科115属193种。其中低海拔样地NS-A的物种数量最少,为94种,隶属于35科65属,科、属数量也是3个样地中最少的。处于中海拔的 NS-B样地物种数量最多,有127种,隶属于44科80属。尽管高海拔区的NS-C样地科数量与NS-B一样,但其属、种数均较后者少(表2)。
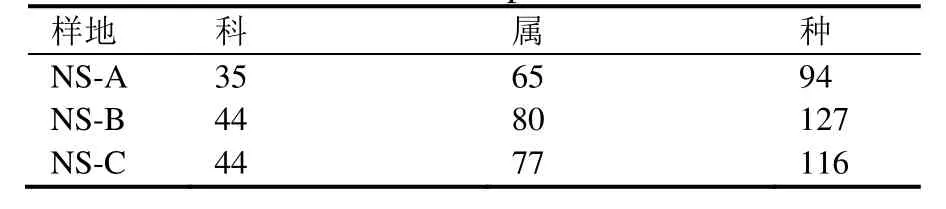
表2 不同样地的科、属、种数Table 2 Number of families, genera and species in different plots
不同样地的优势树种种类不同(表 3)。其中,NS-A样地的罗浮栲(Castanopsis fabri)在样地中占据主要优势,尽管株数不是样地最多的,但其重要值达14.91%。除此之外,栲(Castanopsis fargesii)、华南木姜子(Litsea greenmaniana)、罗浮柿(Diospyros morrisiana)、马尾松(Pinus massoniana)是主要的伴生优势树种。NS-B样地中杉木(Cunninghamia lanceolata)占据绝对优势,重要值为 36.58%,且株数也是样地中最多的(1519株)。而茜树(Aidia cochinchinensis)、黄果厚壳桂(Cryptocarya concinna)、观光木(Michelia odora)、栲的重要值远低于杉木。甜槠(Castanopsis eyrei)是NS-C样地的主要优势树种,重要值为23.83%,株数为1691。尽管凤凰润楠(Machilus phoenicis)的株数与甜槠接近,但其重要值远低于后者,为7.88%。3个样地优势树种中均出现栲。

表3 各样地中重要值排名前5的物种Table 3 The species with the top five importance values in the quadrats
2.2 南山自然保护区木本植物生物多样性指数特征
结果显示,不同样地的Shannon-Wiener多样性指数(H)和 Simpson多样性指数(D)有差异(图 1)。NS-A样地的H值最高,为3.306,随着海拔高度的升高,木本植物的多样性指数降低,H值下降趋势明显,NS-C的H值为3.016。尽管D值随海拔变化的幅度较小,其值也随着海拔的升高由 0.937降至0.898。

图1 不同样地的多样性指数Fig. 1 Diversity index of sample plots
2.3 南山自然保护区木本植物的径级结构特征
研究发现,3个样地的Ⅰ级个体数量分别为3835株、3119株、7546株,分别占据各自样地总数的比例均高于60%,说明样地的树木以小径级的树木个体为主(图2)。位于中等海拔区域的NS-B样地,除了Ⅰ级个体占据主要组成部分之外,Ⅱ、Ⅲ级的个体数量也较多,分别为1184株、257株,占据该样地总数的 25.80%、5.60%,比例值显著高于NS-A、NS-C样地相应径级的个体所占比例。

图2 不同样地木本植物的径级组成Fig. 2 Diameter class distribution of wood plants in sample plots
2.4 南山自然保护区木本植物的生物量特征
调查显示,随着海拔升高,各样地木本植物生物量呈现升高的趋势(图3)。位于高海拔的NS-C样地生物量最大,其中,根 36.16 t·hm-2,干 139.08 t·hm-2,枝 40.90 t·hm-2,叶 8.92 t·hm-2。NS-A 的木本植物生物量为根 21.89 t·hm-2,干 83.66 t·hm-2,枝23.27 t·hm-2,叶 5.49 t·hm-2。NS-B 的木本植物生物量为根31.76 t·hm-2,干 120.99 t·hm-2,枝32.69 t·hm-2,叶8.03 t·hm-2。各部分生物量中,树干均占据最大的生物量,且树干生物量在不同样地间的变化幅度最大。

图3 不同样地木本植物的生物量Fig. 3 Biomass of woody plants in sample plots
3 讨论与结论
不同种类植物的分布依赖于温度、水分、光照等环境因子[15],各自有着不同的生态位[16],因此,伴随着海拔的变化,木本植物的分布也会改变。研究表明,南山保护区的木本植物资源较丰富,且木本植物科、属、种的数量在中海拔区域最高,整体呈现随着海拔升高先增后减的趋势。这与其他地区并不完全一致[8]。这可能是低海拔区域靠近人类生活区,受人类活动的影响相对较大,木本植物种类相对较少。而随着海拔升高,人为干扰逐渐减少,温度、降水等条件更加适宜植物生长,因此物种数量显著上升,研究结果说明中等海拔区域的环境包含更多物种的生态幅。而当海拔继续升高时,温度降低,降水量也随之改变,环境条件的变化会超出部分植物种类的生态幅,从而限制其分布[17]。此外,本研究通过重要值来确定群落中的优势物种,从而了解群落的植被分布特征。重要值是表示某个物种在群落中的地位和作用的综合数量指标[18]。随着海拔升高,不同样地的优势树种变化明显。可能是由于水热条件、土壤理化性质的变化间接改变群落的物种组成、重要值等特征[19]。各样地优势种中,隶属于壳斗科的有6种,隶属于樟科的有4种,说明研究区的优势科为壳斗科、樟科。该结果与前人关于南山保护区优势科的研究结果一致[10]。
物种多样性作为群落特征的一个重要部分,是反映物种丰富度和均匀性的综合指标,也可以用来表示群落结构的稳定程度[20]。但物种多样性同样受到水热条件、土壤性质等非生物条件的影响,因此,海拔变化会对多样性指数产生影响。研究表明,南山保护区的2种多样性指数均随海拔升高呈现下降趋势。这与其他地区的研究结果不一致,有研究发现,植物多样性指数随着海拔的变化呈现单峰的变化格局[21—23]。原因可能是本研究的样地数量不足,尤其是在海拔 300~700 m之间缺乏更多的样地安排,因此,可能忽视了生物多样性指数达到峰值的海拔区域。
径级结构能综合反映植物生长与环境间的关系,是植物群落最基本的结构之一,可用于评价植物群落的生长发育状况、稳定性,预测群落结构的发展。研究表明,南山保护区的高、低海拔区域主要以小径级树木为主,说明林分处于幼龄林阶段。而在中等海拔区域,径级结构相对较为均匀,主要以Ⅰ、Ⅱ、Ⅲ 径级的树木个体为主。说明中等海拔地区的水热条件更适合植物生长,故该样地的径级结构分布更加均匀,林分生长状况整体良好。
生物量可以反映木本植物群落的生产力。木本植物生物量受气候和地形等因子的影响[24]。研究发现,植物的根、干、枝、叶生物量均随着海拔升高而增加。该结果与其他区域关于生物量随海拔变化的研究一致[25]。一方面,是由于高海拔样地木本植物的密度较高,植物个体数量远远高于低海拔和中海拔样地,导致生物量总体较高;另一方面,随着海拔升高,温度、降水等气候因子发生改变,适宜的温度、土壤水分条件有利于植物开展光合作用,从而有效提高了群落的净初级生产力[26]。根据前人研究结果,如果海拔继续升高,随着温度降低,植被类型由乔木型逐渐转为灌木型,生物量可能会随之降低。
综上所述,南山保护区的木本植物共有 60科115属,共计193种,中海拔区域的物种数量最高。随着海拔的变化,林分的优势树种也发生改变,研究区的优势科是壳斗科、樟科。中海拔区域的径级结构分布最为合理,高、低海拔的林分以小径级树木为主。研究区的木本植物生物量随着海拔的升高而增加。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
河北环境工程学院学报(2021年1期)2021-03-19
新教育(2018年8期)2018-08-29
现代园艺(2018年1期)2018-03-15
散文诗(2017年15期)2018-01-19
红岩(2017年6期)2017-11-28
科学种养(2017年6期)2017-06-13
山东青年(2016年1期)2016-02-28
