
树干液流及其主要影响因子对摘芽强度的响应
2021-05-12 06:15李广德张亚雄李豆豆刘金强贾黎明席本野
农业工程学报 2021年5期
李广德,张亚雄,邓 坦,李豆豆,刘金强,贾黎明,席本野
树干液流及其主要影响因子对摘芽强度的响应
李广德1,张亚雄2,3,邓 坦4,李豆豆3,刘金强3,贾黎明3,席本野3※
(1. 国家开放大学农林医药教学部,北京 100039;2.甘肃林业职业技术学院,天水 741020; 3. 北京林业大学省部共建森林培育与保护教育部重点实验室,北京 100083;4. 河南省林业调查规划院,郑州 450045)
为了探讨摘芽强度对树木液流变化特征及其主要环境调控因子的影响,为人工林及果园等高效抚育管理提供参考,该研究以毛白杨()为研究对象,设置4种摘芽处理(不摘芽CK、轻度摘芽BP1.0、中度摘芽BP1.5、重度摘芽BP2.0),采用热扩散式液流检测技术,结合自动气象站,对造林后第2~3个生长季不同摘芽处理后的树干液流、环境因子等进行连续观测。结果表明:1)摘芽对树干液流速率日变化趋势无影响,晴天都呈“单峰型”;2)摘芽对树干液流特征值影响显著,试验期间CK、BP1.0、BP1.5、BP2.0处理的液流速率平均值分别为1.405×10-3、1.503×10-3、1.462×10-3、1.033×10-3cm/s,BP2.0显著小于其他处理,CK、BP1.0、BP1.5之间无显著差异(>0.05);3)影响不同时段、不同处理树干液流的主要环境因子不同,瞬时值日进程的影响因素为太阳辐射和饱和水汽压亏缺,白天及全天日均值的主要影响因子为太阳辐射和日空气温度最大值,夜间液流日均值的影响因子主要为日空气温度最小值和日平均相对湿度;4)不同摘芽强度下毛白杨幼林夜间液流占全天液流的5%~70%,夜间液流与白天及全天液流间呈显著负相关。因此,从促进植物水分生理活动的角度看,轻度和中度摘芽更合适。
蒸腾;环境;温度;耗水;液流;摘芽;毛白杨
0 引 言
树干液流的99.8%用于叶片蒸腾,可直接反映树木的耗水特性,是研究树体内水分运动的常用监测指标。热扩散法是进行树干液流测定的常用和比较准确的方法,得到了非常广泛的应用[1-4]。林木树干液流不仅受自身遗传特性的制约,也受土壤水分和气象因素的影响,但其主导因子随时空的变化而不断变化。通过研究树干液流的变化规律及其与环境因子的关系,能够更好地理解和揭示树木水分利用状况及其对环境条件变化的响应[2-5]。
摘芽是在芽膨大时将其摘掉,促进顶芽生长的一种措施。对于用材树种,摘芽能提高树干圆满度,培育无节高干良材,促进林木生长[6];对于枣树[7]、葡萄[8]、油桃[9]、脐橙[10]、樱桃[11]等经济树种,摘芽能尽早形成理想冠型、提前开花结实、提高坐果率,达到优质丰产。现有研究主要集中在摘芽对冠型、树木生长、目标产物产量和质量、早熟性影响等方面,更多的是将其作为一种栽培技术进行描述,未见摘芽对树木生理生态影响以及摘芽强度选择等方面的研究报道。
毛白杨在苗木培育时,其侧芽常全部萌发长成侧枝,影响苗木质量,降低造林成活率并影响幼树生长[12],因此,非常有必要在毛白杨苗木培育或新造林后对其侧芽进行摘除。关于毛白杨的树干液流,已有学者对其研究方法的准确性、不同方位液流差异、液流对地下水位和气象因子的响应等方面进行了研究[13-15]。但不同时段毛白杨树干液流的相关性及影响因子、摘芽对树干液流的影响等还未见报道。基于此,本研究以造林后不同摘芽强度的毛白杨为研究对象,利用热扩散式探针和自动气象站,研究树干液流变化特征及其与主要气象因子的相关性,主要目的是:1)了解摘芽强度对树干液流特征的影响;2)明确毛白杨夜间液流与白天、全天液流的相关性;3)探究不同时段和不同摘芽强度下毛白杨树干液流的主要影响因子。
1 材料与方法
1.1 研究地概况
试验地位于山东省高唐县国有旧城林场(36°48′47″N,116°05′25″E),海拔30 m。属暖温带半湿润季风区域大陆性气候,光照充足,热量丰富。年均降水量544.7 mm,降水主要集中在7-8月,年均蒸发量1 880 mm,年均温12.0~14.1 ℃,极端最高气温41.2 ℃,极端最低气温−20.8℃。年日照总时数2 651.9 h,无霜期204 d,地下水位深6 m左右。试验地土壤为砂壤土,土壤容重平均1.41 g/cm3,pH值平均8.42,其他基本理化性质参见文献[16]。试验期间(2017年8月-11月,2018年4月-6月)平均气温18.9 ℃,降水量合计229.1 mm。
1.2 试验材料
研究对象为毛白杨杂种优良无性系S86。试验林于2016年3月14日-3月20日营造,两年根两年干苗,摘芽试验林分0.47 hm2,密度为1 666株/hm2(株行距为2 m×3 m),行向南北,无间作。基肥为缓释肥(N:P:K=14:12:5,有机质15%以上),施肥量为150 g/穴。栽植后定干高度为3 m。
1.3 试验设计
造林后分别摘除树干1/3树高(1.0 m,BP1.0,轻度)、1/2树高(1.5 m,BP1.5,中度)、2/3树高(2.0 m,BP2.0,重度)以下所有的芽,另设对照处理(不摘芽,CK)。试验采用完全随机区组布设。5个区组,每个区组4个小区(4种处理),每小区样树12株,区组间设2行保护行,小区间设1行保护行(图1)。
1.4 测定指标及方法
1.4.1 树干液流
选取第Ⅱ区组中间部位样树,于2017年8月15日安装热扩散液流探针(TDP-30, Dynamax Inc., Houston, USA),每种处理各2株,探针安装于距地面约50 cm处以保证所有处理的探针都安装于第一分枝之下。液流速率每10 s测定1次,每10 min计算1次平均值并记录于TDC4S便携式植物茎流采集仪(北京时域通科技有限公司)。样树主要参数见表1。
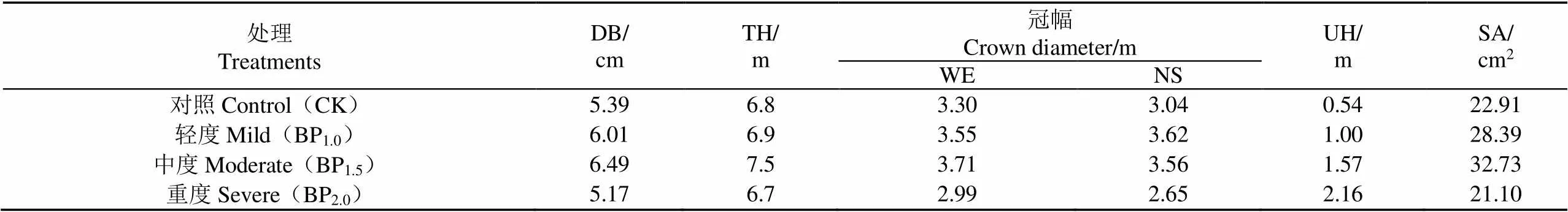
表1 不同摘芽强度处理测定样树主要参数
注:DB,胸径;TH,树高;UH,活枝下高;SA,胸径处边材面积;WE,东西向;NS,南北向。
Note: DB, diameter at breast height; TH, tree height; UH, under-branch height; SA, sapwood area at breast height; WE, from East to West; NS, from South to North.
数据采集下载后,利用DYNAMAX 公司提供的分析软件对原始数据处理与计算。液流速率计算公式如下[1]:
式中V为树干液流速率,m/s;dTM为24 h内上下探针的最大温差值,℃;d为某时刻两探针的温差,℃;为无量纲参数。
1.4.2 气象因子
气象因子利用距试验地约0.9 km空旷地上的全自动气象站(Delta-T Devices Ltd.,Cambridge,England)测定,指标有太阳辐射(Q,w/m2)、空气温度(T,℃)、空气相对湿度(RH,%)、风速(WS,m/s)、降水量(,mm)等,与液流测定同步。饱和水汽压亏缺(VPD,kPa)通过下式计算[17]:
式中、、为常数,分别为0.611 21 kPa,17.502,240.97 ℃。
参考作物潜在蒸散量(ET0,mm)用联合国粮农组织推荐的Penman-Monteith计算[18]:
式中R为冠层表明净辐射,MJ/(m2·d);为土壤热通量,MJ/(m2·d);为饱和水汽压曲线在气温为时的斜率,kPa/℃;为日平均气温,℃;为干湿表常数,kPa/℃;2为距地面2 m处的风速,m/s;e为饱和水汽压,kPa;e为实际水汽压,kPa。
1.5 数据分析
数据分析和处理利用Dynamax公司提供的液流数据分析及SPSS20.0统计软件。不同摘芽强度的摘芽数量、液流速率等的差异利用单因素方差分析进行比较,多重比较在0.05水平上利用最小显著差异(least Significant Difference,LSD)法检验。Spearman分析评价液流与环境因子的相关性。通径分析确定影响液流的主要环境因子,直接通径系数在对因变量正态性检验后通过SPSS软件“Analyze-Regression-Linear”中Coefficients获得。
2 结果与分析
2.1 摘芽强度对摘芽数量的影响
造林后第1次摘芽于2016年4月18日进行,萌发的新芽于2016年5月19日进行了第2次摘除,各处理摘除的侧芽数量为2次摘除的芽的总数,通过记录摘去的芽的数量获得,见表2,被摘除的侧芽数量随着摘芽强度的增大而增加,中度和重度摘芽数显著大于轻度(<0.05)。

表2 不同强度处理的摘芽数
注:不同字母表示在0.05水平上差异显著(2-tailed),下同。
Note: Different lowercase letters indicate significant difference among treatments (<0.05), the same below.
2.2 不同摘芽强度毛白杨树干液流速率变化特征
2.2.1 日变化
晴天树干液流速率日变化在7个典型日(春季:2018年4月17日、5月14日,夏季:2018年6月14日、2017年8月21日,秋季:2017年9月13日、10月13日、11月12日)呈一致的“单峰型”曲线(图2)。液流启动时间春季(05:30-06:10,平均05:40)早于夏季(06:50-08:00,平均07:30)早于秋季(07:50-09:40,平均08:40),到达峰值时间春季(11:00-13:30,平均11:50)早于秋季(12:00-14:40,平均12:45)早于夏季(12:50-13:10,平均13:00);高峰时段运行时长春季(约8 h)长于夏季(7~8 h)长于秋季(3~6 h);夜间液流开始时间随着时间推移而前移(春季20:40-22:00,夏季19:30-20:50,秋季16:30-19:50)(表3)。可见,不同摘芽处理虽然影响了毛白杨的胸径、树高、冠幅、枝下高等树冠结构因子(表1),但并未改变其树干液流日变化趋势。说明在同一环境条件下,同一树种蒸腾耗水日变化特征并不会因树冠结构的改变而改变,而主要受环境因子的综合影响和制约。
不同摘芽强度下毛白杨液流速率峰值及日均值随时间变化而变化(表3)。各处理液流速率峰值在2017年8月和9月差异不显著(>0.05),10月开始(11月除外)呈现出显著差异,峰值最小的是BP2.0,其他处理差异不显著。从日平均液流速率来看,除2017年8月21日各处理无显著差异(>0.05)外,其余都有;而有显著差异的日期,除11月21日外,全是BP2.0最小;不同处理日平均液流速率随时间变化格局与峰值类似,最大值由起初的CK逐步过渡为BP1.0和BP1.5。可见,摘芽强度通过影响树冠结构而影响了毛白杨液流速率峰值、日均值等特征值,但这种影响也需要一定的时间积累才能有所表现,因为在试验前期各处理特征值并没有差异。重度摘芽导致树冠最小的BP2.0液流峰值和日均值除深秋外都最小,但树冠大、侧枝多、叶片多的CK并没有一直保持最大的液流速率。
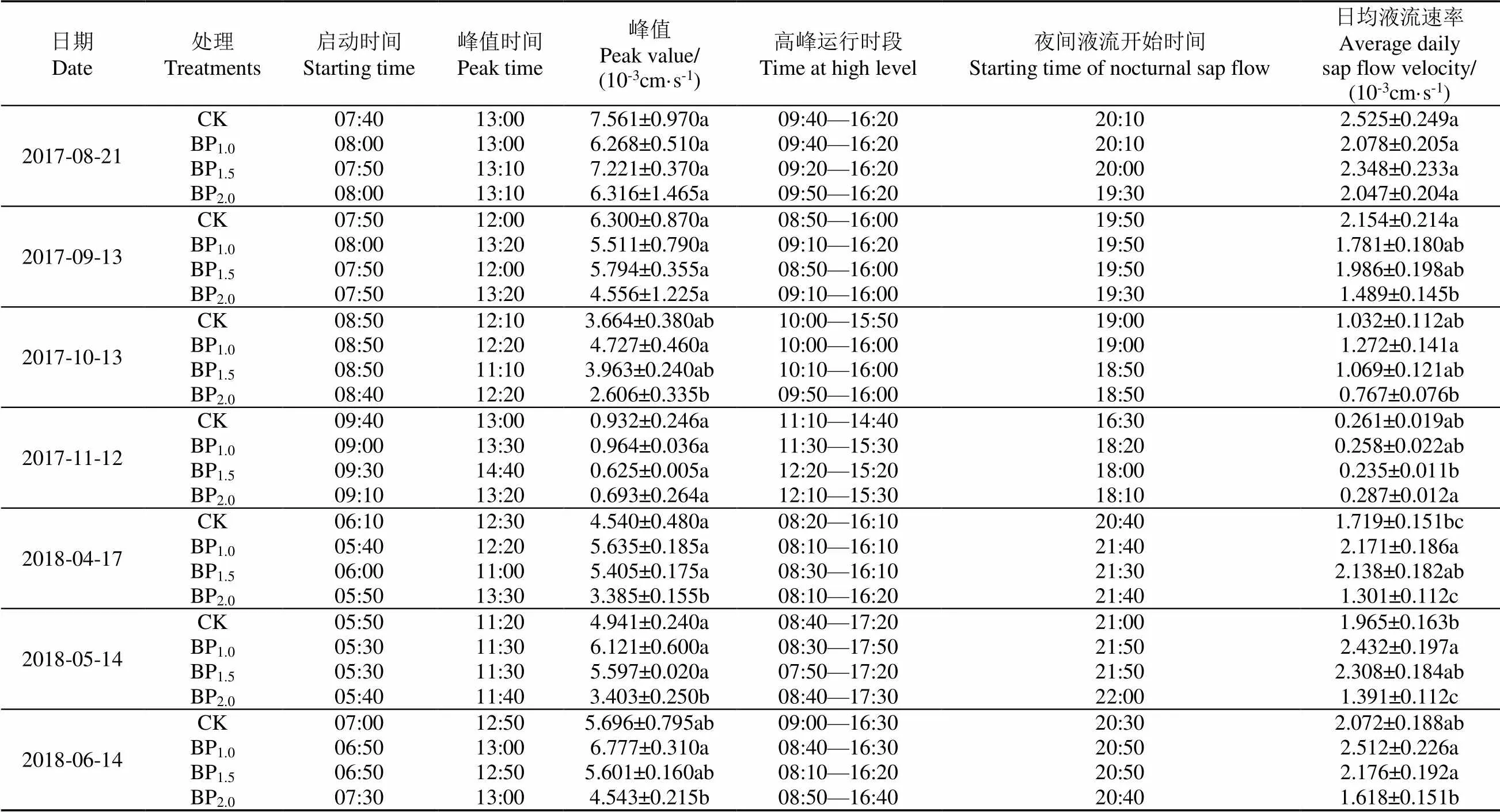
表3 晴天不同摘芽强度毛白杨树干液流日变化特征值
2.2.2 季节变化
“春季-夏季-秋季”各处理液流速日均值呈现“低-高-低”的趋势(表3,图2,图3,图4)。所有测定日CK、BP1.0、BP1.5、BP2.0液流速率平均值分别为1.405×10-3、1.503×10-3、1.462×10-3、1.033×10-3cm/s,差异显著(<0.05),BP2.0显著小于其他处理,CK、BP1.0、BP1.5之间无显著差异。说明水热条件最好的夏季,毛白杨整树蒸腾更旺盛,生理活动更旺盛;从整个生长季尺度看,毛白杨蒸腾耗水量随摘芽强度增大体现为先增大后减小。
从不同月份各处理液流平均值来看,除11月,其余月份都有显著差异(图4)。但不同月份各处理均值大小格局有变化,2017年8月和9月是CK最大,10月开始BP1.0相对最大,到2018年6月时BP1.0显著大于其余处理。11月各处理液流均值虽然差异不明显,但却表现出与其他月份不同的格局,其他月份都处于最低的BP2.0并不是最小的。因此,在季节变化尺度,毛白杨树干液流一方面随环境条件而变,同时也受自身树冠结构特征等多重因素影响,变化复杂,但树冠最小的重度摘芽处理在整个生长季蒸腾耗水量最小且显著小于其他处理。
2.3 环境因子对不同摘芽强度下毛白杨树干液流的影响和调控
2.3.1 对树干液流瞬时值的影响
从不同时段树干液流瞬时值与环境因子的相关系数()及直接通径系数(Direct Path Coefficient,DPC)来看(表4),晴天全天,毛白杨树干液流与Q、TWSVPD等环境因子极显著正相关,与RH极显著负相关;白天时段,与Q、TVPDRH的相关性与全天类似,但与WS负相关;夜间时段,与VPD、WS正相关,与TRH负相关,与Q不相关。通径分析可以发现,影响不同处理不同时段树干液流瞬时值日变化的主要环境因子不同:影响CK、BP1.0、BP1.5、BP2.0液流速率全天变化的环境因子分别为Q和VPD、VPD和Q、Q和VPD、Q和VPD;白天为T和Q、Q和VPD、T和Q、T和Q;夜间为VPD和TVPD和RH、VPD和T、T和RH。说明不同时段主要环境因子对液流速率瞬时值的影响不同,对全天液流的影响大于白天,对夜间液流瞬时值的影响最小;QVPD在全天、白天、夜间3个时段都与液流瞬时值正相关即液流速率随Q和VPD的增大而增大,RH在所有时段都与液流瞬时值负相关。
2.3.2 对树干液流日变化的影响
在以日平均液流速率为计算单位的季节变化尺度上,毛白杨液流日均值在不同时段与主要环境因子的相关性也不同(表5)。全天日均值和白天日均值与环境因子的相关性一致,都与ET0、Q、TVPD等正相关,与、RHWS等负相关;夜间液流日均值与ET0、、Q、TRH等负相关,与WS正相关,而与VPD的相关性在不同强度下表现不同。从液流速率与环境因子相关系数来看(表4和表5),Q、T对日均值的影响与瞬时值类似,但Q对瞬时值影响更大而T对日均值影响更大;RH对日均值影响更小;WS对瞬时值和日均值白天和夜间时段影响相同但对全天的影响不同,对瞬时值的影响更大;VPD对瞬时值的影响更大。直接通径系数值表明,影响毛白杨CK、BP1.0、BP1.5、BP2.0液流速率全天日均值的环境因子分别为Tmax和Q、Q和T、Q和T、Q和Tmax;白天为Tmax和Q、Q和T、Q和Tmax、Tmax和Q;夜间为TminRH和QRH和VPD、Tmin。可见,在更大的时间尺度上,树木液流和蒸腾耗水的影响因素更多,作用更为复杂。
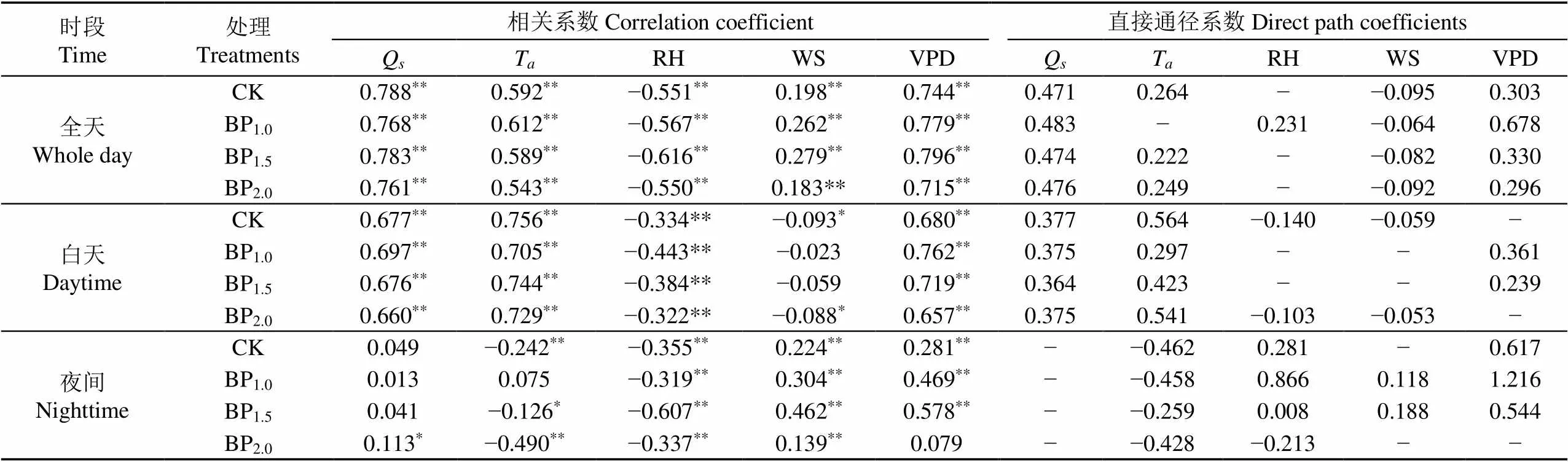
表4 晴天不同摘芽强度树干液流瞬时值与环境因子的相关系数及直接通径系数
注:*表示在0.05水平上显著,**表示在0.01水平上显著;Q、T、RH、WS、VPD分别表示太阳辐射、空气温度、空气相对湿度、风速、水蒸气亏缺;下同。
Note:*indicates significance at 0.05 levels,**indicates significance at 0.01 levels;Q,T, RH, WS and VPD represent solar radiation, air temperature, air relative humidity, wind speed and vapor pressure deficit, respectively; the same below.
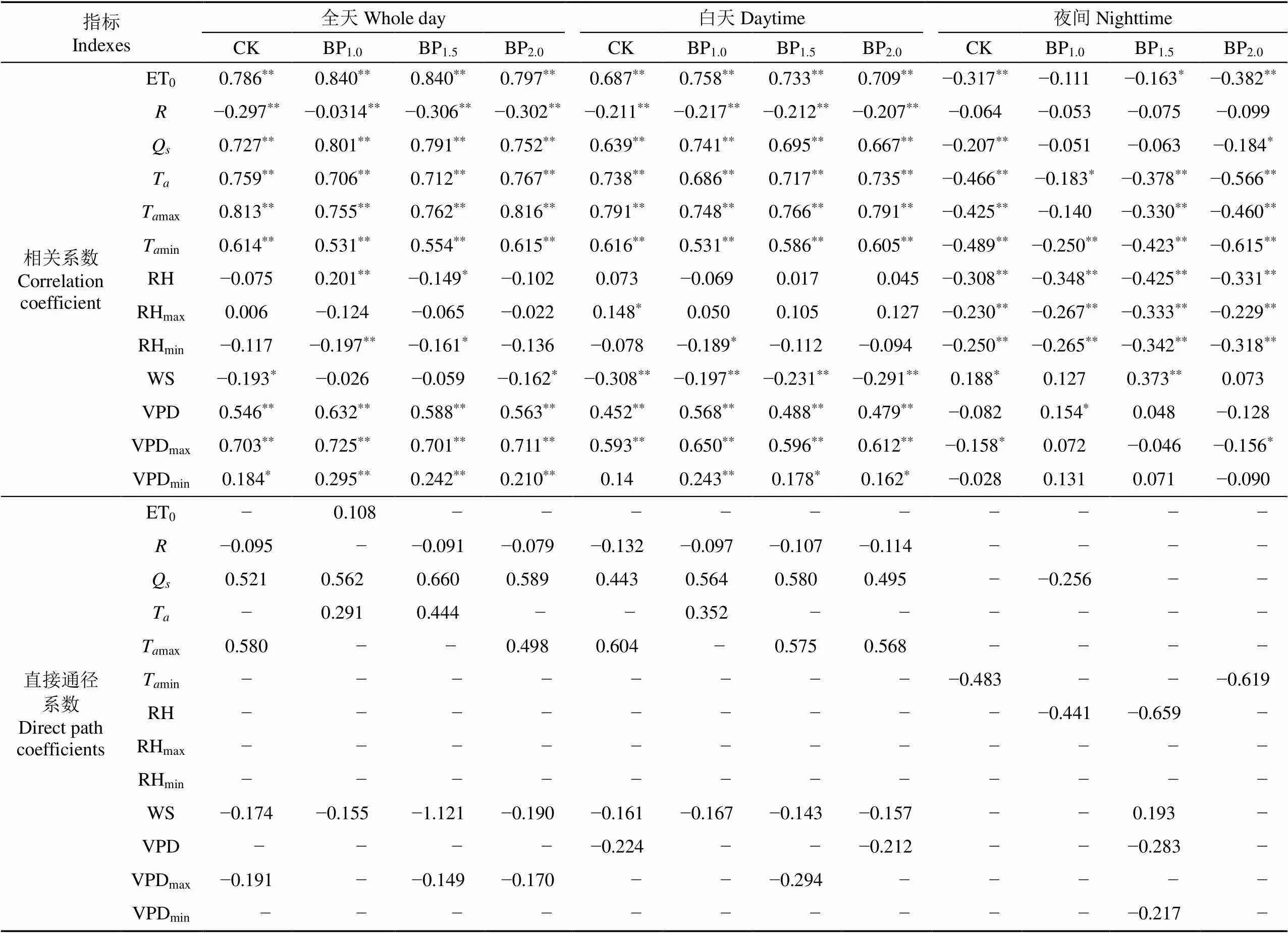
表5 不同摘芽强度毛白杨树干全天、白天、夜间的液流均值与环境因子的相关系数及直接通径系数
注:下标max和min指日最大值与最小值。
Note: max andmin represent daily maximum and minimum values.
2.4 摘芽强度对毛白杨夜间液流的影响
2.4.1 夜间液流变化
毛白杨幼林存在明显的夜间液流(表6)。造林后第2个生长季(2017年),不同处理夜间液流速率无显著差异(>0.05);从第3个生长季(2018年),不同处理夜间液流呈现出显著差异,春季BP1.5夜间液流显著大于BP1.0和BP2.0,夏季夜间液流BP1.0显著高于其他处理(表6)。
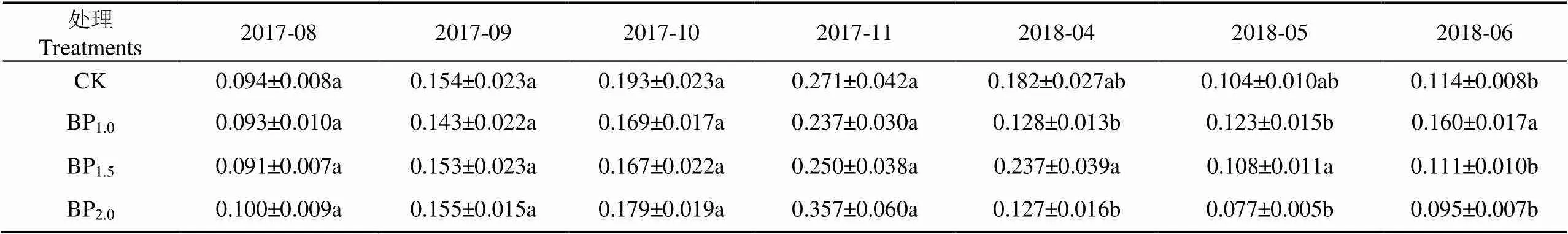
表6 不同摘芽强度毛白杨夜间液流月平均值
从夜间液流与全天液流的比值来看,不同处理夜间液流速率约为全天液流速率的5%~70%(表7),日均液流值相对较小的10、11、4月,其夜间液流占比相对较大,重度处理毛白杨夜间液流速率占比普遍更大。

表7 不同摘芽强度毛白杨夜间液流速率与全天液流速率之比
2.4.2 夜间液流与全天及白天液流的相关性
从不同摘芽强度处理下毛白杨夜间液流日均值与相应的全天及白天液流日均值的相关性来看,毛白杨夜间液流与全天及白天液流都呈负相关,与白天液流均值的相关性大于全天,但液流均值最大的轻度处理的夜间液流与其余液流均值的相关性不显著(表8)。
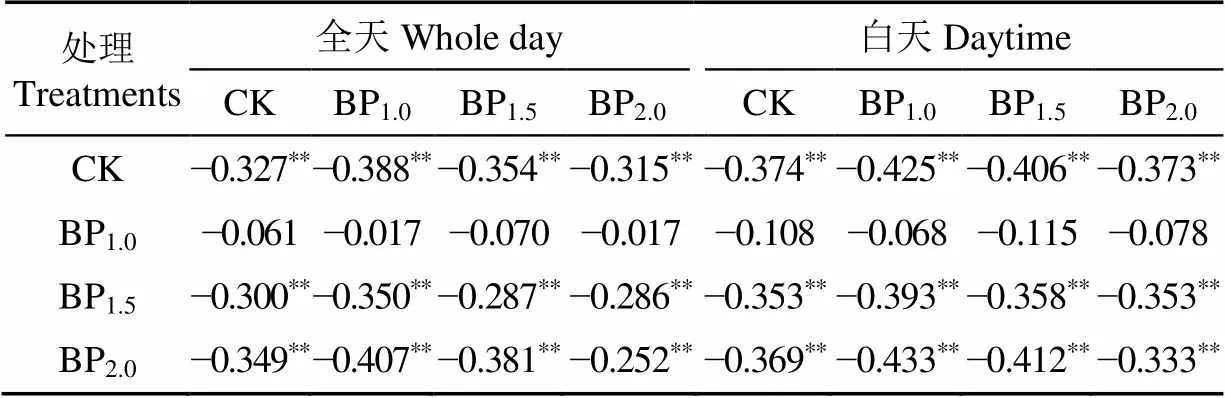
表8 不同摘芽强度毛白杨树干夜间液流均值与日均值、白天液流均值的相关系数
注:样本数179。
Note: Sample size is 179.
3 讨 论
与河北杨()[4]、欧美杨64号(cv. ‘N3016’)[19]、银白杨(×)[20]、北京杨()[21]、小叶杨()[22]、胡杨()[23]等类似,晴天毛白杨无性系S86幼林树干液流速率日变化呈“单峰型”,但其值整体小于辽西农林复合系统中4年生的欧美杨64号[19]、毛乌素沙地40年生的小叶杨[22]、冀北坝上33~36年生的北京杨[21],以及黑河下游荒漠河岸建群种胡杨[23],大于克拉玛依干旱荒漠区9年生的银白杨[20]、陕北黄土区的河北杨[4]。而和6年生三倍体毛白杨无性系B301((×)×)相比[24],其值小于灌溉处理但大于未灌溉。可见,同一树种树干液流日变化趋势相同但其大小则受树龄、气候及立地、抚育管理措施等因素影响。
摘芽对毛白杨瞬时液流速率日变化规律无影响,但在长时间尺度上会引起蒸腾耗水格局的变化。液流速率日平均值在造林后第2个生长季的8月和9月CK最大、BP2.0最小,10月BP1.0最大、BP2.0最小,11月BP2.0相对较大;第3个生长季变化为BP1.0最大、BP2.0最小且差异显著。从月液流均值的角度,除深秋的11月外,不同处理均有显著差异,BP2.0显著小于其他处理。与本研究预想不同的是,CK虽然具有更多的侧枝和叶片,但其液流值却不是一直保持最大,尤其从第3个生长季开始,轻度摘芽后的液流速率一直保持最大,且随着时间推移BP1.0与其他处理的差异越来越明显和显著(图4)。魏新光等[25]的研究表明,枣树蒸腾耗水量并不是随其叶面积指数的增加而增加,而是随着叶面积指数的增加呈现先增加后减小的趋势,与本研究轻度处理后液流最大的研究结果类似,也说明此时的冠层结构最有利于毛白杨的蒸腾。可见,从第3个生长季开始,过多的侧枝和叶片并不能提高毛白杨的整树蒸腾,反而成了影响水分蒸腾的负面因素。因此,从栽培管理角度看,应该从一定时间开始(对于毛白杨,是造林后第3个生长季),剪除树木过多的枝条,以促进其生理活动。
影响植物蒸腾的环境因子众多,树木液流与环境因子的关系也复杂多变,不同时间尺度上环境因子对树木蒸腾的影响程度也不尽相同[26]。本研究表明,影响不同时段、不同强度毛白杨液流速率的主要环境因子不同。影响其瞬时值日进程即日变化的主要因素为Q和VPD;白天影响其瞬时值变化的主要因素为T和Q,夜间影响因素在不同强度摘芽处理下表现复杂,主要为VPD、T和RH。影响毛白杨液流白天及全天日均值的环境因子为Q和Tmax;而夜间液流日均值的影响因子主要为Tmin和RH。可见,对于白天及全天液流值,Q无论在瞬时尺度,还是在比较长的月尺度上,都是影响树干液流的主要因素,与欧美杨64号[19]、银白杨[20]、小叶杨[22]等一致。在分析毛白杨液流较长时间尺度的影响因素时,本研究引入了潜在蒸散量ET0,日最大和最小空气温度、相对湿度、水蒸气压亏缺等因素,发现日最高温、日最低温也是影响树干液流的主要因素,这在已有的研究中是不多见的。可见,植物蒸腾影响和调控机制具有复杂性和相对性,如何构建一套普遍适用的比较系统和科学的蒸腾耗水调控机制需要不断研究。
植物夜间蒸腾和水分补充对维持其水分平衡等具有重要生态水文意义。针对农作物,研究人员已对苜蓿、黄豆、高粱、小麦、番茄、向日葵等的夜间蒸腾做了研究[27]。针对杨树,研究人员已对三倍体毛白杨B301[13]、北京杨[21]、小叶杨[28]、胡杨[29]、欧美杨[30]等作了分析,认为杨树夜间液流占全天液流的2.5%~38%。本研究表明毛白杨杂种无性系S86存在夜间液流,液流速率为全天液流速率的5%~70%。对于不同摘芽处理,从造林后第3个生长季开始呈现出显著差异,与日均液流一致,日均液流值较大的BP1.5和BP1.0夜间液流均值最大(表6)。但液流日均值、月均值均最小的BP2.0夜间液流对全天液流的贡献最大(表3,表7,图3,图4),与液流值较小的雨天小叶杨[28]、不健康北京杨[21]夜间液流贡献类似。
本研究表明毛白杨夜间液流与白天及全天液流负相关(表8),在分析环境因子对液流影响时也发现,全天及白天液流与ET0、Q、T、VPD等的日均值正相关,而夜间液流与这些因素负相关,说明这些指标越大则全天和白天液流越大、夜间液流越小。结合不同摘芽强度对毛白杨夜间液流的影响(表6和表7),可以判断,对于毛白杨,日均树干液流愈小,其夜间液流越活跃,占比愈大。树干夜间上升液流的存在,一方面是根压的作用,一方面是夜间气孔开放和叶片蒸腾,也可能是由于水容存在而补充水分,但无论如何,夜间液流是普遍存在的[31-33]。而为何液流日均值整体偏小的处理其夜间液流更明显,这一规律对于其他树木和农作物是否也同样适用,需进一步研究和探讨。
4 结 论
为研究摘芽强度对树干液流变化特征、夜间液流的影响及其与环境影子的相关性,本文以毛白杨为例开展试验,利用热扩散法连续2 a对不同摘芽强度下毛白杨树干液流及主要环境因子进行监测,初步探明摘芽强度对树干液流日、季节变化规律和夜间液流的影响,明确了毛白杨夜间液流与全天、白天液流的相关性,了解了不同摘芽强度下树干液流对环境因子的响应机制,得出以下结论:
1)摘芽对树干液流速率日变化趋势无影响,晴天都呈“单峰型”曲线,峰值出现在11:00-13:00左右;摘芽对液流峰值、日均值等特征值的影响随时间推移而变化,日平均和月平均的最大值由对照逐渐转变为轻度处理;随摘芽强度的增大液流值先增大后减小,重度摘芽后由于其树冠变小,液流值持续最小;从促进水分生理活动角度看,摘芽强度不宜超过其侧芽总量的1/2。
2)不同时段、不同处理树干液流的主要影响因子不同,日变化尺度下瞬时值的主要影响因子为太阳辐射和饱和水汽压亏缺;整个生长季尺度,白天及全天日均值的主要影响因子为太阳辐射和气温最大值,夜间液流日均值的影响因子主要为日空气温度最小值和日平均相对湿度。
3)毛白杨幼林夜间液流占全天液流的5%~70%,夜间液流与白天及全天液流呈负相关,不同处理夜间液流从造林后第3个生长季开始出现显著差异,日均液流愈小其夜间液流占比愈大。
良好树形、合理树冠的建立是果园丰产优质的基础,整形修剪是树冠养成的关键,在枝条尚未形成前的摘芽作为修枝的一种变形作业,能经济有效控制树形发展,鉴于上述研究结论,对于果园树木,合理强度的摘芽可促进其生理活动,为果园产量和质量的提高奠定基础。
[1]Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements[J]. Tree Physiology, 1987, 3(4): 309-320.
[2]李广德,贾黎明,孔俊杰. 运用热技术检测树干边材液流研究进展[J]. 西北林学院学报,2008,23(3):94-100.
Li Guangde, Jia Liming, Kong Junjie. Advances in thermal techniques on stem sap flow[J]. Journal of Northwest Forestry University, 2008, 23(3): 94-100. (in Chinese with English abstract)
[3]Bovard B D, Curtis P S, Vogel C S, et al. Environmental controls on sap flow in a northern hardwood forest[J]. Tree Physiology, 2005, 25(1): 31-38.
[4]刘崴,魏天兴,朱清科. 水蚀风蚀交错区河北杨树干液流密度特征及其对环境因子的响应[J]. 北京林业大学学报,2018,40(5):73-81.
Liu Wei, Wei Tianxing, Zhu Qingke. Dynamics of sap flow density ofand its responses to environmental variables in the water-wind erosion crisscross region on the Loess Plateau[J]. Journal of Beijing Forestry University, 2018, 40(5): 73-81. (in Chinese with English abstract)
[5]Xu S Q, Yu Z B. Environmental control on transpiration: A case study of a desert ecosystem in northwest China[J]., 2020, 12: 1211. doi:10.3390/w12041211.
[6]沈国舫. 森林培育学[M]. 北京:中国林业出版社,2001,277-278.
[7]唐忠建,何梅,高俊萍. 摘心与抹芽对木质化枣吊形成的影响[J]. 北方园艺,2012(22):26-28.
Tang Zhongjian, He Mei, Gao Junping. Effects of pinching and bud picking on the formation of the hanging wooden of the dates[J]. Northern Horticulture, 2012(22): 26-28. (in Chinese with English abstract)
[8]骆生,朱爱云,韩丽媛. 设施葡萄实用栽培技术[J]. 北方园艺,2010,12:58-59.
[9]林永高,胡章琼,陈雪金. 福州早熟油桃优质丰产栽培技术[J]. 中国南方果树,2016,45(1):133-135.
[10]赵文才. 水库高湿区脐橙园早结丰产优质配套栽培技术[J]. 中国南方果树,2001,30(2):5-6.
[11]Macit I, Lang, G A, Demirsoy H. Bud management affects fruit wood, growth, and precocity of cherry trees[J]. Turkish Journal of Agriculture and Forestry, 2017, 41(1): 42-49.
[12]李文起,池金田. 毛白杨苗木抹芽试验[J]. 山东林业科技,1992(2):38-40.
[13]李广德,贾黎明,富丰珍. 不同方法测定三倍体毛白杨整树蒸腾的比较[J]. 林业科技开发,2014,28(5):41-44.
Li Guangde, Jia Liming, Fu Fengzhen. Comparison on the whole-tree water use of hybrid triploid of Chinese white poplar between the whole tree potometer and thermal dissipation probe[J]. China Forestry Science and Technology, 2014, 28(5): 41-44. (in Chinese with English abstract)
[14]刘洋,王烨,王斐,等. 宽窄行栽植下毛白杨不同方位树干液流的差异[J]. 中南林业科技大学学报,2018,38(10):95-105.
Liu Yang, Wang Ye, Wang Fei, et al. Azimuthal variation in sap flux density ofunder wide and narrow row planting scheme[J]. Journal of Central South University of Forestry & Technology, 2018, 38(10): 95-105. (in Chinese with English abstract)
[15]Di N, Xi B Y, Clothier B, et al. Diurnal and nocturnal transpiration behaviors and their responses to groundwater-table fluctuations and meteorological factors ofin the North China Plain[J]. Forest Ecology and Management, 2019, 448: 445-456.
[16]贺曰林,王烨,张宏锦,等. 地表滴灌水氮耦合对毛白杨幼林生长及土壤水氮分布的影响[J]. 农业工程学报,2018,34(20):90-98.
He Yuelin, Wang Ye, Zhang Hongjin, et al. Coupling effects of water and nitrogen on tree growth and soil water-nitrogen distribution in youngplantations under surface drip irrigation[J]. Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE), 2018, 34(20): 90-98. (in Chinese with English abstract)
[17]Campbell G S, Norman J M. An Introduction to Environmental Biophysics[M]. New York: Springe, 1998.
[18]Allen R G, Pereira L S, Raes D, et al. Crop evapotranspiration: guidelines for computing crop water requirements[M]//FAO Irrigation and Drainage Paper 56. Rome: FAO,1998.
[19]孙迪,关德新,袁凤辉,等. 辽西农林复合系统中杨树液流速率与气象因子的时滞效应[J]. 应用生态学报,2010,21(11):2742-2748.
Sun Di, Guan Dexin, Yuan Fenghui,et al. Time lag effect between poplar's sap flow velocity and microclimate factors in agroforestry system in west Liaoning Province[J]. Chinese Journal of Applied Ecology, 2010, 21(11): 2742-2748. (in Chinese with English abstract)
[20]张俊,李晓飞,李建贵,等. 干旱荒漠区银白杨树干液流动态[J]. 生态学报,2013,33(18):5655-5660.
Zhang Jun, Li Xiaofei, Li Jiangui, et al. Sap flow dynamics ofL. ×plantation in arid desert area[J]. Acta Ecologica Sinica, 2013, 33(18): 5655-5660. (in Chinese with English abstract)
[21]马长明,马玉洁,程月明. 冀西北坝上干旱区北京杨树干液流特征及影响因素分析[J]. 水土保持学报,2017,31(6):338-344.
Ma Changming, Ma Yujie, Cheng Yueming. Characteristics and the driving forces of sap flow in stems ofin bashang area of north-west Hebei[J]. Journal of Soil and Water Conservation, 2017, 31(6): 338-344. (in Chinese with English abstract)
[22]徐丹丹,尹立河,侯光才,等. 毛乌素沙地旱柳和小叶杨树干液流密度及其与气象因子的关系[J]. 干旱区研究,2017,34(2):375-382.
Xu Dandan, Yin Lihe, Hou Guangcai, et al. Relationships between sap flow densities in tree trunks ofandand meteorological factors in the Mu Us sandland[J]. Arid Zone Research, 2017, 34(2): 375-382. (in Chinese with English abstract)
[23]鱼腾飞,冯起,司建华,等. 胡杨的夜间蒸腾:来自树干液流、叶片气体交换及显微结构的证据[J]. 北京林业大学学报,2017,39(9):8-16.
Yu Tengfei, Feng Qi, Si Jianhua, et al. Nocturnal transpiration ofauthenticated by measurements of stem sap flux, leaf gas exchange and stomatal microsturcture[J]. Journal of Beijing Forestry University, 2017, 39(9): 8-16. (in Chinese with English abstract)
[24]席本野,王烨,邸楠,等. 地下滴灌下土壤水势对毛白杨纸浆林生长及生理特性的影响[J]. 生态学报,2012,32(17):5318-5329.
Xi Benye, Wang Ye, Di Nan, et al. Effects of soil water potential on the growth and physiological characteristics ofpulpwood plantation under subsurface drip irrigation[J]. Acta Ecologica Sinica, 2012, 32(17): 5318-5329. (in Chinese with English abstract)
[25]魏新光,陈滇豫,Liu Shouyang,等. 修剪对黄土丘陵区枣树蒸腾的调控作用[J]. 农业机械学报,2014,45(12):194-202,315.
Wei Xinguang, Chen Dianyu, Liu Shouyang, et al. Effect of trim on Jujube transpiration in loess hilly region[J]. Transactions of the Chinese Society for Agricultural Machinery, 2014, 45(12): 194-202, 315. (in Chinese with English abstract)
[26]李波,郑思宇,魏新光,等. 东北寒区日光温室葡萄液流特征及其主要环境影响因子研究[J]. 农业工程学报,2019,35(4):185-193.
Li Bo, Zheng Siyu, Wei Xinguang, et al. Study on sap flow characteristics of grape and its environment influencing factors in cold regions of northeastern China[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(4): 185-193. (in Chinese with English abstract)
[27]司建华,冯起,鱼腾飞,等. 植物夜间蒸腾及其生态水文效应研究进展[J]. 水科学进展,2014,25(6):907-914.
Si Jianhua, Feng Qi, Yu Tengfei, et al. Research advances in nighttime transpiration and its eco-hydrological implications[J]. Advances in Water Science, 2014, 25(6): 907-914. (in Chinese with English abstract)
[28]尹立河,黄金廷,王晓勇,等. 陕西榆林地区旱柳和小叶杨夜间树干液流变化特征分析[J]. 西北农林科技大学学报:自然科学版,2013,41(8):85-90.
Yin Lihe, Huang Jinting, Wang Xiaoyong, et al. Characteristics of night time sap flow ofandin Yulin, Shaanxi[J]. Journal of Northwest A&F University: Natural Science Edition, 2013, 41(8): 85-90. (in Chinese with English abstract)
[29]Yu T F, Feng Q, Si J H, et al. Depressed hydraulic redistribution of roots more by stem refilling than by nocturnal transpiration forOliv. in situ measurement[J]. Ecology & Evolution, 2018, 8(5): 2607-2616.
[30]孔喆,陈胜楠,律江,等. 欧美杨单株液流昼夜组成及其影响因素分析[J]. 林业科学,2020,56(3):8-20.
Kong Zhe, Chen Shengnan, Lü Jiang, et al. Characteristics ofsap flow over day and night and its influencing factors[J]. Scientia Silvae Sinicae, 2020, 56(3): 8-20. (in Chinese with English abstract)
[31]Dawson T E, Burgess S S O, Tu K P, et al. Nighttime transpiration in woody plants from constrasting ecosystem[J]. Tree Physiology, 2007, 27(4): 561-575.
[32]Moore G W, Cleverly J R, Owens M K. Nocturnal transpiration in riparianthickets authenticated by sap flux, eddy covariance and leaf gas exchange measurements[J]. Tree Physiology, 2008, 28(4): 521-528.
[33]de Dios V R, Roy J, Ferrio J P, et al. Processes driving nocturnal transpiration and implications for estimating land evapotranspiration[J]. Scientific Reports, 2015, 5: 10975.
Responses of tree stem sap flow and its main influencing factors to bud pruning
Li Guangde1, Zhang Yaxiong2,3, Deng Tan4, Li Doudou3, Liu Jinqiang3, Jia Liming3, Xi Benye3※
(1,,,100039,; 2,741020,; 3.,,100083,; 4.,450045,)
Stem sap flow rising through the xylem is an important physiological index to measure water use and transport of plants, and it can better reflect the plants transpiration and water metabolism process. Pruning buds after planting is a kind of economic and effective tending measure widely used in crops, economic forest, and timber trees. It can avoid a large number of lateral buds germination, promote plant growth, and improve the yield and quality of target products. The purpose of this study was to investigate the effects of bud pruning intensity on sap flow velocity characteristics and its main environmental factors of. Four treatments of bud pruning were designed. After afforestation in 2016, the buds below 1/3 (1.0 m, BP1.0, mild), 1/2 (1.5 m, BP1.5, moderate) and 2/3 (2.0 m, BP2.0, severe) of tree height were removed respectively. A control treatment (CK) without removing any buds was also designed. The sap flow velocity ofand environmental factors with different bud pruning intensities were continuously determined from August 2017 to June 2018 by using thermal diffusion flow detection technology and automatic weather station. The results showed that: 1) The diurnal dynamics of sap flow velocity showed a single peak curve in sunny days, which increased rapidly from 5:30 to 9:40, reached the peak at 11:00 to 14:40 and then decreased gradually, and began to run at low value at night from 16:30 to 22:00. The shape of diurnal dynamic curve of sap flow was not affected by the bud pruning intensities. 2) The effect of bud pruning intensity on peak value, daily average value and other characteristic values of sap flow velocity changed with time. No significant difference was found in the characteristic values of each treatment in the early stage of the experiment (August and September 2017). As time went on, the maximum values of daily and monthly average of sap flow velocity shifted from CK to BP1.0. The mean value of sap flow velocity was always the smallest under severe bud pruning (BP2.0). 3) The effect of bud pruning on average sap flow velocity was significant (<0.01). The average sap flow velocity values of CK, BP1.0, BP1.5, BP2.0were 1.405×10-3, 1.503×10-3, 1.462×10-3, 1.033×10-3cm/s, respectively from August 2017 to June 2018. And the sap flow velocity of BP2.0was significantly lower than that of the other treatments, but there was no significant difference among CK, BP1.0and BP1.5(>0.05). 4) The main environmental factors affecting sap flow velocity in different periods and treatments were different. Air temperature and radiation were the main factors affecting the instantaneous value during daytime, while vapor pressure deficit, radiation and air humidity were the main factors affecting the instantaneous value at night. However, for the mean sap flow velocity, radiation and maximum air temperature were the main drivers of daytime mean values, and minimum air temperature and air humidity were the main drivers of nighttime mean values. 5) The nighttime sap flow velocity of youngaccounted for 5%-70% of the whole day sap flow velocity under different bud pruning intensities. The nighttime sap flow velocity of different treatments showed significant differences from the third growing season after afforestation. To sum up, bud pruning did not affect the diurnal process of sap flow, but affected the pattern of sap flow and the main environmental factors. With the increase of bud pruning intensity, the smaller average daily sap flow velocity would result in the greater proportion of sap flow velocity at night. From the point of view of promoting plant water physiological activities, the light and moderate bud pruning methods are more suitable.
transpiration; environment; temperature; water consumption; sap flow; bud pruning;
李广德,张亚雄,邓坦,等. 树干液流及其主要影响因子对摘芽强度的响应[J]. 农业工程学报,2021,37(5):131-139.doi:10.11975/j.issn.1002-6819.2021.05.015 http://www.tcsae.org
Li Guangde, Zhang Yaxiong, Deng Tan, et al. Responses of tree stem sap flow and its main influencing factors to bud pruning[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(5): 131-139. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2021.05.015 http://www.tcsae.org
2020-10-09
2020-12-10
“十三五”国家重点研发计划项目(2016YFD0600403);国家自然科学基金面上项目(32001304、31800531、31670625);国家开放大学优秀青年教师培养计划
李广德,博士,副教授,研究方向为人工林高效栽培理论与技术。Email:guangdeli@ouchn.edu.cn
席本野,博士,副教授,博士生导师,研究方向为人工林水分关系。Email:benyexi@bjfu.edu.cn
10.11975/j.issn.1002-6819.2021.05.015
S792.117
A
1002-6819(2021)-05-0131-09
猜你喜欢
军事文摘(2021年16期)2021-11-05
科学导报(2019年19期)2019-09-23
智富时代(2019年4期)2019-06-01
智富时代(2019年4期)2019-06-01
数学大世界(2018年35期)2018-02-22
发明与创新·中学生(2017年5期)2017-05-12
小猕猴学习画刊(2017年1期)2017-02-17
小猕猴学习画刊(2017年1期)2017-02-17
储能科学与技术(2014年5期)2014-02-27
储能科学与技术(2014年1期)2014-02-27
