
鱼-菜共生养殖模式对罗非鱼生长性能、水质和浮游生物的影响
2021-05-06 13:06刘邦辉方彰胜丁悦秀
江苏农业科学 2021年6期
关键词:罗非鱼
刘邦辉 方彰胜 丁悦秀


摘要:以尼罗罗非鱼为研究对象,采用鱼-菜共生养殖技术对6口池塘(A组:试验组,3口池塘;B组:对照组,3口池塘)罗非鱼生长性能、水质影响和养殖效益进行研究。结果显示,在196 d养殖周期内,鱼-菜共生模式(A组)下罗非鱼的生长速度、成活率、肥满度分别显著提高18.9%、13.3%和37.0%(P<0.05),饲料系数降低10.6%(P<005);在水质改良方面,试验组COD、浊度、亚硝酸盐、总氨氮及溶解性正磷酸盐的含量与对照组(B组)相比均具有显著差异(P<005),A组池塘浮游植物密度和浮游动物密度均显著低于B组(P<0.05),但浮游动物生物量显著高于B组(P<005)。同时,A组浮游植物Shannon-Weaver多样性指数(H)和Pielou均匀度指数(J)均显著高于对照池塘(P<005)。表明鱼-菜共生养殖技术,具有提高罗非鱼生长性能、提高养殖池塘浮游生物多样性、改善养殖水质、降低水体污染和增产增值等优点,不失为一种生态健康高效的精准池塘养殖模式。
关键词:罗非鱼;鱼菜共生;精准池塘;养殖模式
多年来,作为传统的池塘养殖国家,我国池塘养殖面积和水产品产量均居世界第一[1],依靠技术进步和模式升级,高密度集约化养殖已成为我国池塘养殖的主要养殖模式[2-3]。随之产生大量的池塘沉积有机物,进而致使池塘水体呈富营养态,排出后甚至会导致附近水源或水资源恶化[4-5]。因此,探索具有增值、高效、環保的新型养殖模式以维持水产养殖增产生态协同发展已成为近年来科技工作者新的研究课题[6-7]。
鱼-菜共生系统是指将高密度水产养殖与水耕栽培(hydroponics)有机结合,形成生物浮床,利用水生植物生长过程中需要吸收大量氮、磷等元素的原理,达到去除或降低池塘水体中的沉余有机物含量以改善水质收获蔬菜的目的,进而形成营养盐良性循环的养殖-种植系统[8]。已有研究表明,鱼-菜共生可以使养殖生物、植物和微生物等系统生物与生态因子达到一种平衡状态,促使养殖水体呈现稳态,进而为养殖水体的原位修复提供有效途径[9-10]。
罗非鱼是我国重要的池塘养殖经济鱼类之一,其肉质鲜美、刺少、蛋白质含量高,近年已成为我国养殖、加工、出口的优质鱼类[11]。本研究以罗非鱼为研究对象,采用鱼菜共生-生物浮床技术对6口池塘(A组:试验组;B组:对照组)罗非鱼生长性能、水质和浮游生物进行研究,以期为鱼菜共生-生物浮床技术在水产养殖中的应用提供技术参数。
1 材料与方法
1.1 试验材料
试验在广东省江门市新会区开源农业有限公司罗非鱼养殖场进行,时间为2018年4月17日至10月30日。试验分为2组,每组3个重复,总计池塘6口,每口池塘面积为2 000 m2,养殖水源为西江水,池塘水深为1.8~2.0 m,所有试验养殖池塘均配置2台叶轮式增氧机,功率为1.5 kW。试验用鱼为佛山市梁氏集团提供的无病无伤尼罗罗非鱼(Oreochromis niloticus),平均体质量为(20.0±12) g/尾,试验用菜为江门市新会市开源农业有限公司提供的蕹菜(Ipomoea aquatica Forsk.),又称空心菜。
1.2 试验方法
试验分为2个组、各3个重复试验(A组、试验组;B组、对照组),每组试验鱼放养规格和密度分别为(20±1.2)g/尾和(19.0±0.1)×104尾/hm2,组间无显著差异。其中,A组池塘水面每隔4 m放置2 m宽浮排结构的装置并种植蕹菜(图1、图2),共6排,B组池塘无放置。
1.2.1 养殖管理 试验期间,尼罗罗非鱼养殖、投料及日常管理参照开源农业有限公司养殖场罗非鱼池塘养殖企业标准进行,试验饲料由恒兴饲料提供,每天06:00—08:00、14:00—18:00各投喂饲料1次,投喂量为鱼质量的3%,试验周期为126 d。试验期间,记录各池塘放养规格、放养量、饲料投喂量等。试验完成时,统计鱼体规格、总质量、成活率、饲料利用率等。
1.2.2 养殖水质因子测定 每周一07:00—08:00采用ET 9908便携式多参数水质测定仪对试验养殖水体的温度(T)、pH值、溶解氧(DO)含量、浊度(Tur)进行定点测定;透明度(Scc)采用萨氏盘法进行测定。采集各试验池塘30 cm处水体用于水质因子测定,其中,采用纳氏试剂分光光度法(HJ 535—2009)测定TAN含量;采用分光光度法(GB 7493—1987)测定NO-2含量;采用紫外分光光度法(HJ/T 346—2007)测定NO-3含量;采用重铬酸盐法(GB 11914—1989)测定COD含量;采用磷钼蓝分光光度法(HJ593—2010)测定溶解性正磷酸盐(SRP)含量。
1.2.3 浮游生物水样采集与测定 浮游生物的采样时间和次数与水质因子测定水样一致。采用《内陆水域渔业自然资源调查手册》方法进行浮游生物采样、计数及种类鉴定。采集前配制鲁哥氏液用以固定水样。在各试验池塘选择5个采样点(2对角线上选择4个点和中心点)分别采集1 000 mL水体,取500 mL混合水样用于测定浮游植物、原生动物和轮虫等等浮游生物量,取1 000 mL混合水样经浮游生物过滤网过滤后用于枝角类和桡足类测定。所有样品经沉淀浓缩后在显微镜下观察计数。浮游生物多样性按以下公式计算:
1.3 计算公式
1.4 数据处理
试验数据均采用SPSS 17.0处理,采用“平均值±标准差”表示。在单因子方差分析(A-NOVA)基础上采用t检验进行分析比较,取95%置信区间(P<0.05)或99%置信区间(P<0.01)。
2 结果与分析
2.1 罗非鱼生长特性及养殖效益分析
由表1可知,与对照组相比,试验组的特定生长率、肥满度、成活率和净产量分别提高了18.90%、0.37%、13.30%和30.70%,饲料系数降低了1060%,均呈现显著差异(P<0.05)。鱼-菜共生养殖设施费用0.3万元,使用周期为3年,平均每年折旧费0.1万元。试验组比对照组净增值0.5万元/1 000 m2(鱼时价9.8 元/kg,饲料时价6.2元/kg)。结果显示,鱼-菜共生养殖技术不仅可提高罗非鱼养殖成活率和特定生长率,还可显著提高饲料利用率进而促进增产增值,不失为一种生态、高效的养殖方式。
2.2 池塘水质因子浓度变化
由表2可知,试验周期内,与对照组相比,试验组养殖水体水温和硝酸盐平均浓度无显著差异(P>0.05),试验组养殖水体pH值、Scc、DO等水环境因子数值均显著高于对照组(P<0.05),试验组养殖水体Tur、TAN、NO-2、COD和SRP等水质因子分别降低了34.34%、18.31%、52.24%、39.7%和38.69%,也均呈现出显著差异(P<0.05)。试验结果表明,与对照组相比,试验组在相同饲养管理的前提下,鱼-菜共生对罗非鱼池塘养殖水体水质因子浓度水平的调节和生态系统的稳定具有较高的效率。
2.3 浮游生物种类组成及百分比
在养殖试验周期内,通过对池塘养殖水体浮游生物种类的鉴定,大致检测出浮游动物24种(属),其中原生动物为10种(属)、轮虫为10种(属)、枝角类为2种(属)、桡足类为2种(属)。检测出浮游植物48種(属),其中绿藻门为19种(属)、硅藻门为12种(属)、蓝藻门为8种(属)、裸藻门为4种(属)、隐藻门为2种(属)、甲藻门为3种(属),2组池塘的优势种群基本相同。
通过对2组池塘养殖水体浮游生物统计(表3)得知,2组罗非鱼养殖池塘内的浮游植物均以蓝藻、绿藻和硅藻为主,3种藻类总和分别占浮游植物的88.2%和89.1%,在各组藻间除绿藻、甲藻百分比无显著差异外(P>0.05),其余藻类均有显著差异(P<005);浮游动物以原生动物为主,试验组和对照组分别占总浮游生物数量的91.1%和85.4%,在各组原生动物间,除原生动物和枝角类间百分比无显著差异外(P>0.05),桡足类和卤虫所占百分比较小,但2组间呈显著差异(P<0.05)。试验结果表明,鱼-菜共生模式不但有效地抑制了蓝藻的繁殖,促进可被罗非鱼有效利用的硅藻种群数量,还提高了维持养殖水体稳态的原生动物的种群数量,进一步提高了养殖水体的生态平衡。
2.4 浮游生物密度与生物量变化
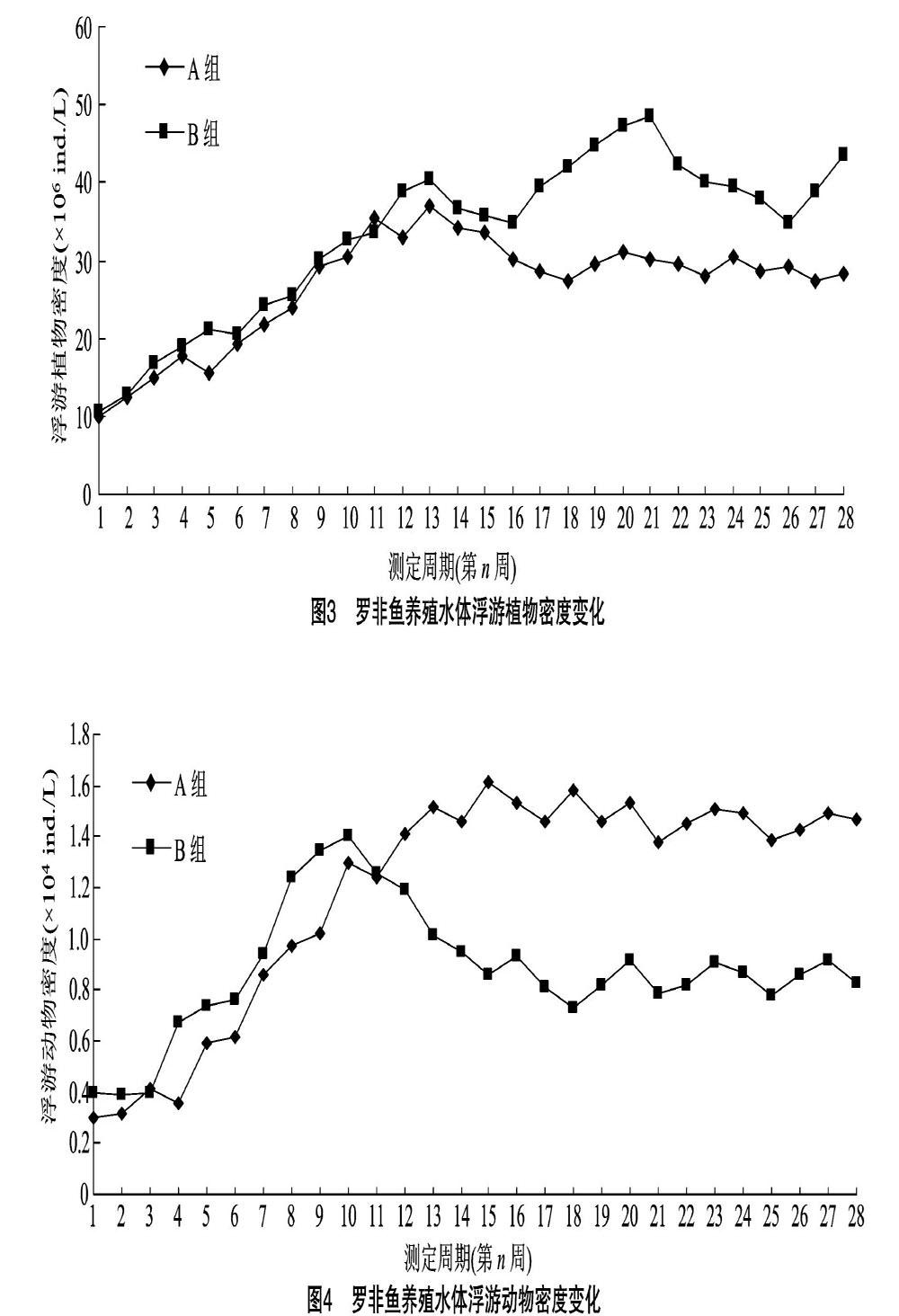
试验周期内2组池塘养殖水体浮游植物密度变化见图3。由图3可见,在试验前期,随着气温升高和养殖饲料投喂量的增加,2组池塘养殖水体中浮游植物密度除在前5~6周时出现较小的下降变化外,整体增长较快并在第13周达到增长拐点。在试验周期13~28周内,试验组和对照组养殖水体浮游植物密度分别在27.3×106~31.2×106 ind./L 和34.8×106~48.6×106 ind./L之间波动,养殖水体的浮游植物密度均值分别为30.525×106 ind./L 和39.125×106 ind./L。试验结果表明,鱼-菜共生养殖模式对罗非鱼养殖水体浮游植物密度有较好的稳定作用,进而促进形成稳态养殖水体。
试验周期内2组池塘养殖水体浮游动物密度变化见图4。由图4可知,在试验期内,随着气温升高和养殖饲料投喂量的增加,2组池塘养殖水体中浮游动物密度除在前4周内无显著变化外,整体呈线性增长并在第9周达到增长变化临界点,这可能与原生动物的数量变化有关。在试验周期10~28周内,试验组和对照组养殖水体浮游动物密度呈现相反的变化波动,且两者的变化分别在1.02×104~158×104 ind./L和0.73×104~1.4×104 ind./L 之间波动。该结果表明,鱼-菜共生养殖模式可间接改变养殖水体浮游动物种群数量变化。
2.5 浮游生物多样性的变化
试验周期内2组养殖水体浮游生物多样性指数见表4。由表4可知,试验前期,2组池塘养殖水体浮游生物多样性及均匀度指数变化不明显,试验中后期变化差异较大。整个试验周期内,除浮游动物的Pielou 均匀度指数(J)无显著差异外(P>0.05),试验组浮游植物Shannon-Weaver多样性指数(H)与 Pielou 均匀度指数(J)和浮游动物 Shannon-Weaver多样性指数(H)均显著高于对照组(P<005)。
3 讨论
(1)鱼-菜共生池塘养殖模式对罗非鱼生长特性的影响。陈家长等研究表明,鱼-菜共生池塘养殖模式对养殖池塘水质改善和经济效益提高方面具有显著促进作用[8,10],随着研究的深入,鱼-菜共生池塘养殖模式在养殖水体富营养化治理、水体健康评价和水质生物监测等方面具有非常广阔的应用前景。该试验数据显示,与普通组相比,鱼-菜共生试验组罗非鱼特定生长率、肥满度、成活率和净产量分别提高了18.90%、0.37%、13.30%和3070%,饲料系数降低了10.60%,同时净增值 0.5万元/1 000 m2[12-16]。本试验数据显示,鱼-菜共生养殖技术不仅可提高罗非鱼养殖成活率和特定生长率,还可显著提高饲料利用率进而达到环保、增值效果,这与陈家长等[10,17]等对养殖池塘研究结果一致,不失为一种高效健康养殖模式。
(2)鱼-菜共生养殖模式对罗非鱼养殖水体水质的调控。大量研究表明,生物浮床或者有根植物能够有效去除水体中总氮、BOD、SS、总磷、氨氮和重金属等污染物[18-21]。本试验鱼菜共生-生物浮床研究结果显示,在对照组与试验组之间除养殖水体水温和硝酸盐平均浓度无显著差异外(P>0.05),试验组养殖水体pH值、透明度、溶解氧含量等水质因子均不同程度地显著高于对照组(P<0.05),试验组养殖水体浊度、氨氮、亚硝酸盐、化学需氧量和溶解性正磷酸盐等水质因子均显著降低(P<0.05)。试验研究结果表明,在相同饲养管理的前提下,对池塘养殖水体水质因子浓度的调节和生态系统的稳定具有较高的效率,该结果与曹煜成等[11,15]有关鱼菜生物浮床对养殖水体调控的研究基本一致。
已有学者研究表明,因生物浮床栽培的水生植物生长在水面,影响水体气液相接触面积,进而降低养殖池塘水体空气溶解量[22]。此外,植物的遮阳作用,阻碍水体藻类等沉水植物光合作用的同时,营呼吸作用的植物根系及其周围营硝化作用的微生物仍会消耗氧气,进一步导致养殖水体“氧源”减少[23]。也有部分学者认为,当生物浮床覆盖面积小于池塘养殖水体面积的20%时,对池塘养殖水体溶解氧含量的变化有影响,但不显著,覆盖面积降低至7.5%时,养殖池塘水体的溶氧量无差异[10-11]。本试验结果显示,试验养殖池塘中水体的溶解氧含量有所增加,与上述研究结果相悖,其可能原因是在低覆盖率条件下,鱼-菜共生系统改变了养殖池塘水体中微生物的种群和密度,促使水体生态系统形成微生态平衡,进而达到水体稳态。这种“综合效应”进一步改善了养殖水体的营养盐流通,降低了含氮化合物含量及溶解氧消耗,具体原理仍需进一步研究。
(3)鱼-菜共生养殖模式对罗非鱼养殖水体浮游动植物密度的影响。浮游生物的种类组成、群落结构和丰度变化,直接影响水体水质、系统内物质能量流和生物资源演替[24],池塘系统内浮游生物群落由多种原生动物和浮游植物种类构成[25],浮游生物群落种群能随外界环境的改变而做出规律性的变化[26-30],形成复杂和多样性的群落组合,呈现出不同的生物多样性指数[25-27],进而促进养殖池塘系统内的能量流动、物质循环和信息传递[31-32]。所以,养殖池塘浮游生物的种类组成、密度及多样性特征可以作为养殖水体健康状况的指示[33-34]。
浮游生物多样性指数通常用以反映养殖水体浮游生物群落结构组成,或富营养化对养殖水生生物的影响[27-29]。Shannon-Wiener 指数表示浮游生物群落组成,指数值越大预示着浮游生物群落结构越复杂,养殖池塘水体稳定性也越大[35]。均匀度是多样性指数实际与理论的比值,描述水体中浮游生物种类、数量的均匀程度,多用于水体浮游生物多样性评估[36]。优势种种类和密度对水体群落结构的稳定性有重要作用,优势种种类多且优势度越小,则群落结构相对稳定[37]。本试验结果表明,2组池塘中原生动物的密度均呈现出先上升后下降、最终趋于稳定的形态,试验组池塘浮游动物多样性显著高于普通组,这与已有的研究结果基本一致。浮游动植物多样性指数和均匀性指数均高于普通组,这与邓金钗等罗非鱼混养池塘研究中浮游动植物多样性指数[38]有部分差异,可能是由于单养和混养主体生物的差异引起,具体原因仍需进一步研究。
鱼-菜共生模式能够在一定程度上影响池塘浮游生物物群落结构,丰富池塘浮游生物多样性,改善养殖水质,促进养殖水体趋于稳定和健康,不失为一种生态、高效的池塘养殖模式。
参考文献:
[1]徐 皓,刘兴国,吴 凡. 池塘养殖系统模式构建主要技术与改造模式[J]. 中国水产,2009,9(8):7-9.
[2]Piedrahita R H. Reducing the potential environmental impact of tank aquaculture effluents through intensification and recirculation[J]. Aquaculture,2003,226(1):35-44.
[3]董双林. 高效低碳——中国水产养殖业发展的必由之路[J]. 水产学报,2011,35(10):1595-1600.
[4]王 峰,雷霁霖,高淳仁,等. 国内外工厂化循环水养殖研究进展[J]. 中国水产科学,2013,20(5):1100-1111.
[5]王彦波,许梓荣. 微生物对水产养殖环境的生物修复作用[J]. 饲料研究,2004(12):42-43.
[6]Barak Y,Cytryn E,Gelfand I,et al. Phosphorus removal in a Marine prototype,recirculating aquaculture system[J]. Aquaculture,2003,220(1):313-326.
[7]Pedersen L F,Meinelt T,David L. Straus peracetic acid degradation in freshwater aquaculture systems and possible practical implications[J]. Aquacult Eng,2013,53:65-71.
[8]丁永良,张明华,张建华,等. 鱼菜共生系统的研究[J]. 中国水产科学,1997,4(增刊1):71-76.
[9]常会庆,寇太记,乔鲜花,等. 几种植物去除污染水体中养分效果研究[J]. 水土保持通报,2009(5):118-122.
[10]陈家长,孟顺龙,胡庚东,等. 空心菜浮床栽培对集约化养殖鱼塘水质的影响[J]. 生态与农村环境学报,2010,26(2):155-159.
[11]贺艳辉,张红燕,龚贇翀,等. 我国罗非鱼出口面临的机遇与挑战[J]. 中国渔业经济,2009,27(3):85-89.
[12]賈 悦,李秀珍,唐莹莹,等. 不同采收方式对富养化河道浮床空心菜生物产出的影响[J]. 生态学杂志,2011,30(6):1091-1099.
[13]周小平,王建国,薛利红,等. 浮床植物系统对富营养化水体中氮、磷净化特征的初步研究[J]. 应用生态学报,2005,16(11):2199-2203.
[14]由文辉,刘淑媛,钱晓燕. 水生经济植物净化受污染水体研究[J]. 华东师范大学学报(自然科学版),2000,3(1):99-102.
[15]杨晓玲,郭金耀. 水蕹菜对富营养化养殖水的净化作用研究[J]. 作物杂志,2012(1):49-52.
[16]陈玉红,刘忠良. 猪场厌氧废水用于空心菜水培试验研究[J]. 农业环境与发展,2011(3):87-90.
[17]谷孝鸿. 不同养殖类型池塘浮游生物群落结构的初步分析[J]. 湖泊科學,1994,6(3):276-282.
[18]周真明,陈灿瑜,叶 青,等. 浮床植物系统对富营养化水体的净化效果[J]. 华侨大学学报(自然科学版),2010(5):576-579.
[19]吕锡武,宋海亮. 水培蔬菜法对富营养化水体中氮磷的去除特性研究[J]. 江苏环境科技,2004,6(2):1-3.
[20]操家顺,李欲如,陈 娟. 水蕹菜对重污染河道净化及克藻功能[J]. 水资源保护,2006,22(2):36-38,41.
[21]Yin Z,Chen J,Zeng L,et al. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight[J]. Molecular Plant-microbe Interactions:MPMI,2000,13(8):869-876.
[22]任照阳,邓春光. 生态浮床技术应用研究进展[J]. 农业环境科学学报,2007(增刊1):261-263.
[23]马井泉,周怀东,董哲仁. 我国应用生态技术修复富营养化湖泊的研究进展[J]. 中国水利水电科学研究院学报,2005,3(3):209-215.
[24]赵 文. 刺参池塘养殖生态学及健康养殖理论[M]. 北京:科学出版社,2009.
[25]李丹丹. 不同模式罗非鱼养殖池塘浮游生物群落结构动态变化[D]. 南京:南京农业大学,2015:1-7.
[26]李志斐,王广军,陈鹏飞,等. 生物浮床技术在水产养殖中的应用概况[J]. 广东农业科学,2013,40(3):106-108,114.
[27]李胜男,周 建,魏利军,等. 淡水超微型浮游植物多样性及其研究方法[J]. 生态学杂志,2015,34(4):1174-1182.
[28]Rakesh R. Yeredla,Xu H F.incorporating strong polarity minerals of tourmaline with semiconductor Titania to improve the photosplitting of water[J]. The Journal of Physical Chem-istry C,2008,112(2):532-539.
[29]Xu H L,Cao Q,Wang X Y,et al. Properties and chemical oxidation polymerization of polyaniline/neutral red/TiO2 composite electrodes[J]. Materials Science and Engineering:B,2010,171(1):104-108.
[30]Yuan Y,Ahmed J,Kim S H. Polyaniline/Carbon black com-posite-supported Iron phthalocyanine as an oxygen reduction catalyst for microbial fuel cells[J]. Journal of Power Sources,2011,196(3):1103-1106.
[31]李 今,吕 田,华江环. 人工浮床水培空心菜生长特性及其在养殖废水净化中的应用[J]. 湖南师范大学自然科学学报,2014,37(2):22-27.
[32]胡绵好. 水生经济植物浮床技术改善富营养化水体水质的研究[D]. 上海:上海交通大学,2008.
[33]Tavernini S,Mura G,Rossetti G. Factors influencing the seasonal phenology and composition of zooplankton communities in mountain temporary pools[J]. International Review of Hydrobiology,2005,90(4):358-375.
[34]Echaniz S A,Vignatti A M,de Paggi S J,et al. Zooplankton seasonal abundance of South American saline shallow lakes[J]. International Review of Hydrobiology,2006,91(1):86-100.
[35]Lai B,Wang P,Li H,et al. Calcined polyaniline-iron composite as a high efficient cathodic catalyst in microbial fuel cells[J]. Bioresource Technology,2013,131(0):321-324.
[36]Rao S M,Xing Y C. Simulation of nanostructured electrodes for polymer electrolyte membrane fuel cells[J]. Journal of Power Sources,2008,185(2):1094-1100.
[37]Logan B E,Murano C,Scott K,et al. Electricity generation from cysteine in a microbial fuel cell[J]. Water Research,2005,39(5):942-952.
[38]邓金钗,熊邦喜,张小敏,等. 池塘主养草鱼三种养殖模式浮游植物群落结构的比较[J]. 淡水渔业,2012,42(5):63-67. 周 聃,崔雁娜,周冬仁,等. 不同养殖模式下加州鲈鱼肉质比较分析[J]. 江苏农业科学,2021,49(6):146-149.
猜你喜欢
当代水产(2023年2期)2023-05-19
当代水产(2022年6期)2022-06-29
今日农业(2021年15期)2021-11-26
当代水产(2021年7期)2021-11-04
当代水产(2020年3期)2020-06-15
当代水产(2019年4期)2019-05-16
当代水产(2018年8期)2018-11-02
当代水产(2018年12期)2018-05-16
农家之友(2018年12期)2018-03-12
农家科技中旬版(2016年12期)2016-04-16
