
巴西橡胶树5个主栽品种幼苗叶片的解剖结构及光合特性研究
2021-05-03 11:25高新生胡欣欣李维国
西南林业大学学报 2021年3期
高新生 胡欣欣 李 廷 李维国 黄 肖
(1. 中国热带农业科学院橡胶研究所,海南 海口 571101;2. 国家橡胶树育种中心,海南 海口 571101;3. 海南大学,海南 海口 570228;4. 海南热带海洋学院,海南 三亚 572022)
在不同的光照强度下,植物通过长期的进化,使自身的形态结构和其生理过程变得更加适应外界环境,从而得到更好的发展。叶片作为植物进行光合作用的主要器官以及植物进行生命活动的主要部位,其解剖结构和生长都会随着光环境的变化自发的发生适应性的变化[1]。因此研究叶片的解剖结构,探索其光合生理特性具有重要的科学和实践价值。
王少先等[2]、廖文婷等[3]、索巍巍[4]研究表明,即使在同一环境下,不同的物种以及相同物种的不同品系间的光合速率均存在差异。巴西橡胶树(Hevea brasiliensis)作为原产于巴西亚马逊河的高大乔木,在长期的栽培驯化过程中,其叶片结构和光合特性具有与热带生境相匹配的特征[5-6]。而经过多年品种选育,不同遗传背景的品系、相同品系不同物候间的净光合速率、光能利用率、水分利用率等均存在明显的差异,如传统主栽品种PB86、热研8813等高产品系净光合速率较高,部分抗寒品系则较低[7-8];而作为反映植物光合作用与环境关系内在探针的叶绿素荧光方面研究也佐证了巴西橡胶树品系间在光合潜能、光适应能力方面存在较大差异[9-11];胡耀华等[12]运用光合模型从群体光合角度证实了不同基因型间光合总量存在差异和季节性波动;同时,研究表明,巴西橡胶树叶片净光合速率的日变化是受到了非气孔因素的存在,即“午休”现象的干扰,而影响叶片净光合速率的主要生态因子是光合有效辐射和大气相对湿度[13],并且即使是同一基因型当中,由于光照、温度、土壤环境、农艺措施等因子等的显著不同,基因型与环境存在较强互作,导致其高、低产材料间光饱和点等重要光合特征参数也存在较大差异[14]。
植物叶片是其进行光合作用的物质载体,对环境因子变化敏感,且变异及可塑性较大,其海绵组织、栅栏组织、上下表皮的厚度等组织结构的发育成熟程度对光合作用起着至关重要的作用。巴西橡胶树中,不同品种间、同品种不同环境和不同倍性材料中叶片解剖结构均存在着很大的差异,植物叶片厚度、栅栏组织、海绵组织等均会受到环境因子的影响而发生相应的变化,对光照强度、抗逆性均存在不同适应性[15-17]。但目前,对巴西橡胶树特定品种如热垦628等潜力主栽新品种叶片解剖结构方面的研究基本未见报道,其对光合效能的具体影响仍有待进一步明晰。
本研究对部分巴西橡胶树主栽品种的叶片解剖结构和光合生理相关因子进行了观测,旨在探索巴西橡胶树主栽品种叶片结构变化与光合生理特征的关系,为巴西橡胶树高光效基因型筛选,抗逆品种选育与抗逆栽培技术研发提供参考依据。
1 材料与方法
1.1 试验地概况与材料来源
试验地点位于海南省儋州市中国热带农业科学院第九试验区(39°54′N,116°28′E),试区属典型热带海岛季风气候区,年均温21.5~28.5 ℃,全年日平均气温≥10 ℃,积温为8 500~9 100 ℃;历年平均降雨量为1 607 mm,主要分布在7—9月,占全年降雨量的70%以上。试区土壤为典型砖红壤,层厚度约200 cm。
参试品种包括热研106、热垦628、IAN873、RRIM600和PR107共5个,均为我国当前生产中重要栽培种。试验材料为1年生大田定植常规管理的幼苗,砧木为GT1种子苗。从品种类型看,PR107、RRIM600为传统的高产和抗逆主栽品种,热研106为早熟高产品种,热垦628为胶木兼优品种,IAN873为抗平流型寒害的中产品种。从系谱看,RRIM600为Tjir1和PB86杂交的品种,PR107为初生代无性系,热研106与PR107,RRIM600亲缘关系较近,IAN873与热垦628亲缘关系较近。
1.2 试验方法
1.2.1 叶片样品采集
在2018年8月,每个参试无性系选择非边行的3株生势一致健壮植株,每株选择当年生树冠中上部外围不同方位的向阳、健康、成熟的叶片12片,并分为4组,每组选定不同方向的3张叶片,分别用于光合特性、SPAD值、光合色素与叶片解剖特征观测。
1.2.2 测定方法
光合特性测定利用Li-6400XT光合测定仪(Li-COR,美国),采用LED红蓝光源叶室,光合有效辐射(PAR)设置为1 500 μmol/(m2·s),气体流速设定为500 μmol/s,温度25 ℃。于2017年8月(天气晴朗)的9:00—11:00观测,每隔3 d测定1次,共观测3次取平均值,测定指标包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)等。水分利用效率(WUE)为瞬时净光合速率与蒸腾速率的比值。叶肉瞬时羧化效率为瞬时净光合速率与胞间二氧化碳的比值。
叶绿素含量的测定方法为95%乙醇提取法[18]。测定指标有叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(Chl)、总类胡萝卜素(Car)、SPAD值。
叶解剖特征采用石蜡切片法(材料1.0 cm×0.5 cm,切片厚度为12 μm)[19]。观测叶片厚度(LT)、上表皮厚度(UET)、下表皮厚度(LET)、栅栏组织厚度(PT)、海绵组织厚度(ST)、组织结构紧密度(栅栏组织厚度与叶片厚度比值,PT/LT)、组织结构疏松度(栅栏组织厚度与海绵组织厚度比值,PT/ST)、栅海比(ST/LT)、瞬时羧化效率(ICE)等指标。
1.2.3 数据处理与分析
数据用DPS V7.05和Excel V2010进行统计分析,数据差异显著性分析采用Duncan新复极差法。
2 结果与分析
2.1 不同品种叶片解剖结构特征比较
从叶片解剖结构看,品种间存在显著差异(表1)。其中,叶片厚度在137.25~150.05 μm,上、下表皮厚度分别在9.75~13.32 μm、9.51~11.66 μm,RRIM600、IAN873叶片最厚,显著厚于热研106、热垦628,其中热研106最薄;从上、下表皮厚度看,PR107、RRIM600最厚,热研106最薄,热垦628、IAN873介于中间。从栅栏、海绵组织看,RRIM600、IAN873最厚,热垦628、热研106最薄。栅海比介于0.93~1.15,RRIM600、IAN873与热研106栅海比较大;而组织结构疏松度、致密度分别介于0.40~0.44、0.41~0.45,其中RRIM600、IAN873、热研106组织结构致密度较高,而热垦628致密度低。整体来看,RRIM600、IAN873不仅叶片厚,且致密度高,而PR107叶片厚度及致密度中等,热垦628叶片厚度及致密度均较差,而热研106虽然叶片最薄,但叶片致密度较高。

表 1 叶片解剖结构的比较分析Table 1 Comparison of leaf anatomic characteristics
2.2 不同品种叶片光合气体交换参数
由表2可知,品种间的净光合速率为11.21~17.41 μmol/(m2·s),其中最高与最低的品种分别为RRIM600与热研106,且品种间存在极显著差异,从蒸腾速率看,IAN873蒸腾速率较大,PR107蒸腾速率小,二者差异显著;从二者比值所得的水分利用率看,结果基本与蒸腾速率结果相反,最高的为PR107,最低的为IAN873,二者间差异显著。从气孔导度看,PR107、RRIM600的气孔导度、胞间二氧化碳浓度较高,极显著高于热研106,胞间CO2浓度结果与之相似。从叶肉瞬时羧化效率看,效率最高的是RRIM600,其次是IAN873,最低的是PR107,且彼此间存在着显著差异。
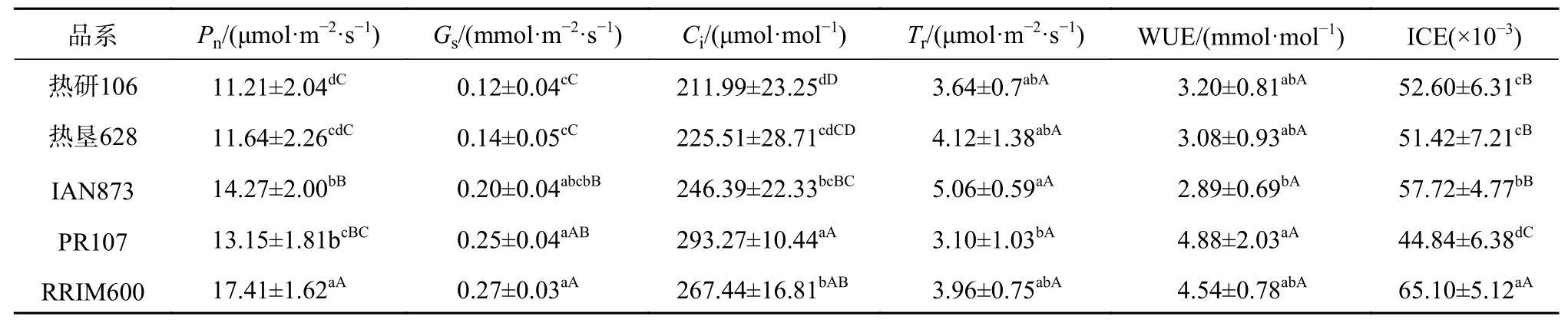
表 2 不同品种叶片光合气体交换参数Table 2 Comparison of gas exchange parameters among different clones
整体来看,净光合速率较高,效率较高的品种有RRIM600、IAN873;而净光合速率一般的品种为热垦628、热研106,品种对应的气孔导度较小,胞间二氧化碳含量也较低,说明净光合速率低可能受到了气孔因素限制。而对水分利用较好的品种有PR107、RRIM600,PR107则在气孔开放与水分利用方面较为均衡,但羧化效率不高。
2.3 不同品种叶片光合色素差异
叶绿素在光合作用的光吸收中起到核心作用,SPAD值一般与叶绿素含量呈现正相关。本研究中,不同品种间,除热垦628外,其余品种间叶绿素含量不存在显著差异,而SPAD值间PR107显著低于其他品种。其中,Chla、Chlb与Chl的范围分别为1.67~2.02、1.14~2.11、2.92~4.13 mg/mL,其中热垦628的Chl与Chla、Chlb含量明显高于其他品种,而RRIM600、热研106的Chl含量较低,但与其他品种基本不存在显著差异。从Chla/Chlb值看,热研106、IAN873较高,说明该品种可能对强光的利用较好,热垦628比值最低,表征其可能对弱光的利用最好。Car范围介于0.12~0.26 mg/mL间,但彼此间均差异不显著。SPAD值范围介于51.00~58.98之间,热研106的SPAD值最高,显著高于PR107和RRIM600,但与其他2个品种无显著差异(表3)。整体来看,SPAD值与光合色素间的变化趋势不尽一致。

表 3 不同品种光合色素含量和SPAD值Table 3 Camparision of photosynthetic pigment content and SPAD values among different clones
2.4 净光合速率与叶片解剖结构的相关分析
由表4中可知,叶片净光合速率(Pn)与PT,LT极显著相关,相关系数达到了0.676、0.721,与LET显著相关,相关系数为0.561。在巴西橡胶树的绝大多数叶片中,含叶绿素的叶绿体主要位于接近上表皮的栅栏组织中,海绵组织次之,因此Pn与PT相关性较强。气孔则直接影响光合气体交换的效率,而其主要集中在叶片下表皮,这可能导致了Pn与LET显著相关。叶片由海绵组织、栅栏组织、下表皮和上表皮构成也导致叶片与之的强相关性。

表 4 净光合速率与叶片解剖特征的相关性分析Table 4 Correlation analysis between Pn and leaf anatomic characteristic
同时,本研究中的PT与ST/LT、PT/LT极显著相关,与PT/ST极显著负相关;ST/LT与PT/LT极显著正相关,与 PT/ST显著负相关,主要是由栅栏组织、海绵组织、栅海比、组织紧密度等指标的定义与计算关系所决定的。
2.5 净光合速率、叶绿素含量与SPAD值间的相关分析
由表5可知,Pn与叶绿素含量、SPAD值均无显著相关性;Chla、Chlb与Chl含量呈显著或极显著正相关,Chla与总类胡萝卜素的含量呈极显著负相关;叶片Chl含量与叶片总类胡萝卜素含量呈极显著负相关,而SPAD值与其他参数均无显著相关性。
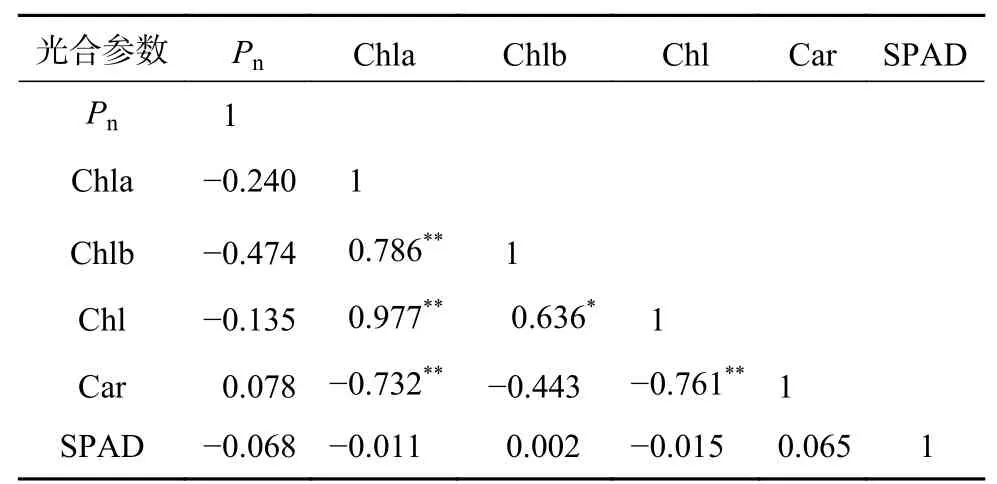
表 5 净光合速率、光合色素与SPAD值间的相关分析Table 5 Correlation analysis among Pn, photosynthetic pigment content and SPAD value
3 结论与讨论
巴西橡胶树作为典型的热带作物,其叶片要兼顾实现较高的蒸腾速率、营养与水分的运输功能。从叶片看,为异面型叶,叶片表皮均由单层细胞构成,上表皮外被角质层,下表皮外被单细胞非腺毛,上、下表皮细胞为不规则的扁长方形,排列紧密。靠近上表皮的栅栏组织细胞为长柱状,细胞分层,胞间隙较小;海绵组织细胞位于栅栏组织和下表皮之间,多为短柱状或椭球形,相互连接成网。本研究中,各品种的上表皮厚度均高于下表皮。一般而言,植物叶片结构与植株光合能力、抗性等密切相关。上表皮较厚,有利于避免强光对叶肉细胞的灼伤,下表皮较薄有利于植株进行气体交换。而植物叶片栅栏与海绵组织的分化程度(组织结构致密度)和水分利用率是反映作物适宜强光照射程度等重要参考指标[20]。本研究中,净光合速率与栅栏组织厚度极显著相关,也支持这一结论。在叶片中,如果栅栏组织发达、细胞层数增加而体积减小,海绵组织则相对减少、细胞间隙减小,一方面表征了叶片涵养水分潜力越大,是植物对水分短缺的一种响应,能更好的应对干旱的胁迫[21];另一方面,高度发达的栅栏组织既可避免叶肉组织受到强光灼伤,又可最大限度利用衍射光进行光合作用,也可增加水分从维管束到表皮间的运输效率,保证光合作用对水分的需求,从而提高叶片的光合效率[22]。
本研究中,从叶片解剖结构看,RRIM600的叶片厚度、栅海比均较高,表明其具备较强的对强光的适应能力。而IAN873的叶片厚度,上表皮、栅栏组织厚度基本显著高于PR107、热研106和热垦628,说明其光合潜力较大,对干旱胁迫的适应性概率更好。从光合性状看,品种间光合速率存在较大的差异,其中RRIM600的最大净光合速率及羧化效率较高,显著高于热研106等品种;品种间的水分利用率也存在较大差异,RRIM600、PR107对水分的利用率较高,蒸腾速率和气体交换方面较为均衡,品种间光合能力、水分利用率(WUE)的较大差异也为后续高光效与高水分利用率品种改良提供了遗传基础。同时,品种净光合速率除了与栅栏组织厚度,叶片厚度极显著相关外,与下表皮厚度也显著相关。因此,叶片栅栏组织发达,叶片较厚尤其是下表皮较厚的品种光合能力可能较强。
光合色素是光合作用的基础,其含量直接影响着光合作用速率,是反映光合强度的重要指标。在本研究中,净光合速率与叶绿素含量、SPAD值均无相关性,而叶片Chl含量与Chlb含量显著性更强。一些报道认为,相当多的植物叶片在正常情况下均含有充足的甚至过剩的叶绿素,当叶绿素含量超过某一阈值后,含量高低与净光合速率不存在必然相关[23-24],巴西橡胶树作为热带常绿乔木,叶色浓绿,本研究几个品种叶片的叶绿素含量主要介于3~4之间,而艾天成等[25]在研究不同作物叶绿素含量与SPAD值相关性研究中,水稻、棉花、高粱等的叶绿素含量则分别最高值分别为3.4、2.0和2.6,推测叶绿素含量充足可能是本研究中净光合速率与叶绿素含量不具显著相关性的重要原因。
本试验以热研106,热垦628等5个当前生产中的主栽品种幼苗为参试材料,系统观测了其光合参数与叶片解剖特征,探讨叶片结构对光合生理的影响。结果表明,不同品种间叶片解剖结构、光合参数及光合色素之间均存在着显著差异,相关分析表明,净光合速率与叶片厚度、栅栏组织厚度极显著相关,与叶绿素不显著相关。巴西橡胶树叶片厚度,栅栏组织的厚度等可作为筛选高光效基因型的重要参考指标。从品种来看,IAN873、PR107、RRIM600叶片较厚,栅栏组织发达,光合能力与水分利用率较强,RRIM600的组织致密度与水分利用率均较高。本研究为以叶片解剖特征辅助评价不同品系光合特性及水分胁迫逆境等的适应能力提供了依据,也为参试品种的配套栽培技术研发提供了参考。
猜你喜欢
热带作物学报(2022年8期)2022-09-16
今日农业(2022年3期)2022-06-05
农业工程学报(2022年4期)2022-04-24
中国农学通报(2022年6期)2022-04-15
农业与技术(2020年12期)2020-07-04
农民致富之友(2019年29期)2019-10-21
祝您健康·文摘版(2019年4期)2019-06-11
散文选刊·下半月(2018年2期)2018-02-23
新课程·下旬(2017年7期)2017-08-14
饮食科学(2016年3期)2016-07-04
