
沙地柠条光合特性与水分利用效率对干旱胁迫的响应
2021-05-03 11:25韩阳瑞许大为张伟艳
西南林业大学学报 2021年3期
韩阳瑞 单 炜 许大为 张伟艳
(1. 东北林业大学工程咨询设计研究院有限公司,黑龙江 哈尔滨 150040;2. 南通科技职业学院,江苏 南通 226007)
水分是限制荒漠生态系统中植被生长、发育、演替的主要环境因子,并因此成为制约脆弱生态系统植被恢复和重建的短板[1]。区域降水量和土壤水分储量,决定了局地环境所能承受的植被种类、数量和质量的阈值,当降水来源不足或植物耗水增大时,供不应求的水分平衡关系将导致植被死亡和生态系统退化[2]。因此,研究植物与水分的供需特征,尤其是在水分匮乏的荒漠区,研究植物生长对干旱胁迫的响应机制,对于该地区生态系统的持续稳定性评价具有十分重要的意义。光合作用是植物生长的基础,是全球碳循环以及其他物质循环的基础和重要的环节[3]。同时,光合作用也是植物抗旱性能的主要指示指标之一[4]。光合进程不仅受植物本身生理特性的影响,同时受环境因素的制约。在光合作用中水是重要的参与因素,如果水分不足,会导致叶片气孔闭合,无法有效吸收二氧化碳,降低反应速率[5]。柠条(Caragana korshinskii)是我国干旱、半干旱荒漠区、黄土区广泛分布的优良乡土植物,具有广泛的适应性,在防风固沙、水土保持等生态效益方面发挥重要作用[6]。长期以来对恶劣环境的适应与演替,导致柠条在形态结构上具备了旱生特征,其优良的抗旱性、抗热性和耐盐碱性,以及对小气候和土壤质地、肥力的改良作用,使得柠条对退化生境具有很强的改善功能[7]。目前,对荒漠灌丛的研究多集中于生长生理特性[8]、林地水量平衡[9]等方面,研究灌木光合特性和水分利用特征的试验也已较多开展[10-11],但对柠条光合生理指标对土壤含水量变化响应的连续观测研究较少。因此,本研究以库布齐沙漠柠条灌丛为研究对象,人工控制土壤水分补给量和干旱程度,利用Li-6800便携式光合测定系统连续监测叶片光合指标及水分利用效率,探讨柠条灌丛对干旱胁迫的生态适应性,为干旱荒漠区柠条的人工栽培和管理提供科学依据。
1 材料与方法
1.1 试验区概况
研究区位于内蒙古自治区鄂尔多斯市准格尔旗布尔陶亥苏木(110°49′1″E,40°03′57″N,海拔1 100~1 300 m),该地区年平均气温在6.1~7.1 ℃,年平均降水量240~360 mm,多集中于7—9月,年平均日照时数3 138 h,年平均蒸发量2 560 mm,无霜期为130~140 d,年平均风速3.3 m/s。该地区土壤主要类型为风沙土,主要植物为柠条、沙柳(Salix psammophila)、杨柴(Hedysarum mongolicum)、油蒿(Artemisiaordosica)、狗尾草(Setaria viridis)、沙竹(Psammochloa villosa)、沙 蓬(Agriophyllum squarrosum)、雾冰藜(Bassia dasyphylla)、猪毛菜(Salsola collina)等。
1.2 试验方法
1.2.1 试验设置
试验于2018年5—8月进行。在试验区选择地势平坦、植物长势良好且形态指标基本均一的3年生柠条灌丛样地,样地基本情况见表1。样地中分别布设10块2 m × 2 m的柠条灌丛样方,并保证每个样方中至少有1株柠条生长,最终确定标准株柠条21株(株高:(90±15) cm,冠幅:(120±20) cm,基径:(2±0.5) cm,分枝数:8±3)。本试验中,利用土壤含水量降低的不同阶段来模拟不同程度的干旱胁迫,依据 GB/T 20481—2017[12]和参考文献[13],荒漠草原干旱程度可分为:土壤含水量12%~15%为轻度干旱,8%~12%为中度干旱,5%~8%为重度干旱,<5%为特重干旱。试验步骤为:1)减雨,在每个柠条样方设置遮雨棚,高度为冠层上方1.5 m 处,遮雨量约为降雨量的60%左右,并能够保证阳光的充足射入;2)隔水,在每个柠条样方四周挖取1 m深隔水槽,并铺设石棉瓦,以隔绝地表及深层土壤水分入渗补给;3)初始土壤水分设定,由于研究区降水稀少,土壤含水量在自然状态下很难达到轻度干旱水平,因此需人工补水。5月9日开始对各个样方进行灌水,为保证土壤水分均匀渗透及植物对水分的吸收适应过程,每5 d灌水1次,共计灌水4次,每个样地每次灌水300 L,共计灌水1 200 L,确保试验前1 m深土层土壤含水量稳定保持在>15%范围;4)土壤自然干旱过程,末次灌水后3 d开始测定植物光合特性及土壤含水量,在隔绝外界降水补给后,通过植物的蒸腾和土壤的蒸散发作用,在较长一段时间内土壤含水量会逐渐降至特重干旱水平。
1.2.2 环境因子测定
研究区空气温度、太阳辐射、光合有效辐射等气象资料由HOBO小型自动气象站记录(Onset,美国);使用TRIME PICO水分速测仪(IMKO,德国),分别测定各样方0~20、20~40、40~60、60~80、80~100 cm深土层土壤体积含水量。
1.2.3 光合参数测定
采用Li-6800便携式光合测定系统(Li-Cor,美国)测定光合参数,使用2 cm × 3 cm叶室,选择晴朗无云天气,于上午10:00—11:00对供试柠条标记叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)进行测定,每次测定重复5次,求得该指标平均值。叶片瞬时水分利用效率(WUE)按公式(1)[14]计算。每株柠条选取顶端健康生长的3片叶片进行光合测定。被测叶片面积使用LI-3000C便携式叶面积仪(Li-Cor,美国)进行测定,得出有效光照面积后,带入测量结果并计算各项光合生理指标。
1.3 数据处理与分析
试验数据处理及作图采用Excel 2007及Sigma Plot 14.0软件完成,采用SPSS 20.0软件进行统计检验,采用Pearson检验对水分利用效率与环境因子进行相关性分析,采用回归分析判别叶片光合特性与土壤含水量的关系,选取最小显著极差法(LSD)对干旱胁迫下的叶片光合特性指标进行差异性检验(α=0.05)。
2 结果与分析
2.1 柠条灌丛环境因子特征
如图1所示,柠条灌丛土壤含水量随干旱程度加深呈递减趋势,平均土壤含水量由16.15%降至3.76%。根据干旱程度分级标准,柠条灌丛土壤在6月3日至6月22日为轻度干旱,6月24至7月4日为中度干旱,7月11日至7月26日重度干旱,8月3至8月16日为特重干旱。柠条灌丛不同深度土层土壤含水量有显著性差异(P<0.05),总体表现为随土层加深,土壤含水量呈先增大后减小的变化趋势。表层0~20 cm土壤含水量最低,平均为8.37%,中间层40~60 cm土壤含水量达到峰值,平均为9.92%。试验期间,柠条灌丛空气温度变化范围是17.90~29.05 ℃,整体呈双峰曲线变化特征,分别在6月15日和7月16日达到温度顶点,在6月7日、6月22日和7月26日为温度低谷。柠条灌丛光合有效辐射呈动态的波动变化,最高值出现在6月7日,达到1 505.37 uE,最低值出现在7月22日,仅为303.70 uE。
2.2 干旱胁迫下柠条叶片光合特性的变化
如图2所示,试验期间柠条叶片光合特性指标均呈现出随监测时间的推移而逐渐降低的趋势,其中,Pn的变化范围是2.02~44.80 μmol/(m2·s),Tr的 变 化 范 围 是0.94~23.41 mmol/(m2·s),Gs的变化范围是0.02~1.09 mol/(m2·s),Ci的变化范围是238.12~323.05 μmol/mol。依据土壤干旱程度划分光合指标变化特征,不同土壤干旱程度下柠条叶片光合指标差异显著(P< 0.05),其中,Pn表现为轻度干旱(37.06 μmol/(m2·s))>中度干旱(28.23 μmol/(m2·s))>重度干旱(22.66 μmol/(m2·s))>特 重 干 旱(4.77 μmol/(m2·s));Tr表现为 轻 度 干 旱(18.08 mmol/(m2·s))>中 度 干 旱(9.98 mmol/(m2·s))>重度干旱(6.81 mmol/(m2·s))>特重干旱(2.21 mmol/(m2·s));Gs表现为轻度干旱(0.88 mol/(m2·s))>中度干旱(0.47 mol/(m2·s))>重度干旱(0.39 mol/(m2·s))>特重干旱(0.08 mol/(m2·s));Ci表现为轻度干旱(307.68 μmol/mol)>中度干旱(279.55 μmol/mol)>重度干旱(268.11 μmol/mol)>特重干旱(257.12 μmol/mol)。由上可知,土壤干旱程度可以显著影响柠条叶片的光合特性,在特重干旱下,Pn、Tr、Gs和Ci大幅度下降,植物的光合作用会受到严重制约。
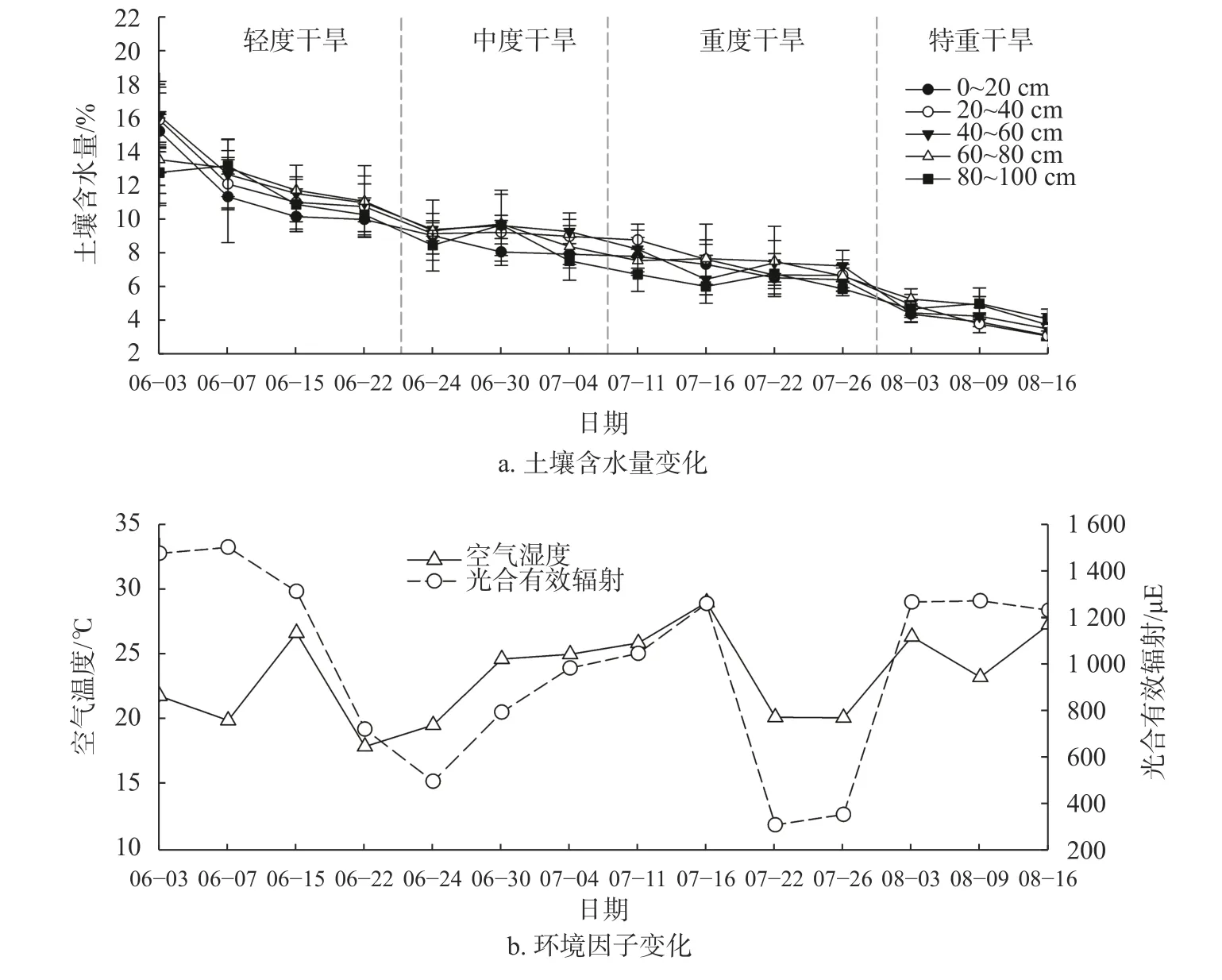
图 1 柠条灌丛环境因子动态变化特征Fig. 1 Dynamics of environment factors of C. korshinskii plantations

图 2 柠条叶片光合特性动态变化特征Fig. 2 Dynamics of physiological characteristics indexes of C. korshinskii leaves
2.3 柠条叶片光合特性对土壤水分的响应
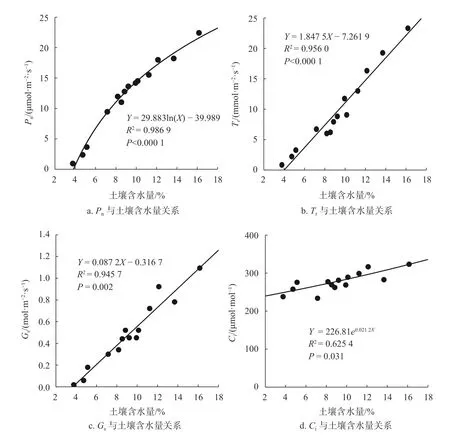
图 3 柠条叶片光合特性与土壤含水量的关系Fig. 3 Relationship between physiological indexes and soil water content of C. korshinskii leaves
通过建立回归方程,分别采用对数模型、指数模型和线性模型拟合土壤含水量对柠条叶片Pn、Tr、Gs和Ci的影响。如图3所示,当土壤含水量处于3.76%~16.15%范围内,Pn随土壤含水量增大而增大,二者的拟合关系符合对数函数,且具有极显著的相关性(P<0.01),土壤含水量可解释Pn变化的98.69%;Tr随土壤含水量的增大而增大,二者的拟合关系符合线性模型,且具有极显著的相关性(P<0.01),土壤含水量可解释Tr变化的95.60%;Gs随土壤含水量的增大而增大,二者的拟合关系符合线性模型,且具有极显著的相关性(P<0.01),土壤含水量可解释Gs变化的94.57%;Ci随土壤含水量的增大而增大,二者的拟合关系符合指数模型,且具有显著的相关性(P<0.05),土壤含水量可解释Ci变化的62.54%。
2.4 干旱胁迫下柠条叶片水分利用特征
如图4a所示,柠条叶片WUE的变化范围是1.88~3.92 μmol/mmol,在试验期间呈现单峰曲线变化趋势。依据土壤干旱程度划分WUE变化,不同土壤干旱程度下柠条叶片WUE差异显著(P<0.05),表现为重度干旱(3.35 μmol/mmol)>中 度 干 旱(2.87 μmol/mmol)>轻 度 干 旱(2.08 μmol/mmol)>特重干旱(2.15 μmol/mmol)。采用多项式回归模型拟合土壤含水量对柠条叶片WUE的影响,如图4b所示,WUE随土壤含水量的增大呈先增大后减小的趋势,当土壤含水量为7%~9%时,WUE达到最优化,在土壤含水量较为充盈或是极端干旱的条件下,WUE均处于较低水平。WUE和土壤含水量的拟合关系符合多项式函数,具有显著的相关性(P<0.05),土壤含水量可解释WUE变化的48.73%。除土壤水分外,一般来说,植物WUE还受到植物生理因子和环境因子等多方面的影响。通过Pearson相关性检验(表2),柠条叶片WUE与太阳总辐射和光合有效辐射呈极显著负相关关系(P<0.01),与空气温度和Tr呈显著负相关关系(P<0.05),而与Pn、Gs、Ci相关性较弱。
3 结论与讨论
土壤水分是植物生长过程的主要决定要素之一,会在不同程度上影响着植物的各种生理和生态过程,水分利用的有效性直接影响植物的光合作用和群落生产力[15]。尤其是在水分匮乏的荒漠生态系统中,有限的降水补给和巨大的蒸散消耗,导致该区土壤经常性处于干旱状态,而干旱胁迫严重影响植物的生长生理进程,是植物受到的一切非生物损害中,最严重的影响因子之一。在本研究中,通过人工控制土壤水分补给量和干旱程度,结果表明土壤干旱程度可以显著影响柠条叶片的光合特性,Pn、Tr、Gs和Ci均表现为轻度干旱>中度干旱>重度干旱>特重干旱,植物的光合作用在干旱胁迫下受到严重制约,这与罗彬莹等[14]对樟树(Cinnamomum camphora)幼苗及Yin等[16]对干旱区杨树(Populussp.)的研究结果一致。柠条叶片光合指标对干旱胁迫的响应,与植物自身的抗旱策略有关[17]。干旱越严重,气孔关闭的程度越大,导致Gs大幅下降,并造成Pn的明显受限。气孔关闭的同时能显著减少水分蒸腾,防止体内水分逸散[18]。因此,植物在干旱胁迫下的自我保护机制,是造成光合作用受限的主要原因[19-20]。
WUE能够反映植物耗水与其生产力之间的关系,荒漠生态系统中的植物以其独特的抗旱策略,在水分亏缺时能以高效的水分利用方式来抵御干旱胁迫[21]。在本研究中,WUE随土壤含水量的增大呈先增大后减小的趋势,当土壤含水量为7%~9%时,WUE达到最优化,而在土壤含水量相对较大或是极端干旱的条件下,WUE均处于较低水平,这与崔英等[22]对祁连园柏及杨司睿等[18]对罗布泊人工梭梭林的的研究结果一致。Jennifer等[23]研究也表明,土壤水分变化能够改变植物的水分利用特征,并且植物对水分变化的适应性与环境关系密切(如持续干旱或高于平均值的降雨)。这说明荒漠区柠条对水分的利用存在一个最适区间,适当干旱的土壤水分环境,能够促进其健康生长和正向演替。因此,在水分匮乏的荒漠区进行人工建植时,应当考虑对拟选植物种WUE的科学评价,以及对植物生长需水与环境水分承载力间动态平衡的考量,从而促进荒漠生态系统的正向演替和发展。
植物WUE既受自身因子调控,同时还受外部环境因子的影响。在本研究中,柠条叶片WUE与太阳总辐射、光合有效辐射、空气温度和Tr呈显著负相关关系,这与苏培玺等[24]对沙拐枣和泡泡刺等荒漠植物以及白灵娜等[25]对青海不同抗旱植物的研究结果一致。植物叶片光合作用、蒸腾作用及气孔变化规律对外部环境因子的敏感性,是叶片WUE差异性表现的基础,太阳辐射、温度等环境因子都是通过对植物的生理生化功能产生影响,进而间接改变了叶片的水分策略[26]。但是也有学者提出,植物WUE不仅受自身和环境因子控制,基因遗传特性也能够在很大程度上改变叶片水力行为[27]。然而,目前对于柠条光合和水分利用的相关调控基因研究仍比较缺乏,这是今后需要深入研究的科学问题。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
云南大学学报(自然科学版)(2022年4期)2022-08-03
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
少儿科学周刊·少年版(2021年20期)2021-01-17
农家科技(2020年3期)2020-05-11
当代陕西(2019年7期)2019-04-25
湖南饲料(2018年2期)2018-12-05
哈尔滨理工大学学报(2016年2期)2016-09-12
