
Assessment of the Effectiveness of Skull Morphometric Indices for Discrimination of Wild and Farmed American Mink
2021-04-28 01:49MOUXinQIYonglaiHUAYanYANGShuhuiPhilipSTOTTLIUZhenshengXUYanchun
野生动物学报 2021年2期
MOU Xin QI Yonglai HUA Yan YANG ShuhuiPhilip STOTT LIU Zhensheng XU Yanchun,5,6*
(1.College of Wildlife and Protected Area,Northeast Forestry University,Harbin,150040,China;2.Qinghai Qingqing Water Conservancy Technology Co.,LTD,Xining,810099,China;3.Guangdong Provincial Key Laboratory of Silviculture,Protection and Utilization,Guangdong Academy of Forestry,Guangzhou,510520,China;4.School of Animal and Veterinary Sciences,University of Adelaide,Adelaide,5005,Australia;5.National Forestry and Grassland Administration Detecting Center of Wildlife,Harbin,150040,China;6.National Forestry and Grassland Administration Research Center of Engineering Technology for Wildlife Conservation and Utilization,Harbin,150040,China)
Abstract:Captive rearing of wild animals is an important means of obtaining wild animal products.But products from poached wild animals can enter the market if fraudulently labeled as products of captive breeding.This poses a challenge to the protection of wild animals.Accurately identifying whether traded animal products are derived from the farm or the wild is key to protecting wild populations and maintaining the legality of the breeding industry.Taking the American mink(Neovison vison)as a model for small carnivores,we measured zygomatic arch space(ZAS),zygomatic breadth(ZAA and ZBA),basal length(BSL),mandible length(MDL),postorbital constriction(POC),greatest upper facial breadth(GFB),and other parameters to establish 9 skull morphometric indicators,namely POC/GFB,POC/BSL,POC/MDL,ZASSQRT/BSL,ZASSQRT/MDL,ZBA×BSL/2ZAS,ZBA×MDL/2ZAS,ZBB×BSL/2ZAS,and ZBB×MDL/2ZAS.The comparison between wild American mink(n=32,9 males and 23 females)and farmed American mink(n=45,35 males and 10 females)showed that all indices proved effective with moderate overall discrimination correctness for samples of known origin(73.2%-85.5%).The single exception was POC/GFB.When the sex was known,overall correctness was 82.4 to 93.8% for males,but 41.7 to 85.7% for females.Therefore,corresponding reference data sets should be established for animals of different genders and different sources,and the likelihood ratio should be used for identification.This result is not only valid for North American mink,but also provides reference for the identification of the sources of other small carnivores.
Key words: Forensic science;Origin discrimination;Skull morphometrics;Neovison vison;Small carnivore
Wildlife was one of the crucial resources during human evolution.Even in the present industrial era when wildlife resources have been mostly replaced with other resources,wildlife still plays important roles in human life as a carrier of different cultures[1].The still existing demand drives exploitation of wildlife resource.Exploitation together with habitat loss,climate change,and inadequate management has resulted in decline and even endangerment of species worldwide[2].Therefore,species conservation is an important mission and action in the present era.
Banning or restricting exploitation is an important strategy for wildlife conservation.Meantime,farming of wildlife is an alternative approach to reduce exploitation pressure placed on wild population through providing products from farms[3].However,wildlife farming leads to a conundrum in that illegally obtained wild products may enter the market in the guise of legal farm products.To enable effective control of trade in products derived from free-living populations of wild animal species that are also farmed,law enforcement officers need tools that can discriminate between products derived from these two sources.
The American mink(Neovisonvison)is farmed extensively in North America,northern Europe and China.Escapees and individuals deliberately released for hunting have established free-living feral populations in farming countries and are influencing local biodiversity[4-6].Where the mink is an exotic species the removal of feral animals is greatly encouraged in order to reduce adverse impacts[7-8]and there are no legal impediments to their harvesting allowing the development of an industry based on the wild population.In such a situation,both captive-reared and wild-reared animals may be readily available.The American mink is representative of all small carnivores with economic value as fur-bearers,and therefore it is an ideal model animal to develop techniques to separate farmed animals from their wild counterparts.
The skull is the feature that offers the most potential for discrimination between animals based on their source.The structure of skull is very complex and its morphology is sensitive to environment changes[9-10].Morphometric approaches can mine information that demonstrates the animals’ response to different environment factors between farm and wild with low time and financial costs.
In mustelids,many classic cranial indices have been applied to morphometric analysis of sexual dimorphism[11-12];between-species and-subspecies morphological variation[13-16];static,growth phase or geographic variations[17-18];and environmental influences on cranial development[19-20].The advantage of classic indices is that they are measured using common methods enabling comparisons of datasets created in different studies.However,biological meanings of these indices are not well described in relation to functions.Consequently,morphological differences cannot be interpreted in relation to adaptations to different environments,even though principal components analyses have identified some indices making major contributions to variations.Designation of methods for discrimination between wild and farmed conspecific individuals relies on physiological response or adaptation mode of animals to different factors in farm or wild environments and consequent morphological alterations in skeletal systems.Three dimensional(3D)morphometric technologies provide alternative approaches using arbitrary landmarks to access morphological characteristics that are poorly or not able to be described using classic measurements[21-23].The combination of 3D tools and functional morphometric analysis would facilitate the exploration of novel indices for discrimination between wild and farmed conspecific individuals.
It has been discovered that farmed minks have a larger skull,a narrower postorbital constriction,and a shortened palate[9,24].Using classic measurements,the condylobasal length(CBL)and the postorbital constriction(POC)were able to correctly identify 100% males and 90% females[24].However,relative indices(ratios)rather than raw measurements are used in most instances for comparisons of shape and size in skull morphometric analysis because of size-adjustment functions[25].Choice of reference is essential for making up relative indices with functional meanings.For example,in cats the cubic root of the product of greatest skull length,bizygomatic breadth and mandible length was regarded as a gold reference to adjust for different skull sizes[26].However,the skull is not always complete and so the suite of measurements required for some indices used in taxonomic studies may not be available.Hence,in forensic practice,it is necessary to develop alternative relative indices based on multiple skull measurements.
We previously observed that the temporalis and masseter muscles and the sagittal crest and nuchal crests are all better developed in farmed minks than in wild minks.The temporalis muscle starts from the sagittal crest,passes through the inner space of the zygomatic arch and ends at the coronoid process of the mandible.The temporalis muscle pulls on the coronoid process during contraction and pivots the mandible upward around the axis of the mandibular joint to achieve biting or chewing[27].The contraction force of a skeletal muscle is related to its cross-sectional area[28]and so it can be expected that a large space within the zygomatic arch is required to hold a well developed temporalis muscle.Therefore,it can be predicted that the zygomatic arch space(ZAS)would be greater in farmed minks than in wild ones.If differences were significant,novel indices could then be set to separate farmed from wild animals up based onZAS.
In addition,the mandible is morphologically and functionally associated with the maxilla which contributes to the overall length of the skull.Thus the mandibular length(MDL)should be conservative in relation to skull length.If so,MDLwould be an alternative reference for setting up indices if measurements of skull length such asCBLor basal length(BSL)cannot be made due to incompleteness of the skull.
In the present study,we tested the scaled relationships of skull measurements namely zygomatic breadth(ZB),mandibular length(MDL),postorbital constriction(POC),greatest upper facial breadth(GFB)and zygomatic arch space(ZAS)with the basal length(BSL)on the farmed and wild American minks,and assessed the effectiveness of six indices based on these measurements for origin discrimination.
1 Materials and Methods
Wild American minks(9 males and 23 females)were trapped during winter in Greater Khingan Mountains of China in 2011 and 2012.Farmed American minks(35 males and 10 females)were obtained from a fur farm in the same region.The head of each mink was isolated from the carcass.Muscles were roughly removed from the skull using a scalpel,then the skulls were cooked in boiling water until the retaining muscle and tendons were soft enough to remove.The cleaned skulls were then briefly boiled in water containing-0.1% Na2CO3to remove fat,and then dried naturally.
The skulls were examined qualitatively,particularly on the shape of sagittal nuchal crests,postorbital constrictions,parietal surface and cranial cavity shape.For quantitative examination,each skull was fixed on the support frame of a 3 dimensional digitizer(MicroScribe-G2LX,Immersion Corporation,US).Orthogonal coordinates of 5 landmarks were measured:A,the tip of the frontal process of the zygomatic bone;B,zygion;C,the most caudal point of the fossa in which the zygomatic process of the temporal bone reflects onto the facies temporalis;D,the point at which the sphenoid,frontal,and parietal bones touch;and E,the tip of the ectorbital process(Figure 1).The area of the 3D pentagon ABCDE was regarded as the zygomatic arch space(ZAS),and calculated by dividing it into three trianglesΔABC,ΔACEandΔDCE.The each side length of each triangle was calculated using the equation:

whereDABis the side length betweenAandB,xA,yA,zA,xB,yBandzB.The area of each triangle was calculated using Heron’s formula based on side lengths calculated above:
SΔABC=[p(p-DAB)(p-DAC)(p-DBC)]1/2
whereSΔABCis the area of the triangleΔABC,P=(DAB+DAC+DBC)/2.The area of the pentagon ABCDE(ZAS)was the sum of the three triangles.
Zygomatic breadth at landmark A(ZBA)and B(ZBB),basal length(BSL),mandibular length(MDL),postorbital constriction(POC),and greatest upper facial breadth(GFB)were directly measured using an electrical vernier caliper to the nearest 0.1 mm according to the standard method[29].
Independent samples tests were used to compare means ofZBA,ZBB,BSL,MDL,POC,GFB,ZAS,POC/GFB,POC/BSLorPOC/MDL,ZASSQRT/BSL,BSL×ZBA/2ZASandBSL×ZBB/2ZASbetween farmed and wild group and between sex groups with significance criteria<0.05.Linear regression was used to analyze scaling relationship between LogBSLand logarithmic transformedMDL,POC,GFB,ZBA,ZBBandZASSQRTin farm and wild group.In order to assess the effectiveness of relative indicesPOC/GFB,POC/BSL,POC/MDL,ZASSQRT/BSL,ZASSQRT/MDL,ZBA×BSL/2ZAS,ZBA×MDL/2ZAS,ZBB×BSL/2ZAS,andZBB×MDL/2ZASfor discrimination of origin,Fisher’s discrimination was used to test the percent correct classification of each group.All statistical analyses were performed using software SPSS 19.0(SPSS Inc.,Chicago,IL,USA).
2 Results
2.1 Differences and sexual dimorphism of indices between farm and wild group
The skull of the farm group fell into two types.Type Ⅰ including 35 males and 4 females exhibited well-developed sagittal and nuchal crests,narrow postorbital constrictions,rough parietal surfaces and narrow foreparts of the cranial cavities that contains the olfactory bulbs.Type Ⅱ including 6 female samples exhibited less developed sagittal and nuchal crests,broad postorbital constrictions,smooth parietal surfaces and plump foreparts of their cranial cavities.The wild group,regardless of sex,resembled Type Ⅱ farmed females in all mentioned features,but the sagittal and nuchal crests were even less well developed(Figure 2).The Type Ⅰ skull is limited to farmed minks,but Type Ⅱ seemed to be a transitional form between the wild group and the farm group.
As shown in Table 1,BSLwas greater in farm group than in wild group(t=8.984,P<0.001).It also exhibited significant sexual difference between the two groups(t=14.101 for farm group andt=5.965 for wild group,P<0.001 for both).A similar tendency was observed forMDLin that it was greater in farm group than in wild group(t=9.198,P<0.001)and in males it was greater than in females(t=14.478 for farm group andt=5.732 for wild group,P<0.001 for both).

Tab.1 Comparisons of skull measurements between farmed and wild American minks
GFBvaried between 15.7 mm and 22.1 mm for farm group,and between 13.1 mm and 18.3 mm for wild group.Significant mean differences were detected between wild and farm group(t=8.483,P<0.001)and between sexes(t=2.482-8.425,P<0.023).In contrast,POCranged from 10.4 mm to 15.6 mm for farm group,and from 10.4 to 13.2 for wild group.No differences between sexes were detected within farm group(t=1.691,P=0.100),but not within wild group(t=2.608,P=0.016).A significant difference was detected between the farm group and wild group(t=2.055,P=0.044).The ratioPOC/GFBshowed a significant difference between sexes within farm group(t=4.084,P<0.001),but not within wild group(t=0.193,P=0.849).It also differed between farm group and wild group(t=6.686,P<0.001).
The zygomatic breadths at landmark A(ZBA)and B(ZBB)were both significantly greater in farm group than in wild group(tA=7.259,PA=0.000;tB=5.991,PB=0.000).BothZBAandZBBexhibited significant sexual dimorphism:greater in males than in females(tZBA=9.451,PZBA=0.000;tZBB=11.505,PZBB=0.000 for farm group andtZBA=2.958,PZBA=0.008;tZBB=2.075,PZBB=0.052 for wild group).
TakingBSLas a reference,the relative zygomatic breadths at A(ZBA/BSL)and B(ZBB/BSL)were calculated,but no significant differences were detected between either the farm and wild group(tZBA=1.733,PZBA=0.145;tZBB=0.086,PZBB=0.932)or between sexes(t=0.416-1.986,P=0.055-0.682).Mean zygomatic arch space(ZAS)was 331.6 mm2(range 190.9-526.3)for all minks.The farm group had greaterZASthan wild group(t=8.121,P<0.001).ZASalso revealed significant sexual dimorphism:it was greater in males than in females(P<0.001).
2.2 Scaling of indices
Taking LogBSLas a reference,differences of scaling relationships between farm and wild group were compared(Figure 3).For the linear model Log(y)=Log(b)+α(Log(x))[30],xisBSL,andyisMDL,POC,GFB,ZBA,ZBBandZASSQRT,respectively.Results showed the allometric coefficientαMDLof the regression between LogMDLand LogBSLwas slightly higher in the wild group(α=1.156)than in the farm group(α=1.191),but the intercepts Log(bMDL)were very close(-0.550 for farm group,-0.490 for wild group),suggestingMDLgrows at almost the same speed and retains the same proportion in the wild and farmed animals.
LogPOCvaried greatly between the two groups.There was a slight but significant relationship with LogBSLfor the wild group(R2=0.239,P=0.013),but no relationship for the farm group(R2=0.024,P=0.364).In contrast,LogGFBwas less variable and showed significant regression with LogBSL(R2=0.677,P<0.000 for farm group,R2=0.352,P=0.005 for wild group).The allometric coefficientαGFBwas 1.092 in farm group and 0.932 in wild group.The intercept Log(bGFB)was -0.729 for farm group,and-0.443 for wild group.
LogZBArevealed significant regression with LogBSLin both groups(R2=0.795 for farm group and 0.827 for wild group,P<0.001 for both).The coefficientαZBAwas very close(0.986 for farm group and 1.033 for wild group),but a considerable difference between the intercepts was observed;it was higher in farm group(-0.251 for farm group and -0.347 for wild group).LogZBBshowed the same significant regression trends(R2=0.910 for farm group and 0.817 for wild group,P<0.001 for both)as LogZBA,but great different ofαZBBwas detected between farm group(1.232)and wild group(0.994).
LogZASSQRTexhibited similar trends as the LogZBBin regression significance(R2=0.947 for farm group and 0.891 for wild group,P<0.001 for both)and difference in coefficientαZASSQRT(1.621 for farm group and 1.251 for wild group).Figure 3 also demonstrates the partition of the logarithmic data of all measurements between farm and wild group.
2.3 Indices and their effectiveness for discrimination of origin
Based on the scaling analysis above,relative indices were set up using measurements with different intercepts(Log(b))and/or allometric coefficient(α)between farm and wild group.The indices includePOC/GFB,POC/BSL,POC/MDL,ZASSQRT/BSL,ZASSQRT/MDL,ZBA×BSL/2ZAS,ZBA×MDL/2ZAS,ZBB×BSL/2ZAS,andZBB×MDL/2ZAS.Their biological meanings are shown in Table 2.

Tab.2 Biological meanings of skull morphometric indices for discrimination of farm and wild American minks
All indices were significantly different between farm group and wild group(P<0.001).The values of all indices were broader in ranges in farm group than in wild group,and considerably overlapped(Figure 4).The effectiveness for origin discrimination varied from 78.2% to 85.5% whenBSLwas the reference used to set up the index.WhenBSLwas replaced withMDL,the effectiveness declined by 1.60%-8.60% for all indices exceptZBB×BSL/2ZASwhose correctness increased from 78.2% to 81.8%.
It was apparent that the correctness for farm group and wild group differed.The greatest difference was seen onPOC/GFBfor which the correctness was 83.8% for farm group,but became as low as 68.0% for wild group.ZBB×MDL/2ZASalso demonstrated great difference of effectiveness between farm(76.5%)and wild group(90.5%).The most balanced effectiveness was seen onPOC/BSLandZBA×MDL/2ZASof which the differences were 2.50% and 2.70% respectively.IfPOC/GFBandZBA×MDL/2ZASwere combined,the overall correctness of origin classification increased to 83.8%,and meantime,the correctness was compromised to 79.4% for farm group and 90.0% for wild group.
All the measurements were sexually dimorphic(Table 1).The effectiveness of the indices above was also influenced by sex.When sexes were known,our data demonstrated the overall correctness of origin classification ranged from 82.4% to 93.8% for males,but from 41.7% to 85.7% for females.OnlyPOC/BSLandPOC/MDLshowed balanced effectiveness for males and females(82.4% vs 85.7% forPOC/BSLand 88.2% vs 85.7% forPOC/MDL)(Table 3).
Considering the similarity of Type Ⅱ farm skulls to wild specimen,we separately tested the effectiveness of all nine indices for Type Ⅰ and Type Ⅱ farm skulls.When Type Ⅱ skulls were excluded from the farm group,the overall correctness was between 82.0%-89.3%;while the correctness declined to 55.6%-67.7% if Type Ⅰ skulls were excluded(Table 3).
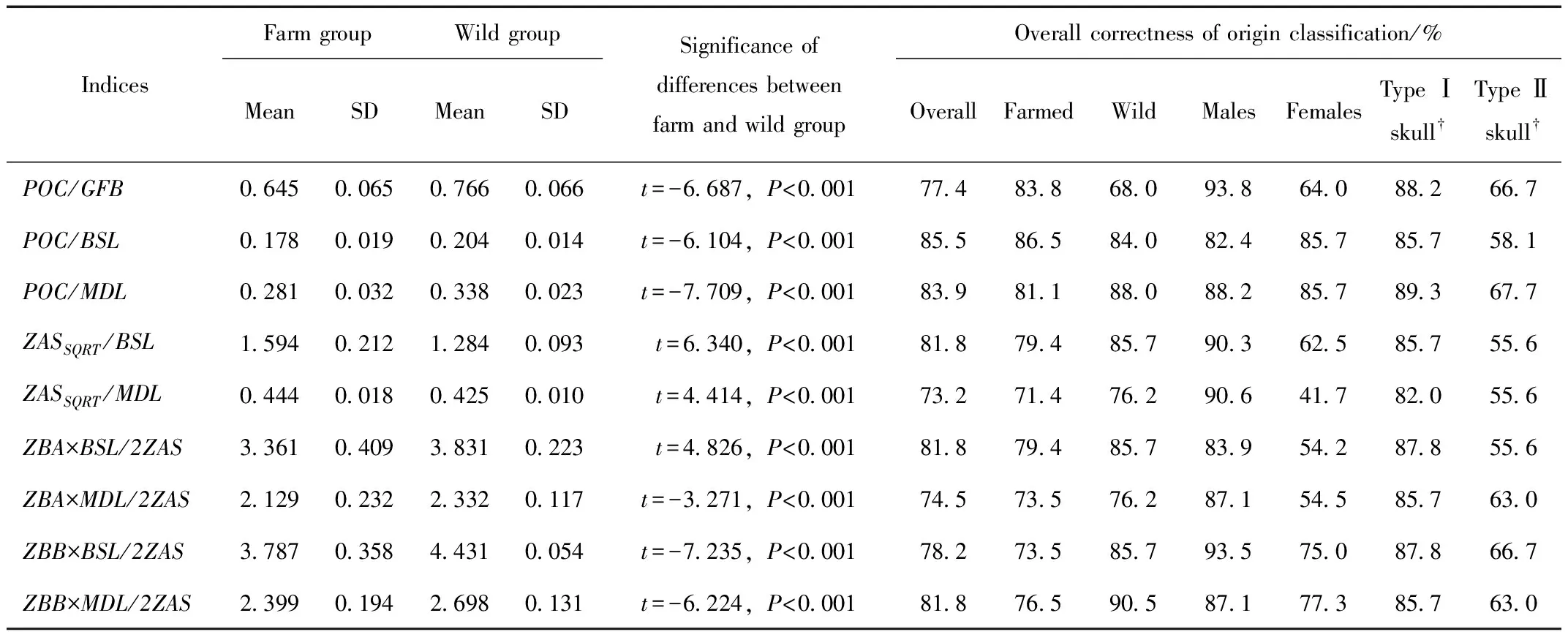
Tab.3 Effectiveness of skull morphometric indices for discrimination of farm and wild American minks
3 Discussion
3.1 Morphometric changes of mink skull in farms
Breeding in a farm applies selective environmental pressures to the animals that differ from those applying in the wild.For example,farmed animals have to adapt to different components,availability,texture and nutritional quality of foods,different movement patterns and different stress factors.The shift of those factors results in morphological changes in many aspects[31].For instances,caged wild animals are characterized by shortened ulnas and tibias[19-20],shortened muzzles[5,31],reduced brain sizes and reduced sizes of components of the brain[5]and narrowed postorbital constrictions[24],comparing to wild individuals.
Those morphological changes are representations of phenotypic plasticity and evolutionary adaptation[32].Phenotypic plasticity is the ability of a single genotype to produce multiple phenotypes in response to variation in the environment[33].Muscles and the skeleton interact with each other during growth.Locomotor activity type and level influence the architecture,potential excursion and force production of muscle and also the growth rate of bone[34].The captive environment,particularly the small,simple cages and the rigid lifestyle,may induce stereotypic behavior and thus influence ontogenic development.For instance,stereotypic behavior in the form of heightened rotation of the head and neck in the lateral plane leads to malformed external occipital protuberance,and reduced jaw activity leads to a flattening of the sagittal crest in captive tigers[35].There have also been many stereotypic behaviors identified in farmed minks,such as repetitive head/upper body movements,locomotive route-tracing,subject scrabbling or scratching[36],as well as biting the cage,chewing nest materials and even hair[37].Such farm-unique behaviors would result in morphological changes ontogenically.Frequent and intensive use of the muscles of mastication and neck muscles that attached to the head by farmed minks would stimulate the development of the sagittal and nuchal crests,the zygomatic arch and the mandible.
Our data support this expectation.Both the sagittal and nuchal crests were observed to be well developed in typical farmed minks(Figure 1).Zygomatic arch related measurements,ZBA,ZBBandZAS,were all significantly greater in farmed animals.Meanwhile,scaling with LogBSLshowed LogZBAhad similar rates of changes(α)between farm and wild group,but a high intercept(Logb)in farm group;LogZBBand LogZASSQRThad high rates of change and low intercepts in farm group(Figure 4).However,mandible length(MDL)violated our expectation and a previous report that farm breeding may lead to a shortened and broadened snout[5].The scaling with LogBSLshowed similar rate of change and intercept with LogMDL(Figure 3).Type Ⅱ skulls of farmed minks were similar to wild samples,but possessed better developed crests.This might be the result from less stereotypic or other farm-unique behaviors[38]than by those individuals with typical farm skulls.
From the adaptation aspect,commercial farming of wildlife often favors productive performance,such as large body(pelt)size,growth rate,litter size etc.towards which artificial selections are applied[39],and even non-commercial captive breeding for reintroduction also inevitably makes changes in morphology and reproductive rates due to relaxed selection for traits required for survival in the wild[40].For instance,selection of silver foxes(a melanistic form of red fox,Vulpesvulpes)for tameness in a 40-year experiment led to remarkable transformations including reduced cranial height and width,and a shortened and broadened snout in addition to the tame behaviors[41].Like the silver fox,the farmed American mink is also selected for tameness,body size and coat color[32,42-45]and also shows changes in skull shape[9].The present study showed that farmed minks exhibited narrowed postorbital constriction(POC/GFB)and less plump front cranial cavity(containing olfactory bulb)compared to wild minks(Figure 2,Table 1 and 3).This support the finding in Kruska et al.[5],implying that wild minks rely more on olfactory sensation than farm minks and have better developed olfactory bulbs,which is reflected in the shape of the skull in the postorbital zygomatic space.The modifications of skull morphology specific to farmed mink might be a side outcome of selection.It could be expected that the longer the selection pressure was applied,the more stable the farm-specific morphology would be.
The effects of phenotypic plasticity and adaptation cannot be separated rigidly since the two mechanisms always work in association on traits[46].Nevertheless,the changes provide opportunities to set up indices for discrimination between farmed and wild animals.This prediction seems to have universality to all wildlife species whose behavior significantly alters in a farm environment,and/or to which artificial selection is applied.
3.2 Effectiveness of indices for origin discrimination
Since farming leads to changes in skull morphology,differentiation of farmed from wild animals becomes possible based on morphometric analysis.The present study set up nine indices for origin discrimination using the farmed and feral American minks(Table 2,3 and Figure 4).Table 3 showed all nine indices were significantly different between farm and wild group(P<0.001 for all).The overall correctness of origin was moderate,varying between 73.2% and 85.5%.
The effectiveness of these indices is influenced by two factors.The first and the more essential factor is the extent of substantial changes in skull morphology when animals grow in a farm environment.We observed two types of skull in the farm group.Type Ⅰ was essentially different from skulls in the wild group.However,Type Ⅱ skulls were quite similar to wild male specimens.The reduced difference between farm and wild skulls diminished the effectiveness of indices.Statistical analyses in general indicated significant but moderate differences(Table 1).This demonstrates that the effectiveness of indices based on these measurements would be moderate at best.
The second influencing factor is the sexual dimorphism.Observations in the ferret(Mustelaputoriusfuro)demonstrated that craniofacial growth follows the same pattern for both sexes in the ferret,but slows down and finally ceases earlier in the female than in male.Meanwhile,the skull of the male ferret is not simply a larger version of the female one:the males has a proportionally broader face than the females[23].In the present study of American minks,20 of 25 cranial characters were significantly higher in males and the degree of dimorphism even varied geographically[12].This suggests that sexual dimorphism is environment-dependent.There was an expectation that sexual dimorphism would be reduced in farmed minks because resource-driven evolutionary pressure maintaining dimorphism would have become relaxed[9].However,in our study significant dimorphism was still detected for all seven measurements in both the farm and wild groups(P=0.001-0.023,Table 1),counteracting the effectiveness of those indices for origin discrimination.
The scaling plots(Figure 3)reveal a general partition of values of the six measurements between the farm and wild groups.However,there was no significant difference between farmed females and wild males(P=0.262-0.975).Overlapping of value ranges would lead to ambiguous judgment of farmed females and wild males and lower the effectiveness of indices for origin discrimination.Figure 4 showedPOC/BSLandPOC/MDLare the least overlapped between farm and wild group than the rest of the indices.Their effectiveness was apparently higher than other indices consequently(Table 3).When sex was taken into account,the correctness of origin discrimination of all indices butPOC/BSLcould be significantly improved achieving 82.4%-93.8% for males.However,the correctness of all indices butPOC/BSLandPOC/MDLdeclined by 3.20%-31.50% for females(41.7%-85.7%,Table 3).This is mainly because of the influence of Type Ⅱ skulls in farm group.If Type Ⅱ skulls were excluded from the farm group,the correctness was generally improved.In contrast,the correctness declined to below 70% if Type Ⅰ skulls were excluded.It suggests the indices are effective for Type Ⅰ skulls,but not so for Type Ⅱ skulls.
3.3 Application concerns
The nine indices tested are generally effective tools to discriminate the origin of mink skulls.They are also potentially applicable to other small carnivores.However,the effectiveness was considerably restricted by the great morphometric variation in the farm specimens and was particularly poor for separating Type Ⅱ farm skulls from wild skulls.We recommend setting up three separate databases for wild,Type Ⅰ and Type Ⅱ farm animals using the indices.Each database should contain large numbers of specimens.Likelihood ratios could be calculated from indices for skulls of known origin using the database,then a skull presented as evidence could be assigned to a category by comparing the likelihood ratios.However,before morphometric examination of the evidence was undertaken,skulls should be primary assigned according to the qualitative characteristics of typical farm skulls,including well developed sagittal and nuchal crests,narrow postorbital contractions and rough parietal surfaces,which would help to improve the reliability of the conclusions.When the likelihood values of the evidence being assigned to Type Ⅱ or the wild category were about equal,additional information would be needed.
4 Acknowledgements
The authors are grateful to Dr.MA Yue,Dr.WANG Zhen,Miss ZHOU Xiaoli at Northeast Forestry University,China for their assistance of sample treatments.We also thank Dr.Declan Morris at the University of Adelaide,Australia for his help in data treatment.
