
含ACC 脱氨酶的根际细菌提高植物抗盐性的研究进展
2021-04-26 01:23王琪媛王甲辰叶磊姜帆
生物技术通报 2021年2期
王琪媛 王甲辰 叶磊 姜帆
(1.北京师范大学生命科学学院,北京 100875;2.北京农林科学院植物营养与资源研究所,北京 100089;3.国网(北京)综合能源规划设计研究院有限公司,北京 100082)
土壤盐渍化严重危害农业生产。在近十几年中,导致土壤盐渍化的主要原因是用含有盐分的水进行灌溉,随着水分的蒸发作用,大量盐分积累进而导致土壤盐化。据统计,全球范围内共逾1.2×108hm2(约为陆地总面积的10%)的土地是盐土,可耕种土壤中约50%以上的面积受盐害威胁[1-2]。盐害问题的严重性还随着农业可耕地移作他用(如农村城镇化)而愈演愈烈。据估算,按现在的速度发展,至2050 年农业生产将不足以维持全球93 亿人口所需[3]。因此,可耕地的盐碱化是全球性的重大资源和环境问题。我国盐碱地面积约3.5×107hm2,主要分布在17 个省份。虽然我国是盐碱地大国,但是仍有约1.3×107hm2的盐渍化荒地和盐碱障碍耕地具有潜在的农业利用价值[3-4]。因此帮助植物在盐渍土中生长具有重要的意义。
很多科研工作者利用转基因技术或传统的杂交手段筛选培育耐盐作物。迄今为止已有的实验数据表明,这些手段虽然已经取得了一些成功,但是对这些植物遗传稳定性的长效测试是一个费时费力的过程[5]。另一种策略是利用对植物有益的菌群,即植物生长促进菌群(Plant growth promoting bacterium,PGPB)[6]。PGPB 主要存在于植物根系周围的土壤中,即被称为根际的微生态系统中,也存在于植物组织中以及叶片或茎的表面。在这一大类菌群中,根际促生细菌的种类繁多,并且通过多种机制促进植物生长,提高植物的抗逆性。含ACC 脱氨酶的根际细菌在众多细菌中格外引人注目,因为它在提高植物对多种非生物胁迫的抗逆性方面发挥着显著作用,尤其在提高植物的抗盐性方面[7-8]。
本文回顾了近十几年已发表的关于含ACC 脱氨酶的根际细菌提高植物耐盐性的研究成果,并总结这类根际细菌提高植物耐盐性的生理和分子机制。明确含ACC 脱氨酶的根际细菌在盐碱化农业生态系统中潜在的应用价值。
1 根际促生细菌的概念
植物遭受干旱、盐碱等胁迫时往往由植物根系率先感知并作出应答。一般把距离根轴表面数毫米范围内的土壤空间称为植物根际,植物根际土壤中微生物的多样性远远超过没有植物定植的土壤[9-10]。定植于植物根际土壤空间,并且能够通过植物根系与植物发生直接或间接的物质与能量交流的土壤微生物称为植物根际微生物。在从植物根系获取相应养分的同时,根际有益微生物能够通过形式多样的作用机制促进植物的生长发育,进而实现互利互惠的作用效果。目前研究较多的根际有益微生物是根际真菌和根际细菌两大类。
植物根际促生细菌(Plant growth-promoting rhizobacteria,PGPR)指定植于植物根系或根际土壤中的能够通过多种机制促进植物生长发育的有益细菌。它们在改善植物根际土壤环境,促进植物生长,提高植物在生物胁迫和重金属毒害、干旱、盐碱等非生物胁迫下的抗逆能力等方面发挥着重要作用。根际促生细菌通过提高植物的营养状况或者改变植物体内的激素水平来促进植物生长,提高植物对逆境的抵御能力。这其中含有ACC 脱氨酶的根际细菌在提高植物抗逆性方面所发挥的重要作用引起了科研工作者的广泛关注[7-8]。
2 含ACC 脱氨酶的植物根际促生细菌的作用机理
有些根际促生细菌含有ACC 脱氨酶,它可以将ACC(乙烯合成的直接前体)分解为氨和α-酮丁酸,这两种产物可以分别作为细菌生长的氮源和碳源[11]。该酶已经得以纯化,并且在1991 年由Sheehy 等[12]测定了该酶基因的全部序列。此后,ACC 脱氨酶的活性在许多其他的微生物(主要是植物根际促生细菌)中被测定,部分植物根际促生细菌的ACC 脱氨酶基因也被测序。比如假单孢菌UW4 的ACC 脱氨酶蛋白以及它的调控水平都已经被广泛研究[13-14]。绝大多数的植物根际促生细菌体内的ACC 脱氨酶的Km 值相对较小(约为3.4 mmol/L),最适pH 约为8,由3 个亚基(各约36.8 kD)组成,而且每个亚基都需要一个磷酸吡哆醛作为辅因子[13-14]。
含有ACC 脱氨酶的植物根际促生细菌生活在土壤中,并且对植物的根际分泌物(如有机酸、氨基酸、核苷酸和维生素等)具有趋化性,从而吸附于植物根系的表面(图1)。ACC(1-aminocyclopropane-1-carboxylate)是乙烯生物合成的直接前体,当植物持续遭受干旱、盐碱等外界环境胁迫时,ACC 合成酶的含量和活性上升,植物细胞内就会产生更多的ACC,而部分ACC 可以从根系分泌到根际,这部分ACC 可以被细菌吸收进而被ACC 脱氨酶分解。由于根外部的ACC 不断的被分解,浓度不断地下降,植物细胞内的ACC 就会顺着浓度梯度源源不断地向根外分泌。通过这种方式,细菌成为ACC 库,从而降低植物体内能被ACC 氧化酶催化生成乙烯的ACC 总量,进而降低胁迫条件下植物体内的乙烯含量[15-16]。
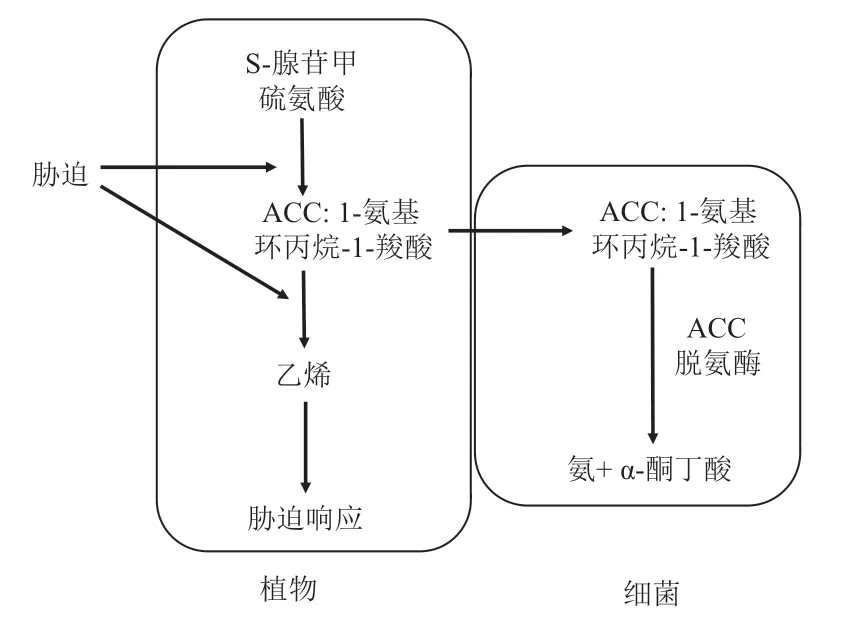
图1 细菌体内的ACC 脱氨酶降解植物产生的ACC 示意图[16]
3 乙烯的生理功能及其生物合成
乙烯调控着植物的许多生命过程,包括种子萌发、根系发育、开花结果、衰老和脱落[17-18]。它在细胞伸长、细胞死亡、生物和非生物胁迫应答、结瘤和对病原菌的系统性防御过程中也发挥着重要作用[19]。乙烯最显著的生理效应之一是“三重反应”[20]。在有些植物中,乙烯参与打破种子休眠和促进种子萌发的过程[21]。在种子萌发过程中,乙烯促进顶端弯钩的形成,这对于下胚轴钻出地面而言是必需的。乙烯能够与生长素和独角金内酯共同作用并促进拟南芥根毛的生长[22]。在淹水条件下,乙烯能够与生长素相互作用,调控植物不定根的发生与生长[23-24]。此外,乙烯还能够促进果实的成熟,诱导凤梨开花。乙烯对植物的生长也有抑制作用。有实验数据显示,在高浓度的硝酸盐胁迫下拟南芥侧根中乙烯的含量上升,侧根的生长受到抑制[25]。乙烯还通过抑制表皮细胞的生长进而抑制拟南芥植株的生长[26]。乙烯能够抑制由ABA 所诱导的气孔关闭[27-28]。
许多生物和非生物胁迫都会引起乙烯的大量合成[19]。乙烯生成量的激增在下面几种胁迫条件下都有发生:盐胁迫[15]、氧化胁迫[29]、高温胁迫和干旱胁迫[30-33]、冻害、水涝、机械压力[34]、重金属胁迫[35-38]和病原体感染[39-41]。
在高等植物体内,乙烯的合成前体是S-腺苷甲硫氨酸,它在ACC 合成酶的催化下转化为ACC(1-氨基环丙烷-1-羧酸),之后在ACC 氧化酶的催化下,转变为乙烯。在乙烯的生物合成途径中,ACC 是乙烯生物合成的直接前体物质,同时也是乙烯生物合成的限速步骤,ACC 含量的降低将直接导致乙烯含量的降低。
非生物胁迫条件下(如盐害)测定乙烯的含量,结果显示有两个明显的乙烯生成阶段[42]。第一个阶段是胁迫开始后不久的小高峰,该阶段比较短暂,被认为是保护性防御响应的开启过程;第二个阶段是胁迫开始后的数小时或数天出现的大高峰,乙烯的急剧累积会诱导植物叶片衰老脱落等一系列反应,抑制植物的生长发育,降低了植物体的抗逆性,因此被称为胁迫乙烯(Stress ethylene)。即较低浓度的乙烯参与植物抵御非生物胁迫的应答过程,而当乙烯的产生量超过一定的阈值后就会转变为抑制植物生长发育的胁迫乙烯(图2),降低植株对环境胁迫的抵御能力[7,9,42]。

图2 环境胁迫下植物激素乙烯含量随时间的变化[42]
当胁迫刚开始时,植物组织内的ACC 含量较低,ACC 氧化酶将ACC 转化为乙烯,生成的乙烯通过上调ACC 合成酶和ACC 氧化酶基因的表达催化自身的级联放大反应,最终生成大量的乙烯[43-44]。ACC合成酶的基因表达还受环境因素的调控,胁迫条件下ACC 合成酶的活性会升高[45]。
降解SAM(S-腺苷甲硫氨酸)或ACC 的酶以及乙烯合成抑制剂[如氨基乙氧基乙烯基甘氨酸(AVG)]都可以降低胁迫条件下乙烯的含量,从而维持植物正常的生理过程[46-47]。此外,降低乙烯的感知能力也能提高植物的胁迫耐受性。然而,乙烯合成的化学抑制物(如AVG)不仅昂贵而且会给环境带来有害的影响[11],因此含有ACC 脱氨酶的根际促生细菌就成为了环境友好型的选择。
4 盐胁迫对植物的毒害作用
土壤盐渍化的主要特征是土壤溶液中含有高浓度的Na+、Mg2+、Ca2+(阳离子)和CO32-、HCO3-、Cl-、SO42-(阴离子),其中以Na+和Cl-浓度最高[48]。当土壤溶液的电容达到4 dS/m 及以上时,土壤就成为盐渍化土壤。当土壤溶液中NaCl 的浓度达到40 mmol/L 时,其渗透压约为0.2 MPa,这样高的盐浓度会抑制绝大多数农作物的生长[49]。
高盐对植物最直接的伤害是渗透胁迫和离子毒害。土壤中过多积累的盐分降低了土壤溶液的水势,使植物根系无法正常地吸收水分,甚至导致根系失水[50]。长期在盐土中生长的植物,通常还会遭受离子毒害。能产生毒害效应的离子有:Na+、Cl-、硫酸盐、碳酸氢盐等[49]。当土壤中存在过量的NaCl 时,植物体内Na+和Cl-就会过量积累,植物体内有异常高的钠钾比和高浓度的总离子含量,这会使细胞内催化各种生化反应的酶失活并且会抑制酶蛋白的合成[51]。植物体内高浓度的Na+会从细胞质膜上替换Ca2+,这会破坏细胞膜的选择透过性从而导致细胞内养分K+和P 的外流;土壤溶液中过量的Na+和Cl-还会抑制植物根系对Ca2+、Mn2+、Mg2+和N 的吸收,而这些离子对作物的生长和生殖是必需的[52]。叶绿体内积累高浓度的Na+和Cl-时,光合作用中的光合磷酸化和碳同化过程也会被抑制。这是因为光合系统中的电子传递链等光合结构对盐胁迫非常敏感[53]。
盐胁迫条件下,植物细胞内活性氧(ROS)成分的合成和清除的平衡被打破,使得细胞内的活性氧过度积累,造成膜蛋白和膜脂分子过氧化和脱酰化,导致生物膜系统遭受破坏。过量的ROS 还会破坏DNA 复制过程、改变核酸的碱基结构,进而损伤细胞内的遗传物质。细胞内许多催化生化反应的酶蛋白含有巯基和色氨酸残基,过度积累的活性氧会与这些基团发生反应,导致酶蛋白失活以及细胞内生理过程紊乱[54-55]。
5 含ACC 脱氨酶的根际促生细菌提高植物抗盐性的作用机制
研究表明,随着盐害时间的延长,番茄叶片中乙烯的合成量升高,其增加的幅度与Na+累积的量相关[15,56]。用100 mmol/L NaCl 处理两周,发现根组织、木质部汁液和叶片中ACC 浓度升高[57]。盐害诱导植物体内胁迫乙烯的合成,高浓度的乙烯通常会抑制植物生长[18]。
含有ACC 脱氨酶的根际细菌能够降低胁迫乙烯的水平,使植物在胁迫条件下生长的更好。为了验证这一假说,Mayak 等[15]从干热和盐土中分离出含ACC 脱氨酶的根际促生细菌,并发现这种根际细菌能促进植物在高盐环境中生长。在Mayak 等[15]的开创性实验之后,很多实验室都成功的利用含ACC脱氨酶的根际促生细菌来缓解高盐对植物的伤害(表1)。
植物受到渗透胁迫时,通常会表现出气孔阻力增大、渗透势降低、木质部平衡压升高等特点[58]。Amor 等[59]报道在NaCl 胁迫条件下生长的柿子椒(Capsicum annuumL.)幼苗的气孔阻力会显著升高,但接种根际细菌Azospirullumbrasilense和Pantoea dispersa后这种效应得以缓解。这可能是由于接种根际细菌后根系的导水率(L)提高了,正如其他一些研究所报道的干旱胁迫和盐胁迫条件下接种根际细菌也能提高根系导水率[60]。接种根际细菌后根系导水率的提高很可能是由于含有ACC 脱氨酶的根际细菌能降低根系乙烯的合成量,因为乙烯会降低水通道蛋白的活性从而降低根系的导水率[61]。Wang 等[62]研究表明接种根际细菌Variovorax paradoxus5C-2 后豌豆渗透势显著提高,木质部平衡压降低,这表明植物拥有更好的水分状态[63]。一些含ACC 脱氨酶的根际细菌可以促进根系生长从而提高根系的吸水面积并进一步提高根系的吸水量[64]。

表1 含ACC 脱氨酶的根际细菌提高植物耐盐性的主要相关文献列表
已有研究报道了植物应答含ACC 脱氨酶的根际促生细菌的过程中,植株地上部K+/Na+会升高[62,65]。Wang 等[62]利用养分流动模型,详细的解析了盐胁迫条件下,含ACC 脱氨酶的根际细菌V.Paradoxus5C-2 对豌豆体内的K+和Na+在木质部和韧皮部中运输的动态影响,以及这两种离子在植物体内的分配状况,实验结果表明接种根际细菌能提高根系吸收K+的量、木质部中由根系向地上部运输的K+量,这保障了通过木质部向地上部提供充分的K+,与此同时Na+在根系中积累,木质部中运往地上部的Na+降低,提高了地上部的K+/Na+,从而维持地上部的生长。Zhang 等[66]报道接种根际促生细菌B. subtilisGB03 后可以调控拟南芥根系及地上部的钾离子运载体HKT1 的表达,并同时降低Na+的水平和Na+在整株植物体内的再循环过程,结果导致拟南芥地上部的K+/Na+比值升高。Quadros 等[67]也报道了接种根际细菌Azospirillum后,玉米体内的K+/Na+比值也显著提高了。此外,还有其他科研工作者报道了接种根际细菌可以调控K+高亲和性运载体(AtHKT1)的表达以提高植物体内K+含量以及K+/Na+比值[68-70]。众所周知,在盐胁迫条件下,维持高钾钠比对植物体正常的气孔运动、酶的活化、蛋白质的生物合成、细胞的渗透调节、光合作用以及膨压的维持而言都是至关重要的,因此接种这些根际细菌后K+/Na+比值的提高就有利于维持植物体内许多生理过程正常进行。接种根际细菌后,植物根系Na+累积量的提高可能是由于一些基因表达量发生变化(如Na+载体HKT1 和SOS1)[66,71],未来需要更多的分子实验解析这一过程。
盐害不仅会降低植物体内Ca2+和K+的有效性,而且还会降低Ca2+和K+向植物生长部位的移动性和运输能力,同时还会造成其他矿质元素失衡。然而,许多研究表明接种含ACC 脱氨酶的根际促生细菌可以改善矿质营养失衡的状况。Fu 等[72]称给盐胁迫条件下生长的茄子接种假单孢菌后,植物地上部的Ca2+含量显著高于不接种的对照组。Yao 等[73]阐明接种植物根际促生细菌可以显著提高棉花对Mg2+和Ca2+的吸收量。Wang 等[62]发现接种根际细菌V.paradoxus5C-2 不仅有利于维持植物体内的K、Na平衡,还能促进对Ca2+、Mg2+、P 的吸收和积累。
植物根际促生细菌可以通过帮助植物积累渗透调节物质来提高植物的胁迫耐受性。脯氨酸是胁迫条件下含量变化最为明显的游离氨基酸,它可以增加植物细胞的渗透调节能力并且维持大分子物质结构的稳定性,是植物应答盐胁迫的一个重要的生理指标[74],Nia 等[75]报道了接种含ACC 脱氨酶的根际促生细菌后,小麦组织中含有更高水平的脯氨酸。Wang 等[76]从中国新疆的盐碱地分离出了13 个具有ACC 脱氨酶活性的根际菌株,其中芽孢杆菌属的一个菌株WU-9 提高了辣椒脯氨酸水平,显著增加了盐胁迫条件下辣椒植株的生物量。另有研究表明,中度盐胁迫条件下,接种含ACC 脱氨酶的内生型根际细菌Arthrobactersp. 和Bacillussp. 提高了甜椒(Capsicum annuum)组织中脯氨酸含量,并且还发现接种细菌后胁迫诱导的基因CaACCO(ACC 氧化酶)和CaLTPI(脂质转移蛋白)下调[77]。
盐胁迫条件下活性氧(ROS)过量积累会破坏植物膜系统,降低蛋白质的数量。目前已经确定,根际细菌通过触发细胞抗氧化水平来诱导植物的抗应激能力[78]。在2018 年的一项研究中,Heydarian等[79]观察到,表达细菌acdS基因的转基因油茶植株通过调控与活性氧水平相关基因的表达来防止细胞损伤,从而表现出更高的耐盐性。接种含ACC脱氨酶的根际细菌Stenotrophomonas maltophilia使小麦组织中超氧化物歧化酶、过氧化氢酶和过氧化物酶活性显著升高[80]。在75 mmol/L NaCl 条件下,用Enterobactersp. UPMR18 接种秋葵(Abelmoschus esculentus)后,植株体内抗氧化酶活性和ROS 通路基因的转录水平都升高,植物表现出更高的耐盐性[81]。
植物体遭受胁迫时,光合作用是最敏感的生理过程,光合电子传递受高盐胁迫的影响非常剧烈[82]。众所周知,高浓度的NaCl 能通过渗透胁迫诱导气孔关闭,还会加速膜蛋白降解等过程[83],从而导致光合系统的结构以及电子传递链破坏,进而光合作用受抑制。光合作用受抑制通常与类囊体膜上电子传递复合体的活性下降有关,这些复合体包括与放氧复合体相关蛋白和反应中心蛋白等。相对而言,光系统II 比光系统I 对盐胁迫更为敏感、更易受损伤。Wang 等[62]用Mini-PAM(Pulse-amplitude-modulation)测定大豆成熟叶片的最大光合潜能(Fv/Fm)和最大电子传递效率(最大ETR),其研究结果表明在盐胁迫条件下给豌豆幼苗接种根际细菌V. paradoxus5C-2 缓解了盐胁迫对光合作用的不利影响,豌豆幼苗的Fv/Fm 和ETR 都提高,说明接种根际细菌V.paradoxus5C-2 有利于提高盐胁迫条件下植物的光合作用。Win 等[84]利用内生型的含ACC 脱氨酶的Pseudomonasspp. 提高了遭受盐胁迫的番茄的光合速率。
接种含ACC 脱氨酶的根际细菌提高了遭受盐胁迫的植株的光合效率,这可能是因为接种细菌后植物体内的水分关系和离子平衡得以改善,ROS 的水平维持在合适范围,除了这些因素的变化外,接种这类细菌还提高了植物叶绿素含量,叶绿素含量的增加是植物耐盐性的指标之一。含ACC 脱氨酶的假单胞菌菌株处理番茄后,导致其叶绿素含量升高[85]。Pourbabaee 等[86]从伊朗的根际的土壤中分离和定性了一个盐生菌株Bacillus mojavensisK78,它具有ACC 脱氨酶活性,并使在盐胁迫条件下生长的小麦叶绿素含量增加。另有研究结果显示,接种含ACC脱氨酶的根际细菌Stenotrophomonas maltophilia使小麦组织中叶绿素含量升高[80]。这类细菌导致遭受盐害的植株叶绿素含量升高应当归因于乙烯含量的降低。大量实验结果表明当植物体遭受干旱、盐害等非生物胁迫时,乙烯的过量积累会诱导植株叶片叶绿素的降解,因为乙烯能促进叶绿素降解酶相关基因的表达[87]。降低乙烯在植物体中的累积量将会延缓叶绿素的降解,增加叶绿素的含量[88]。
大量研究结果表明含ACC 脱氨酶的根际促生细菌能帮助植物抵御盐胁迫,并且类似的实验结果还在不断增多。尽管含ACC 脱氨酶的根际细菌同时还产生其他物质如生长素、赤霉素、铁载体等[8,80],它们也可能在促进植物生长方面发挥作用。但是有大量文献报道,植物生长促进细菌(PGPB)中ACC 脱氨酶活性的存在是细菌促进植物生长,提高植物抵御逆境的主要机制[89-90],能够证明这一点的直接证据来自acdS突变细菌。例如,acdS突变的Pseudomonassp. 的ACC 脱氨酶活性明显下降,植物生长受阻,耐盐性降低[85,91](表1)。
6 展望
探究盐胁迫条件下含ACC 脱氨酶的根际细菌对植物的促进作用具有十分重要的理论意义和实践意义。已发表的文献结果皆在生理水平上系统地阐明了含有ACC 脱氨酶的根际细菌对植物水分关系、养分平衡、相容性溶质合成过程的改善。未来还需要从核酸水平和蛋白质水平上研究这类细菌对植物体内与这些生理过程相关的基因转录和蛋白表达的影响,从而全方位地阐明含ACC 脱氨酶的根际细菌提高植物耐盐性的机制。众所周知,乙烯与植物体内其他激素的信号相互整合,形成复杂的调控网络,因此还需重点研究接种这类细菌对盐胁迫下植物体内多种激素的调控作用。未来,还需了解土壤微生物在田间土壤中的适用性,大力开展长期的田间实验,为提高作物生产系统的抗盐能力提供有效的策略。
猜你喜欢
现代园艺(2022年7期)2022-11-19
林业科技(2022年5期)2022-10-08
浙江农业科学(2022年7期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
四川劳动保障(2021年3期)2021-06-09
水泵技术(2021年4期)2021-01-22
石油化工技术与经济(2021年4期)2021-01-13
天然产物研究与开发(2018年3期)2018-05-07
中国农业文摘-农业工程(2016年5期)2016-04-12
自动化博览(2014年9期)2014-02-28
