
枫香树林挥发物和空气负离子季节变化及对微生物数量的影响
2021-04-24 13:22马玲张静赵亚红王彬韦赛君吕嘉欣高岩
浙江林业科技 2021年2期
马玲,张静,赵亚红,王彬,韦赛君,吕嘉欣,高岩
(浙江农林大学,亚热带森林培育国家重点实验室,浙江 杭州 311300)
植物挥发性有机物(Plant volatile organic compounds,PVOCs)是通过植物体内次生代谢途径合成的低沸点、易挥发的小分子化合物,主要分为萜烯类、苯基/苯丙烷类和脂肪酸衍生物[1-2],具有抑菌、杀菌、净化空气、消除疲劳等功效[3-4]。林富平等发现木犀Osmanthus fragra ns叶片释放的VOCs(挥发性有机物)能降低空气中微生物的数量[5]。Gomez-Rodriguez等发现万寿菊Tagetes erect a释放的挥发物可降低番茄枯萎病病菌Fusarium oxysporumsp.的孢子萌发率[6]。南美蟛蜞菊Wedelia trilobata释放VOCs对水稻稻瘟病病菌Magnaporthe grisea、水稻纹枯病病菌Rhizoctonia solani和玉米纹枯病病菌的抑菌效果显著[7]。Fadil等发现百里香Thymus mongolicus、迷迭香Rosmarinus officinalis和香桃木Myrtus communis精油对鼠伤寒沙门氏菌Salmnella typhimurium具有协同抑菌作用[8]。柠檬Citrus limon精油和欧洲银冷杉Abies alba精油混合后能净化室内空气质量[9]。
空气负离子是由植物的尖端放电和光电效应产生,具有杀菌、降尘、改善情绪的作用[10]。研究发现气象因子、植被类型对空气负离子浓度影响较大。潘辉等发现不同林分中空气负、正离子浓度与空气温度、光照强度之间呈负相关,与空气湿度呈正相关[11]。对四种植被类型的空气负离子浓度调查发现自然林>经济林>绿地>农田[12]。空气负离子浓度在生态环境和空气质量评价体系中是不可或缺的重要指标之一。
枫香树Liquidambar formosana属金缕梅科Hamamelidaceae枫香树属Liquidambar落叶乔木。目前,对枫香树的研究主要集中在繁育[13]、生理生化[14]、光合[15]等方面,但对枫香树VOCs抑菌作用的研究较少。因此,本文以枫香树为对象,研究了枫香树单株释放的VOCs和枫香树林空气VOCs成分,测定了林内空气负离子浓度和微生物数量,分析了不同季节枫香树林的抑菌特性和空气负离子效应,以期为森林康养建设中的树种选择提供理论依据。
1 材料与方法
1.1 试验样地
试验样地设在浙江省杭州市临安区浙江农林大学东湖校区枫香树林教学实习基地,地理坐标为119°43'39"E,30°15'25"N,海拔约39 m,林下有人工栽植的菲白竹Pleioblastus fortunei和自然生长的马兰Aster indicus、爵床Justicia procumbens等草本植物,平均高度在30~50 cm,地被植物总盖度约30%,周边无其他树木、机动车及其他污染源影响。属中亚热带季风气候区,温暖湿润,四季分明,雨热同期,有梅雨季节,空气湿润。全年平均气温为16.4°C,年日照时数为1 939.0 h,1月温度最低,日平均气温为5°C,极端最低温度为-12°C;7月温度最高,日平均气温为30°C,极端最高温度为41°C,全年降水量为1 628.6 mm,年平均相对湿度在70%以上。
1.2 试验材料和试验林
枫香树树龄均在20 a左右,平均高约为15 m,平均胸径约为25 cm,植株长势良好,无病虫害和机械损伤。枫香树林为人工纯林,土壤为红壤,造林面积约900 m2,种植密度约为1 650株·hm-2,郁闭度0.8左右,平均枝下高约2 m,并以距林缘15 m处的无林地作对照。
1.3 VOCs采集与分析
枫香树单株VOCs测定:于2019年5月多次选择雨后晴朗无风天气的9∶00-11∶00,采用QC-2型大气采样仪(北京市劳动保护科学研究所),采用动态顶空气体循环采集法[16]采集活体枫香树单株的VOCs,根据树木平均高度采用五点采样法选择5株枫香树单株,每株作为1个独立实验,共5次重复,采样位置位于树冠向阳背风中部健康无病虫害枝叶,采集VOCs后采摘叶片称量。气体循环流量为100 mL·min-1,采气时间为1 h。
枫香树林分VOCs测定:于2019年春季(5月)、夏季(8月)和秋季(11月)多次选择雨后晴朗无风天气的9∶00-11∶00,采用开放式采样法采集枫香树林空气VOCs。在距地面1.5 m处对枫香树林空气VOCs进行采集,均匀的选择5个点,每个点作为1次独立实验,共5次重复。气体循环流量为100 mL·min-1,采气时间为1 h。
采用热脱附-气相色谱-质谱(TDS-GC-MS)联用技术分析VOCs,仪器及参数设置条件参照高群英等的方法[16]。采用NIST2008谱库检索,并根据已报道的植物VOCs保留时间对VOCs各组分进行定性,然后选取特征离子进行进一步定量分析。枫香树单株VOCs定量方法采用单位采样时间内每克植物叶片释放出的VOCs特征离子峰峰面积进行定量。

1.4 空气负离子的测定
采用YZQ-308A森林康养检测系统(翼鬃麒科技有限公司)于春季、夏季和秋季于枫香树VOCs测定同时检测枫香树林和对照空气负离子浓度,采样高度为1.5 m,时间25 min,每5 min记录一次数据,共5次重复(五点取样法选5个点)。
1.5 培养基的配制
参照周德庆[17]的方法配置细菌培养基(牛肉蛋白胨培养基)、真菌培养基(马丁氏培养基)、放线菌培养基(高氏1号培养基)。植物挥发性有机物单体培养基参照GAO Y等[18]的方法,无菌条件下,分别在柠檬烯、月桂烯、β-蒎烯、γ-萜品烯(Sigma公司生产)中加入两滴吐温,用无菌水将原液稀释为100、200、1 000倍,分别取稀释液1 mL加入培养皿中,再加入9 mL培养基,充分摇匀凝固后待用。
1.6 空气微生物的采集
2019年春季(5月)、夏季(8月)和秋季(11月),多次选择雨后晴朗无风天气的10:00,采用自然沉降法采集空气微生物(细菌、真菌和放线菌),分别在枫香树林与对照地随机各设5个采样点,每次5个重复,将带有不同培养基的培养皿分别放在距树冠水平距离0.3 m左右,高约1.5 m的平板支架上,打开培养皿盖,采集空气中微生物,将培养基在空气中暴露10 min后,盖上培养皿盖并用封口膜封好,置于30℃恒温培养箱中培养48~96 h,细菌培养48 h,真菌培养72 h,放线菌培养96 h后,检查菌落。
1.7 数据处理
按照1991年颁布的公共场所每立方米空气微生物的计算公式,计算出各处理的微生物浓度[19]。

式中,N为培养皿中菌落的平均数(个);A为培养皿的面积(cm2);T为培养皿盖打开的时间(min)。

式中,所有数据均为5次重复的平均值±标准误差。
利用Origin 9.0软件(OriginLab公司,美国)进行统计分析和作图。采用One-Way ANOVA对枫香树林和对照分别进行检验,并进行Tukey多重比较(P<0.05)。
2 结果与分析
2.1 枫香树挥发物成分分析
枫香树单株VOCs共鉴定出3类26种化合物(见图1,表1)。由图1和表1可知,VOCs以萜烯类物质为主要成分,共23种,主要有柠檬烯(22.4%)、月桂烯(12.3%)、β-蒎烯(13.7%)、γ-萜品烯(5.7%)、石竹烯(1.2%)、花柏烯(2.0%)等,占VOCs总量的96.6%;烃类物质仅1种为对伞花烃,占VOCs总量的1.4%;酯类物质有2种,分别为顺-3-己烯乙酸酯(1.4%)和酞酸二乙酯(0.6%),占VOCs总量的2.0%。

图1 单株枫香树VOCs总离子流图Figure 1 Total ion flow diagram of VOCs from single L.formosana
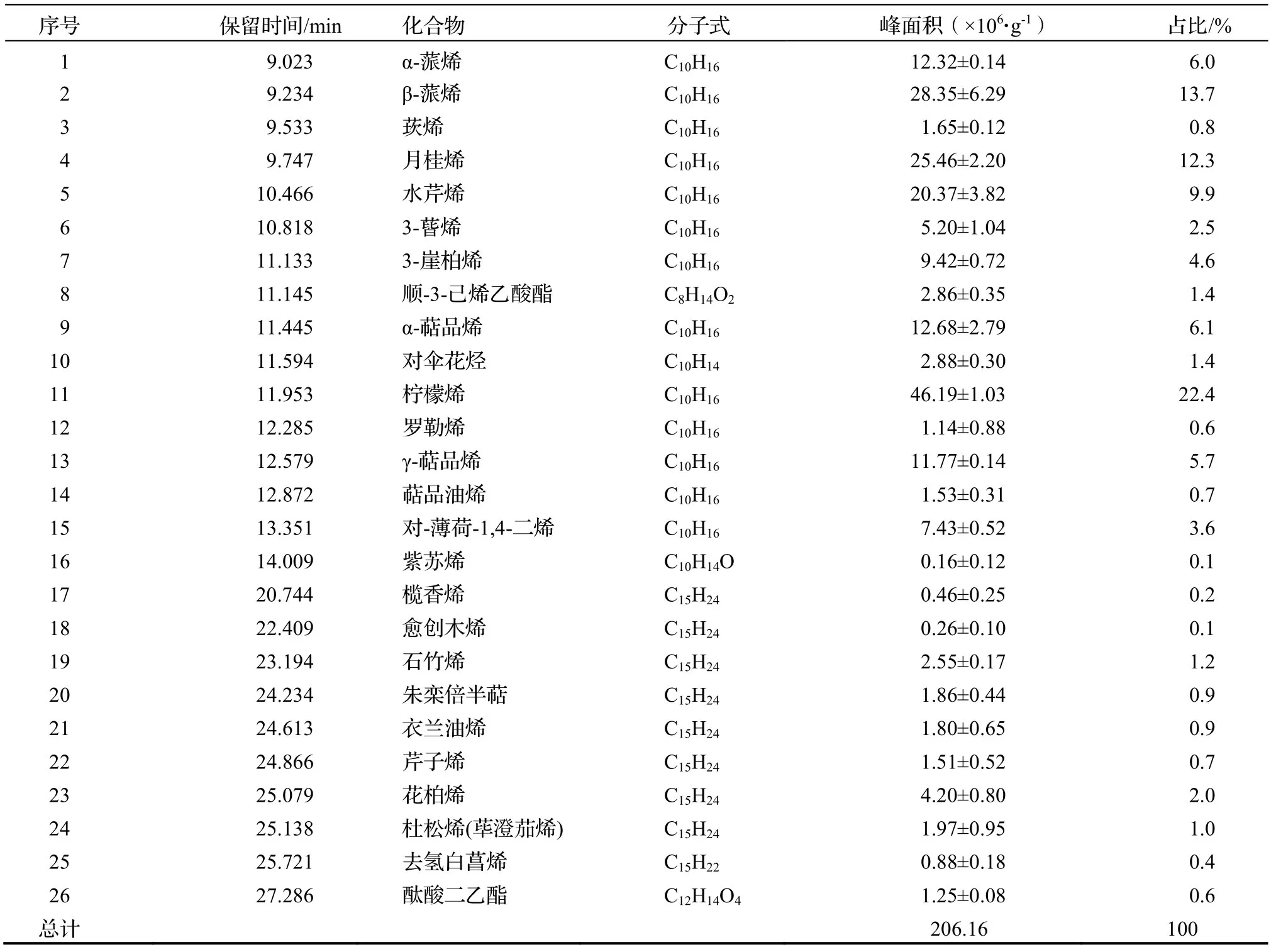
表1 枫香树叶片释放VOCs的主要成分Table1 Main component of VOCs from single L.formosana
2.2 不同季节枫香树林空气VOCs成分分析
由图2和表2知,春、夏、秋季枫香树林空气VOCs共有7类28种化合物,主要以萜烯类、苯类物质为主,不同季节VOCs的种类与含量存在差异,夏季总释放量分别是春、秋季的1.2倍、69.5倍。

图2 不同季节枫香树林空气VOCs总离子流图Figure 2 Total ion flow diagram of VOCs from L.formosana stand at different seasons
春季,共检测出5类21种化合物,其中,萜烯类物质有5种,主要为柠檬烯(13.3%)、β-蒎烯(9.7%)等,占VOCs总量的28.4%;苯类物质有8种,主要为甲苯(16.5%)、乙苯(12.4%)等,占VOCs总量的44.1%;烃类物质有5种,主要为1,3,5-环庚三烯(10.1%)、庚烷(5.7%)、1,2-二氯乙烷(3.0%)等,占VOCs总量的20.1%;酯类物质有2种,分别为乙酸丁酯(1.0%)和邻苯二甲酸二异丁酯(0.2%),占VOCs总量的1.2%;醇类物质有1种,为异植物醇,占VOCs总量的6.1%。

表2 不同季节枫香树林空气VOCs成分Table 2 Components of VOCs from L.formosana stand at different seasons
夏季,同样检测到5类21种化合物,其中,萜烯类物质有8种,主要成分为柠檬烯(16.6%)、β-蒎烯(10.6%)、月桂烯(4.6%)等,占总量的40.5%;苯类物质有7种,以甲苯(22.6%)、乙苯(4.6%)为主要成分,占VOCs总量的35.7%;酯类有2种,分别为乙酸丁酯(2.7%)和邻苯二甲酸二异丁酯(9.1%),占VOCs总量的11.8%;烃类物质有3种,主要为庚烷(4.3%),占VOCs总量的6.8%;醇类物质仅1种,为异植物醇,占VOCs总量的5.1%。
秋季,共检测到4类6种化合物,其中,苯类物质有3种,主要成分为甲苯(23.6%)、乙苯(6.3%)、邻二甲苯(15.4%)等,占VOCs总量的45.2%;酸类物质有1种,为十六烷酸,占VOCs总量的35.4%;萜烯类物质有1种,为α-蒎烯,占VOCs总量的13.8%;酮类物质有1种,为2-十二烷酮,占VOCs总量的5.6%。
夏季,在对照空气中共检测到3类12种化合物,其中苯类物质有8种,占VOCs总量的87.4%;烃类物质有2种,占VOCs总量的4.6%;酯类物质有2种,占VOCs总量的8.0%。不同季节枫香树林释放的VOCs与对照对比,春、夏季枫香树林内的VOCs总释放量分别是对照的5.1和6.1倍,秋季枫香林内的VOCs总释放量低于对照。
2.3 不同季节枫香树林空气负离子浓度变化
由图3知,不同季节枫香树林与对照空气负离子浓度变化趋势均为夏季最高,春季次之,秋季最低。夏季对照的空气负离子浓度分别比春季和秋季增加了39.3%和55.6%,3个季节之间均差异显著(P<0.05);夏季枫香树林空气负离子浓度分别比春季和秋季增加了41.9%和65.9%,与对照间均差异显著(P<0.05);春、夏、秋季枫香树林空气负离子浓度分别比对照增加了61.8%、68.7%和29.5%,与对照间均差异显著(P<0.05)。
2.4 不同季节枫香树林空气微生物含量变化
由图4知,枫香树林内微生物数量在不同季节有一定的动态变化。夏季枫香树林内的细菌含量比春季和秋季分别降低了112.0%和166.0%,与这二个季节之间均差异显著(P<0.05);春、夏、秋季林内细菌含量与对照相比分别降低了27.9%、49.5%和18.9%,与对照之间均差异显著(P<0.05)。

图3 不同季节枫香树林空气负离子浓度变化Figure 3 Variation of negative ion concentration in L.formo sana stand at different seasons.
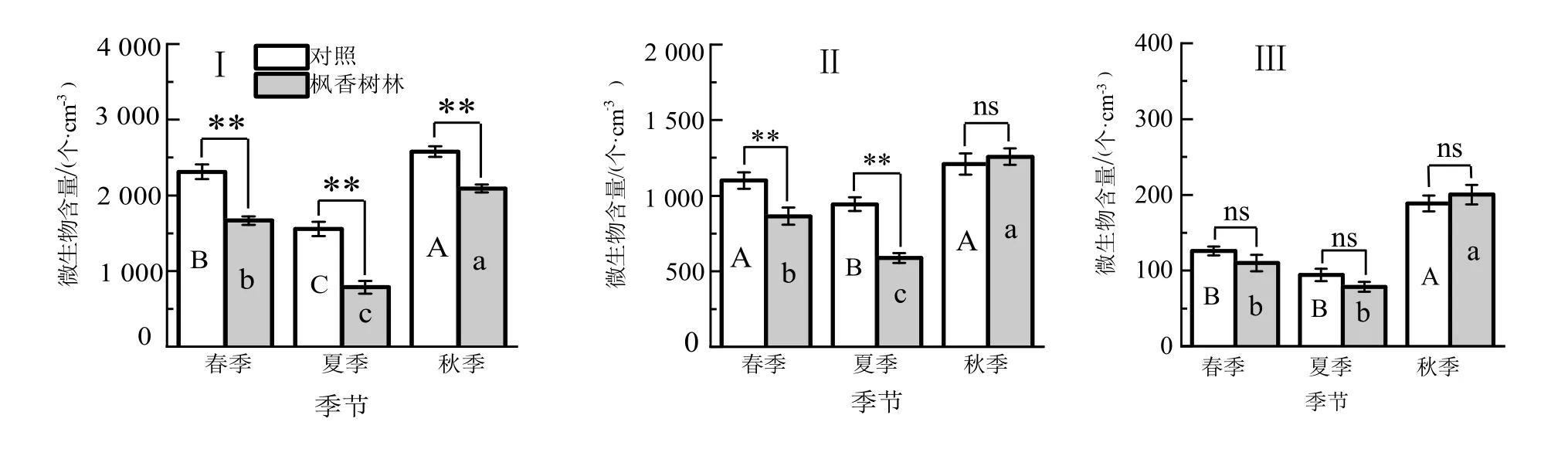
图4 不同季节枫香林空气微生物含量Figure 4 Air microbe content in L.formosana stand at different seasons
夏季枫香树林内的真菌含量比春季和秋季分别降低了48.6%和132.4%,与这二个季节之间均差异显著(P<0.05);春、夏季与对照相比分别降低了21.4%和38.3%,与对照之间均差异显著(P<0.05)。放线菌数量变化趋势与真菌一致,春、夏季与对照相比分别降低了12.6%和16.7%,与对照间均差异显著(P<0.05)。
2.5 VOCs单体对空气微生物的抑制
由图5知,随着单体浓度的升高,β-蒎烯、柠檬烯、月桂烯和γ-萜品烯与对照相比对细菌的抑制作用增强,0.1%浓度β-蒎烯和柠檬烯处理与对照相比对细菌的抑制率分别为30.0%和34.6%(P<0.05),1%浓度月桂烯和γ-萜品烯处理与对照相比对细菌的抑制率分别为31.4%和21.2%(P<0.05);1%浓度月桂烯、柠檬烯、γ-萜品烯处理与对照相比对真菌抑制率分别为28.7%、35.3%和32.1%(P<0.05),随着浓度的升高三种萜烯类处理与对照相比对真菌的抑制作用加强;0.1%浓度柠檬烯、β-蒎烯、月桂烯、γ-萜品烯处理与对照相比对放线菌的生长影响不大,1%浓度时柠檬烯处理与对照相比对放线菌抑制率为27.8%(P<0.05)。
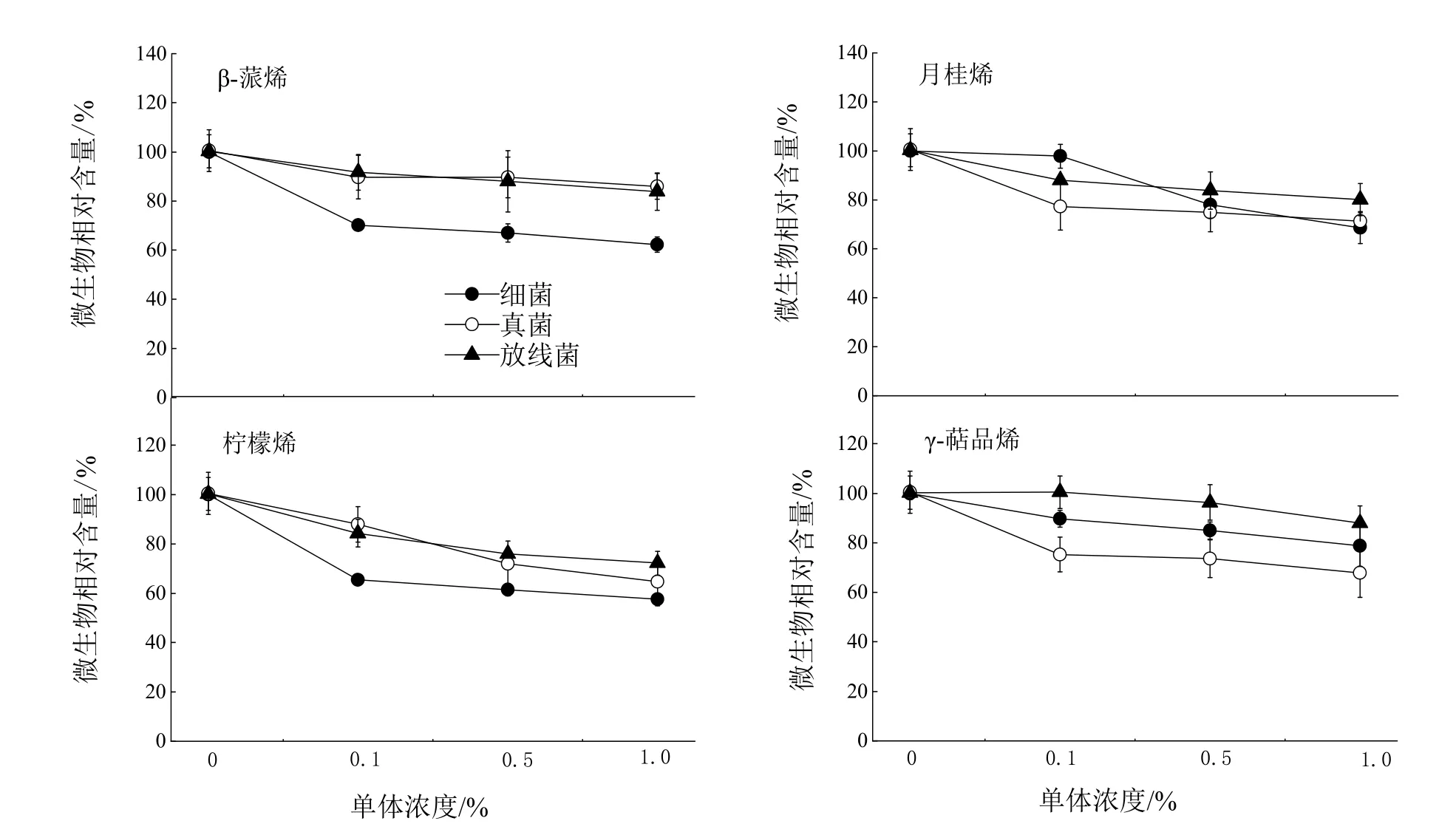
图5 单体对空气微生物的抑制Figure 5 Inhibition of monomers on air microorganisms
3 讨论
植物释放VOCs的种类及含量与植物生长发育阶段、体内合成相关酶的活性和环境因子密切相关。辣薄荷Mentha pi perita幼叶期释放的VOCs以柠檬烯为主,随着叶片的生长发育,释放的VOCs以薄荷酮为主[20]。光照使黄花蒿Artemisia annuaβ-蒎烯合酶的表达水平增高[21]。木犀在盛花期释放的萜烯类种类和含量高于衰败期[22]。高温能促进桉Eucalyptus camaldulensis和香蕉Musa nana叶片的单萜释放[23-24]。迷迭香在干旱处理后其萜烯类的释放量增加[25]。本研究中,枫香树林空气中的萜烯类含量主要由枫香树的叶片释放,春季、夏季和秋季随着叶片生长发育阶段的不同,萜烯类的种类和含量也不同,夏季成熟叶片释放的萜烯类含量最高,秋季叶片衰老,VOCs大量减少。其原因一方面可能是受环境因素(光照、温度、湿度)的影响,夏季光照时间变长,温度逐渐升高,植物进入生长旺盛期,体内有机物合成、代谢加快,释放VOCs的种类和含量增多[26-27];同时,植物旺盛的光合作用为萜烯类合成提供充足的碳源和还原力,促进萜烯类合成[28]。秋季气温降低、叶片衰老,叶绿素降解,花青素含量增多,叶片变红,花青素吸收的光不能用于光合作用,合成萜烯类的碳源和能量大量减少。另一方面可能是受相关生物合成酶的影响所致[29],异戊烯二磷酸为底物,在萜类合酶的催化下合成单萜、倍半萜和二萜等,萜类合酶的活性和数量决定了萜类化合物种类和结构的多样性[30]。这可能导致VOCs种类和释放量多样化。
空气负离子浓度是衡量空气清洁度与生态环境的重要指标之一,由植物尖端放电和光电效应产生,具有降尘、杀菌、净化空气的作用,其浓度受气象因素、植被自身特性的影响[11-12]。冯燕珠等研究发现福州闽江公园空气负离子浓度在秋季最高[31]。吴际友对空气负离子浓度研究发现夏、秋季大于春、冬季[32]。这可能与环境因子、林分及观测地区春、秋季的差异性有关。本研究中,春、夏、秋季枫香树林内的空气负离子浓度随着VOCs的变化呈同步变化,以夏季林内空气负离子浓度最高。这一方面可能是植物VOCs对空气负离子的促进作用,植物VOCs中萜烯类是可以挥发的中性气体分子,当受到碰撞(喷筒效应)、电场力(路德格效应)、辐射(光电效应)等外力作用影响时能产生较多的空气负离子[33];另一方面可能是夏季降水增多、植物生长旺盛、光合作用强,太阳紫外线强烈等因素产生了较多的空气负离子,秋季温度降低,叶片衰老,光合作用较弱,植物逐渐进入休眠状态,林内的空气负离子浓度减少。
植物释放的VOCs具有抵抗病原微生物入侵、生长和繁衍,减少或杀死空气微生物的作用[6,34]。Wang Y等研究发现肉桂醛可以显著减少中华猕猴桃Actinidia Chinensis在贮藏期间微生物的数量[35]。本研究中,夏季枫香树林释放的VOCs以萜烯类为主,林内细菌、真菌、放线菌含量低于春季和秋季,不同季节枫香树林内的细菌、真菌含量均显著低于对照。魏琦等研究发现乙酸乙酯能抑制细菌生长[36]。高群英等研究发现壬醛、柠檬烯、β-蒎烯能抑制微生物生长[16]。植物释放的VOCs对空气中的微生物生长有抑制作用[6-7]。为了进一步探索枫香树释放的VOCs中具体抑菌物质,选择萜烯类中含量较高的柠檬烯、β-蒎烯、月桂烯、γ-萜品烯进行单体抑菌试验,结果表明4种萜烯类均对细菌生长抑制作用显著,柠檬烯、月桂烯、γ-萜品烯对真菌生长抑制作用显著。柠檬烯、β-蒎烯、月桂烯、γ-萜品烯可能是主要的微生物抑制剂。
4 结论
上述研究结果表明,单株枫香树释放的VOCs共鉴定出3类26种化合物,以萜烯类为主,春、夏、秋季枫香树林中的空气VOCs共有7类28种化合物,主要以萜烯类、苯类物质为主,其中,春季共有5类21种化合物,主要有柠檬烯、β-蒎烯、甲苯等;夏季共有5类21种化合物,主要有柠檬烯、β-蒎烯、甲苯等;秋季共有4类6种化合物,主要有甲苯、邻二甲苯、十六烷酸等。不同季节VOCs的种类与含量存在差异,夏季总释放量是春、秋季的1.2倍、69.5倍,夏季枫香树林内萜烯类含量高于春季和秋季;不同季节枫香树林中空气负离子浓度变化趋势为夏季最高,春季次之,秋季最低;枫香树林内微生物数量在不同季节有一定的动态变化,夏季枫香树林内萜烯类含量最高,细菌和真菌含量显著降低,柠檬烯、β-蒎烯、月桂烯、γ-萜品烯可能是微生物主要抑制物质。
基于目前植物VOCs排放的物种变化和不同植物VOCs对空气负离子及微生物抑制作用的差异来看,建议在为城市环境选择树种时,除了考虑树木的适应性、抗污染性和观赏价值标准外,还应考虑植物的VOCs和空气负离子浓度。
猜你喜欢
红蜻蜓·中年级(2022年11期)2022-11-23
作物学报(2022年12期)2022-10-14
浙江林业科技(2022年1期)2022-02-20
中老年保健(2021年3期)2021-12-03
小学生优秀作文(低年级)(2021年10期)2021-11-24
科学与生活(2021年3期)2021-11-10
科学之谜(2021年4期)2021-07-09
林产工业(2020年2期)2020-03-30
现代农村科技(2018年5期)2018-05-31
橡塑技术与装备(2018年21期)2018-02-19
