
黄河源区发草适生地植物群落特征及其土壤因子解释
2021-04-23 03:21罗巧玉王彦龙杜雷刘念李丽马玉寿
草业学报 2021年4期
罗巧玉,王彦龙,杜雷,刘念,李丽,马玉寿*
(1.青海大学农牧学院,青海 西宁810008;2.青海省青藏高原药用动植物资源重点实验室,青海师范大学生命科学学院,青海 西宁810008)
植物群落是植物种群与环境因子共同构成的一个有机整体,不同物种种群在群落中所处空间分布格局取决于物种的生物学特性、生境条件及其交互作用[1-2]。探究植物群落多样性及其维持机制,是了解植物群落生态功能和稳定的基础,也一直是群落生态学研究的中心议题[3]。对石羊河下游不同类型荒漠草地黑果枸杞(Lycium ruthenicum)群落结构与土壤特性关系的研究发现,固定/半固定沙地和覆沙草地中黑果枸杞为优势种,砾质荒漠草地和盐渍化草地中碱蓬(Suaeda glauca)和狗尾草(Setaria viridis)为优势种[4]。在宁夏震湖滩涂湿地研究中,盐生灌丛柽柳(Tamarix chinensis)群落及盐生草甸盐角草(Salicornia europaea)群落、碱蓬群落和芦苇(Phragmites australis)群落土壤全盐在表层(0~10 cm)产生显著聚集效应[3]。水分、氮素、水氮交互对内蒙古乌兰察布市四子王旗短花针茅(Stipa breviflora)荒漠草原生物量具有显著影响[5]。由于生物、环境和空间3大因素的多样性,国内外学者就不同植物群落分布格局与环境之间的相互关系得到的结论不尽相同。但理论上讲,植物群落生态特征是植物与气候、土壤、地形等生态环境综合体长期适应和协同进化的结果。土壤资源生境因子影响植物群落结构、组成及多样性特征,植物与土壤之间发生着频繁的物质交换[3,6]。
发草(Deschampsia caespitosa)是禾本科(Gramineae)发草属(Deschampsia)多年生草本植物[7],具有耐寒、耐旱、耐水淹、耐盐碱、耐修剪等特性,是一种优良牧草[8]和城市园林地被植物[9-11]。此外,发草种子产量大、发芽率高[7,12],可以有效弥补高寒沼泽湿地典型物种如华扁穗草(Blysmus sinocompressus)、藏嵩草(Kobresia schoenoides)、青藏苔草(Carex moorcroftii)等植物因种子产量低和自然萌发率低[13-14]而很难应用于退化高寒沼泽湿地修复工作的不足,是青藏高原高寒沼泽湿地生态脆弱带植被恢复中值得重点研究的天然植物群落之一。
迄今为止,关于发草群落的研究仅见于其作为藏嵩草、青藏苔草群落伴生种的分布调查、起源等[15-16],而专门针对发草野生生境地调查及发草种群与环境因子关系的研究鲜有报道。本研究以发草适生地群落为研究对象,基于发草适生地群落和土壤及生物因子调查数据,采用相关性分析(correlation analysis)和冗余分析(redundancy analysis,RDA)方法,分析发草生境地植物群落物种结构组成、多样性分布和土壤理化特性及其关系,揭示影响发草种群在复杂多样的高寒沼泽异质生境中适应性的关键环境因子,为发草群落资源的合理开发利用及繁育建植提供科学依据,也为退化高寒沼泽湿地植被恢复工作的开展提供一定的理论基础。
1 材料与方法
1.1 研究区概况
研究区位于黄河源区玛沁县境内。玛沁县地处青海省东南部(33°43′-35°16′N,98°-100°56′E),属典型高原大陆性气候,具有多风、寒冷、辐射强、日照丰富等特征。年均降水量为260~770 mm,由西北向东南递增,并具有明显的区域分异性[17]。年均温-0.6℃(最高26.6℃,最低-34.9℃),年均日照时长2571.733 h。该地区土壤类型主要为高山草甸土,土壤有机质丰富、含水量较高。主要植被类型为湿地和高寒草甸。由于受气候条件、地貌特征以及土壤类型等综合影响,孕育了丰富、独特的生物种类与群落类型[18]。
1.2 研究方法
1.2.1 样地设置 通过查阅《青海省植物志》及其他文献资料[7,12,19],根据玛沁地区发草生长的分布范围和生境特征,确定5个典型分布区域(表1)。通过路线踏察在研究区域选取发草分布较集中、群落类型具有一定差异的样点,记录各样点的经纬度、海拔、生境等环境因子(表1)。
1.2.2 植物群落特征调查 2018年8月中旬植物生长旺盛期,在每个样点选择群落结构和组成分布均匀的区域,随机选取12~15个1 m×1 m的调查研究样方,调查群落内各物种的高度、盖度。随后分不同物种把地上部分齐根部剪下,去除枯落物后装至档案袋带回实验室,65℃烘干至恒重后称干重记录。
1.2.3 土壤样品采集 在测定完植物群落特征的各样方,将样方内土壤表面枯落物去除干净,用环刀分0~10 cm、10~20 cm土层取土样于干燥铝盒中准确称重,然后将其放于100℃烘箱中烘干至恒重称其重量,计算土壤含水量(soil water,W)。用内径为3.5 cm土钻分0~10 cm、10~20 cm土层取土样,每个取土点采集10个土样混合,装入密封袋带回实验室。自然风干后去除杂草、根系、石块等,过1、0.25和0.1 mm筛备用,其中过1 mm筛土壤用于测定pH值,过0.25 mm筛土壤用于测定有机质(soil organic matter,SOM)含量,过0.1 mm筛土壤用于测定全氮(total nitrogen,N)、全磷(total phosphorus,P)、全钾(total potassium,K)、碳(carbon,C)含量。
1.2.4 群落多样性分析 群落生物多样性包含生物丰富度和均匀度等,有关群落生物多样性的计算模型很多,差别在于对丰富度和均匀度这2个变量所赋予的权重不同[20-21]。本研究选用Simpson优势度指数(D)、Shannon-Wiener指数(H)、Alatalo均匀度指数(Ea)、Pielou均匀度指数(Jsi和Jsw)。计算公式如下:
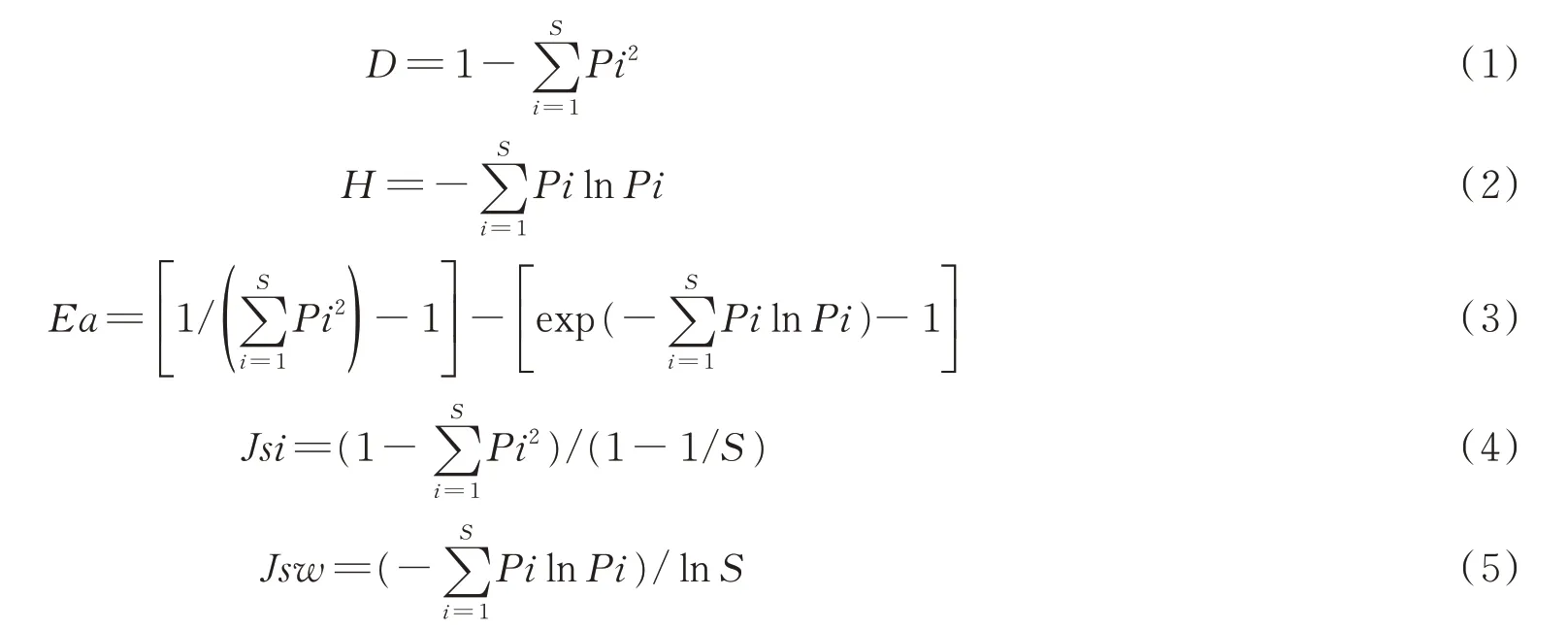
式中:Pi为相对重要值,Pi=(RC+RH+RB)/3,RC为相对盖度,RH为相对高度,RB为相对生物量;S为物种丰富度,即样方内出现的物种数。
1.2.5 土壤理化性质测定 参照土壤指标常规方法测定[22-23]。其中采用元素分析仪(CHNSO vario EL,德国Elementar)测定全氮、碳含量,采用钼锑抗比色法测定全磷含量,采用重铬酸钾滴定法测定土壤有机质含量,利用原子吸收光谱仪(M6 AA System,美国Thermo)测定全钾含量,采用酸碱度法测定土壤pH值(水土比为1∶1,仪器为PHS-3G,上海雷磁)。
1.3 数据分析
采用SPSS 19.0软件的完全随机设计模型,对不同发草适生地植物群落及土壤因子进行单因素方差分析;对发草种群特征与群落物种多样性、土壤因子进行相关性分析。采用CANOCO 4.5对发草适生地植物群落物种多样性指数与土壤因子进行冗余分析[24]。
2 结果与分析
2.1 发草适生地植物群落结构组成和植物多样性特征
在玛沁地区野外调查的所有样地中,共记录到83种物种,均为被子、草本植物,隶属于17科49属。其中含种数较多的科为菊科(Asteraceae)10属14种,禾本科(Gramineae)9属14种,莎草科(Cyperaceae)3属13种,毛茛科(Ranunculaceae)5属6种,龙胆科(Gentianaceae)3属6种,玄参科(Scrophulariaceae)3属6种。根据群落中物种重要值的大小将所有样地中的植物群落划分为5种群落类型(表1)。其中,Ⅰ:发草群落的优势物种为发草,次优势种为青藏苔草、垂穗披碱草(Elymus nutans),伴生种为鹅绒委陵菜(Potentilla anserina);Ⅱ:华扁穗草群落的优势物种为华扁穗草,伴生种为发草、高原嵩草(Kobresia pusilla)、甘肃嵩草(Carex pseuduncinoides);Ⅲ:华扁穗草+发草群落的优势物种为华扁穗草、发草,藏嵩草、细叶嵩草(Kobresia filifolia)、垂穗披碱草是伴生种;Ⅳ:藏嵩草群落的优势物种为藏嵩草,伴生种为发草、垂穗披碱草、华扁穗草、甘肃嵩草、细叶嵩草;Ⅴ:藏嵩草+发草群落的优势物种为藏嵩草、发草,伴生种为垂穗披碱草、鹅绒委陵菜、高原嵩草、细叶嵩草。

表1 发草适生地植物群落类型及其结构组成Table 1 The plant community type and its structure in D.caespitosa adaptive area
通过发草适生地植物群落物种多样性特征分析(表2)发现,各植物群落中物种多样性特征变幅较大。群落间物种丰富度(S)为4~27;Simpson优势度指数(D)为0.69~0.92;Shannon-Wiener指数(H)为1.27~2.82;Alatalo均匀度指数(Ea)为0.43~0.86;Pielou均匀度指数Jsi为0.87~0.97;Pielou均匀度指数Jsw为0.80~0.92。发草在各群落中所起作用也有很大差异,发草盖度(Cd)为1.33%~91.67%;发草高度(Hd)为37.08~75.40 cm;发草生物量(Bd)为11.33~681.78 g·m-2;发草重要值(Valued)为0.029~0.453。根据方差分析可知,群落Ⅲ的Simpson优势度指数、Shannon-Wiener指数、Pielou均匀度指数Jsi均显著大于其他群落(P<0.05)。群落Ⅳ的物种数达到27种,显著高于其他群落(P<0.05)。群落I的物种只有4种,但是其Alatalo均匀度指数和Pielou均匀度指数Jsw显著大于其他群落(P<0.05)。对于发草来说,群落I中发草的盖度、株高、生物量、重要值均显著高于其他群落(P<0.05)。
2.2 发草适生地土壤特征
植物群落的分布格局是不同尺度上经纬度、海拔、地形、气候、土壤等各种环境因子综合作用的结果。在微生境尺度上,土壤理化性质决定了植物群落类型及物种多样性。通过分析玛沁地区发草适生地植物群落土壤因子特征发现(表3),对于0~10 cm层,群落Ⅳ和Ⅴ的N、C、SOM、W含量显著高于群落Ⅰ、Ⅱ、Ⅲ(P<0.05),群落Ⅳ的P含量显著高于群落Ⅰ、Ⅱ、Ⅲ(P<0.05),群落Ⅰ、Ⅲ的pH显著高于群落Ⅱ、Ⅳ、Ⅴ(P<0.05);对于10~20 cm土层,群落Ⅴ的N、C含量显著高于群落Ⅰ、Ⅱ、Ⅲ、Ⅳ(P<0.05),群落Ⅳ的P含量显著高于群落Ⅰ、Ⅱ、Ⅲ(P<0.05),群落Ⅲ的K含量、pH显著高于群落Ⅰ、Ⅱ、Ⅳ、Ⅴ(P<0.05);群落Ⅴ的W含量显著高于群落Ⅰ(P<0.05)。
2.3 发草种群特征与主要环境因子间的相关性
从发草种群特征与群落物种多样性指标间相关性分析发现(表4),对于发草来说,其盖度和生物量与群落物种丰富度、Simpson优势度指数、Shannon-Wiener指数间存在极显著负相关关系(P<0.01),与Alatalo均匀度指数、Pielou均匀度指数Jsw间存在极显著正相关关系(P<0.01)。发草的株高与群落物种丰富度和Shannon-Wiener指数存在极显著负相关关系(P<0.01),与Simpson优势度指数存在显著负相关关系(P<0.05),与Alatalo均匀度指数存在显著正相关关系(P<0.05)。发草在群落中的重要值与群落物种丰富度、Simpson优势度指数、Shannon-Wiener指数间存在极显著负相关关系(P<0.001),同时与Alatalo均匀度指数和Pielou均匀度指数Jsw间存在极显著正相关关系(P<0.001)。

表2 发草适生地植物群落物种多样性特征Table 2 Diversity indexes of plant community in D.caespitosa adaptive area(Mean±SD)
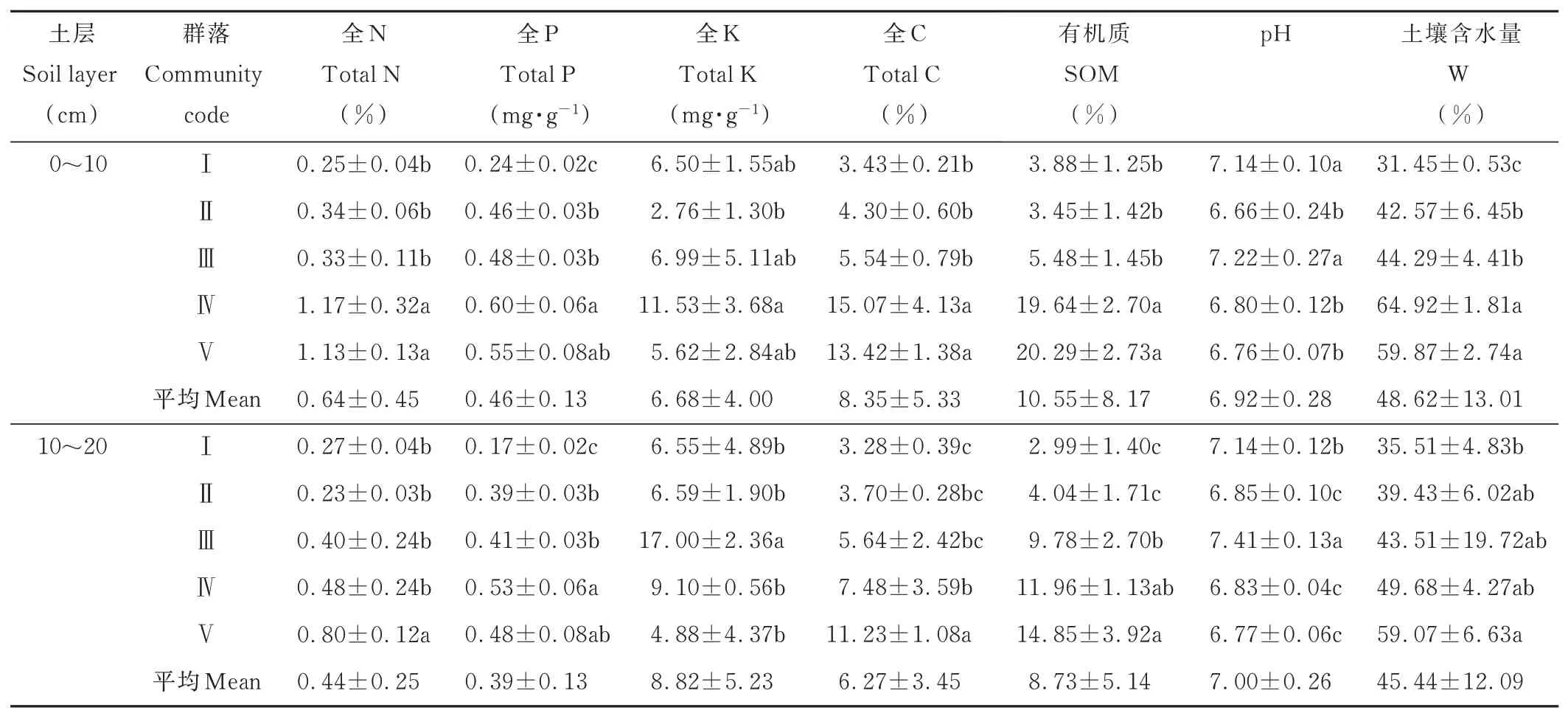
表3 发草适生地植物群落土壤特征Table 3 The general situation of plant community soil factors in D.caespitosa adaptive area(Mean±SD)

表4 发草种群特征与群落物种多样性间相关性Table 4 Correlation analysis of population characteristics of D.caespitosa and community species diversity
根据发草种群特征与土壤因子间相关性分析(表5),发草的盖度、生物量和重要值与0~10 cm土层P含量、水分含量及10~20 cm土层P含量间存在显著的负相关关系(P<0.05)。另外,作为研究某物种在群落中的地位和作用的综合数量指标,重要值与0~10 cm土层pH间存在显著正相关关系(P<0.05)。

表5 发草种群特征与土壤因子间相关性Table 5 Correlation analysis of population characteristics of D.caespitosa and soil factors
2.4 发草适生地植物群落与主要土壤因子的排序分析
RDA排序法能够有效对环境梯度下的多个环境指标进行统计检验,可以更好地反映群落物种多样性与环境因子之间的关系。RDA排序图中,蓝色箭头表示群落物种多样性指数,红色箭头表示环境因子。通过物种箭头之间的夹角来表示物种之间的相关性:其中夹角越大,表示相关性越小;如果箭头同向,表示正相关,箭头相反,表示负相关[25]。从发草适生地植物群落物种多样性指数与0~10 cm土层和10~20 cm土层的7个土壤因子进行的RDA排序图分析发现(图1),发草群落的D、S、H之间及Jsw、Ea、Cd、Bd、Valued之间均具有强相关性,D、S、H与Jsw、Ea、Cd、Bd、Valued之间具有强烈负相关性。土壤pH跟其他土壤理化特征之间呈负相关,0~10 cm土层中pH显著负向影响植物群落的D、S、H,而显著正向影响植物群落中的Jsw、Ea、Cd、Bd、Valued。

图1 发草适生地植物群落物种多样性指数与土壤因子的RDA排序Fig.1 RDA ordination of diversity indexes of plant community and soil factors in D.caespitosa adaptive area
3 讨论
3.1 发草种群及伴生群落结构特征
植物与植物之间均存在着广泛而复杂的相互作用。植物种群结构可反映种群的数量动态、发展趋势与环境之间的相互关系[26],也可揭示植物种群与生境相适应的结果,对预测群落未来发展趋势具有重要意义[27-28]。本研究黄河源区发草适生地植物群落的5个样地中,共记录到83种物种,隶属于17科49属。各群落的结构组成及物种多样性指数变幅较大,这可能是因为黄河源区地处青藏高原腹地,深居亚欧大陆内部,远离海洋,属典型高原大陆性气候[18]。常年气候干燥、寒冷,年温差和日温差都很大。日照丰富、辐射强。降水由西北向东南递增,并具有明显的区域分异性。境内地貌复杂,土壤类型多样。由于受复杂的地理、气候以及土壤条件等综合影响,形成了复杂多样的生境类型,孕育了丰富、独特的生物种类与群落结构组成[17-18]。这与卢慧等[29]在三江源区6个高寒草甸样地中发现共有21科51属74种植物的结果基本一致。本研究发现发草既可以成为某一群落的优势物种,也能够成为某一群落的亚优势种或者伴生种。某一物种在该群落中的功能或者角色受到生境地生物和非生物因素的综合影响[2]。
3.2 发草种群分布的主要环境解释
植物种群多样性特征及其空间分布格局能够反映环境对种群内个体生长和生存的影响,也能反映和指示植物种群的生态适应对策[30-31]。目前,关于环境因子与种群特征关系的研究多集中在特殊生境中植物种群对环境因子的适应与响应上。植物种群特征通常由最大限制环境因子决定,因此,不同植物种群特征与环境因子关系的研究结果并不一致。研究发现,四川牡丹(Paeonia szechuanica)种群密度平均基径受环境因子影响显著,其中坡度是影响种群特征的首要环境因子,其次是土壤自然含水率[32]。新疆天山中段典型中山带的天山云杉(Picea tianschanica)群落分布格局受人工采伐干扰因子和海拔、坡度、土壤含水量、土壤pH的影响显著[33]。很多研究表明,群落物种多样性指数受到土壤营养元素的影响。在库车山区,影响新疆假龙胆(Gentianella turkestanorum)适生地植物群落组成、物种多样性分布的主要环境因子是土壤N、P、可溶性K、pH[34]。通过对卡拉麦里山自然保护区准噶尔沙蒿(Artemisia songarica)群落结构与环境因子进行典范对应分析(canonical correspondence analysis,CCA)发现,N、P是影响群落物种分布的关键因子,可以通过适当提高土壤N、P含量来提高该区准噶尔沙蒿群落的稳定和健康发展[35]。但是,本研究结果与以上结果相反,本研究中黄河源区发草的盖度、生物量和重要值均与0~10 cm土层P和W含量及10~20 cm土层P含量间存在显著负相关关系。pH值是土壤重要的基本性质之一,与土壤的肥力状况、植物生长及微生物活动有密切关系。pH能够通过影响土壤溶液中各种离子的浓度影响营养元素的有效性,进而影响植物生长[36-37]。本研究中土壤pH跟其他土壤理化特征之间呈负相关,这与郭成久等[38]在草甸土壤理化性质特征研究中的结果一致。0~10 cm土层中pH显著促进发草植物群落中的Jsw、Ea、Cd、Bd、Valued。在三江源区高寒草甸植物多样性分布格局研究中也发现,土壤pH值逐渐升高时禾本科和杂类草植物的数量增加,这些植物比嵩草属植物更加耐旱、抗盐碱,对环境适应性更强[29]。CCA分析结果表明,土壤pH值可能是影响高寒草甸植物群落结构最重要的因素[29]。另外,本研究中发草的盖度、生物量和重要值均与群落物种丰富度、Simpson优势度指数、Shannon-Wiener指数间存在显著负相关关系。这可能是因为发草耐贫瘠,而研究区处于未退化高寒沼泽湿地和高寒草甸,土壤水分及营养相对充足,对水肥要求较高的藏嵩草等莎草类植物生长旺盛[39-40],其物种丰富度、Simpson优势度指数、Shannon-Wiener指数均较大,进而与发草产生对光照、空间、养分等的竞争。虽然关于发草对土壤不同P含量及pH的响应的研究尚未见报道,但任青吉等[41]对高寒沼泽化草甸上的华扁穗草、草地早熟禾(Poa pratensis)、发草、湿生扁蕾(Gentianopsis paludosa)等包括莎草、禾本科草、双子叶类杂草的51种植物进行了叶片形态特征和光合生理的比较研究,发现发草的水分利用效率(water use efficiency,WUE)最高,达到3.76μmol CO2·mmol-1H2O;而蒸腾速率(transpiration rate,Tr)最低,只有4 mmol H2O·m-2·s-1。高的水分利用效率和低的蒸腾速率保证了发草可以在水肥状况较差的土壤上正常生长。
4 结论
本研究利用相关分析和RDA排序法,探讨了黄河源区发草种群特征与群落物种多样性特征,揭示了发草种群特征与群落物种多样性指标及土壤因子间的相互关系。本研究发现,发草的重要值显著受群落物种丰富度、Simpson优势度指数与Shannon-Wiener指数的负向影响,同时显著受0~10 cm土层pH的正向影响,说明发草具有先锋植物的特点。另外,发草具有种子产量大、发芽率高、抗逆性强等优质特征。因此,建议在对退化高寒沼泽湿地、高寒草甸、高寒草原等生态系统开展植物修复的过程中可将发草作为先锋物种之一进行优化植物配置。研究发草种群特征与环境之间的关系,对发草资源的合理开发利用具有重要价值。
本研究由于调查取样范围较小,测量植物个体功能性状和群落特性的指标较少,使得研究结果具有一定的局限性,但是本研究从植物种群特征及其环境因子解释的角度为治理退化高寒草地提供了部分理论依据,从而建议未来在治理退化高寒草地时,应考虑其退化程度,根据不同的退化程度选择相适应的植物配置。
猜你喜欢
今日农业(2022年15期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
河南科学(2020年3期)2020-06-02
河南畜牧兽医(2020年21期)2020-01-10
红土地(2018年7期)2018-09-26
北京航空航天大学学报(2017年3期)2017-11-23
草业科学(2015年9期)2015-12-11
中国药业(2014年17期)2014-05-26
中成药(2014年10期)2014-02-28
