
蓖麻地上部主茎器官形态建成的预测模型
2021-04-21 09:52:26徐寿军德木其格王金波齐海祥朱国立
作物杂志 2021年1期
徐寿军 王 磊 德木其格 王金波 齐海祥 朱国立 ,5
(1内蒙古民族大学农学院,028043,内蒙古通辽;2内蒙古自治区高校蓖麻产业工程技术研究中心,028043,内蒙古通辽;3内蒙古自治区蓖麻育种重点实验室,028043,内蒙古通辽;4内蒙古自治区蓖麻产业协同创新培育中心,028043,内蒙古通辽;5内蒙古通辽市农业科学研究院,028015,内蒙古通辽)
虚拟作物研究是借助计算机模拟作物生长发育和空间形态变化过程,为作物研究提供了一种新的方法,在农业、林业、生态和遥感等领域得到广阔的应用,已成为农业信息技术研究的热点之一[1-4]。而作物器官形态模拟模型是虚拟作物研究的关键技术,它是基于三维空间实现作物可视化的基础。目前,众多学者以多种作物为对象,构建了大量的作物地上器官形态建成预测模型[5-9]。其中,关于水稻[10]、小麦[11]和玉米[12]等禾本科作物器官形态建成的动态模型的研究较为系统深入,不仅模拟了茎秆、叶片和果穗等重要器官的生长动态,而且构建了叶鞘、茎鞘夹角、根系和叶片曲线乃至叶色的生长变化动态模型,为这些作物的可视化研究奠定了良好基础。而棉花[13-14]、大豆[15]和蓖麻[16]等双子叶作物因具有无限生长习性,器官种类多且结构复杂,对其器官形态建成的模拟研究还处于初步探索阶段。尤其是关于蓖麻主茎形态建成的预测模型研究鲜见报道。本研究选择地上器官差异较大的蓖麻品种,以生理发育时间为模拟尺度,在系统观测的基础上,构建了蓖麻地上部器官形态建成的预测模型,为开展蓖麻的可视化研究提供理论基础。
1 材料与方法
1.1 供试材料
参试品种为通蓖9号、通蓖10号、通蓖11号和通蓖13号,均由内蒙古通辽市农业科学研究院提供。其中通蓖11号和通蓖13号植株较矮,茎秆较细;通蓖9号和通蓖10号植株较高,茎秆较粗。
1.2 试验设计
1.2.1 播期试验 于2016年在内蒙古通辽市农业科学研究院进行播期试验,设4月17日、5月7日和5月27日3个播期。分2次施纯氮120kg/hm2,其中基肥84kg/hm2,追肥36kg/hm2,于现蕾前施用。磷肥和钾肥作为基肥一次性施用,用量分别为30和60kg/hm2。行距65cm,株距70cm,小区面积90m2。随机区组设计,3次重复。栽培管理与当地大田生产相同。
1.2.2 氮肥试验 于2017年在通辽市科尔沁区农牧业高新科技示范园区进行氮肥试验,设0、60、120和180kg/hm2共4个氮肥水平(纯氮),基追比7∶3,追肥在现蕾期前施用。磷肥和钾肥作为基肥一次性施用,用量分别为30和60kg/hm2。5月3日播种,行距65cm,株距60cm,小区面积60m2。随机区组设计,3次重复。栽培管理与当地大田生产相同。
1.3 测定项目和方法
1.3.1 生育期 在蓖麻主茎第1片复叶开始展开时,每小区选择株高、茎粗和叶片扩展等长势接近的3株蓖麻进行标记。记录标记植株第3复叶、主茎各节间、主茎果穗以及蒴果生长开始和结束的日期。
1.3.2 农艺性状 在每个节间生长开始至生长结束的时间段内,每3d测定1次标记植株主茎的节间长度和直径,使用直尺测量节间长度,使用游标卡尺测量各节直径(节间最粗处)。蓖麻同一品种各个节位复叶的形态基本相似。选取主茎第3复叶作为模拟对象,在第3片复叶开始出现至停止生长的时间段内,每3d测量1次叶长(叶片中心的最长主脉的长度)和叶宽(与最长主脉垂直的两叶裂间的最大距离)。自主茎果穗出现开始,每3d用直尺测量1次果穗长。选取主茎果穗的某一蒴果标记,每3d用游标卡尺测量1次蒴果长和蒴果直径(蒴果最粗处)。
1.3.3 植株含氮量 各处理分别在出苗期、现蕾期、开花期、结果期和成熟期取样,105℃下杀青0.5h,80℃下烘48h至恒重,粉碎后采用凯氏定氮法[17]测定氮含量。
1.4 数据处理
用播期试验中5月7日和5月27日播种的数据及氮肥试验数据构建模型,并调试参数,用播期试验中4月17日播种的数据检验模型。
1.5 模型构建方法与步骤
以生理发育时间作为模拟尺度。生理发育时间(physiological development time,PDT)是指在最适温光条件下,作物完成某一发育阶段所需的最短时间。对于某一特定基因型品种来说,完成某一特定的生育阶段所需的生理发育时间基本恒定[18]。蓖麻各生育阶段所对应的生理发育时间见文献[19]。
在分析节间长度、节直径、叶片长度、叶片宽度、果穗长度、蒴果长度和蒴果直径随PDT动态变化的基础上,运用Richards方程W=A/(1+be-kt)m构建各器官形态建成预测模型,采用顾世梁等[20]提出的缩张算法对b、k和m等参数进行确定。
2 结果与分析
2.1 主茎生长预测模型
2.1.1 节间伸长模型 根据文献[19]的计算方法,运用本试验的观测数据计算可知,蓖麻第一节间开始伸长的生理发育时间为10.9d,主茎各节间从开始伸长到定长需要22.1个生理日,蓖麻主茎各节间的生长具有一定的重叠性,存在叠次生长的现象,相邻节间叠次伸长间隔1.7个生理日,见式(1)。
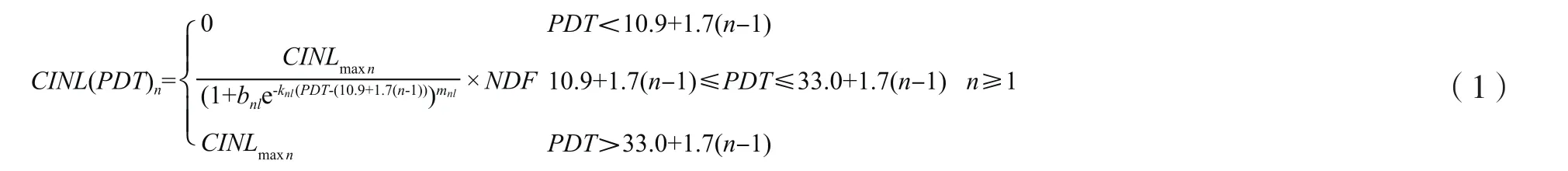
式(1)为节间伸长预测模型,PDT代表生理发育时间,CINL(PDT)n代表蓖麻主茎某一生理发育时间第n节节间长度(m),CINLmaxn代表蓖麻主茎第n节节间长度极值(m),为品种遗传参数;bnl、knl和mnl为模型参数;NDF为氮肥影响因子,计算方法见文献[19]。式(1)为3段函数:当PDT小于某一节间开始伸长的时间时,其节间长度为0;当PDT大于某一节间停止伸长的时间时,其节间长度为节间长度极值CINLmaxn;当PDT处于某一节间开始伸长和停止伸长时间范围内时,其节间长度可用Richards方程计算,并受氮肥因子影响。
本试验观测结果表明,随着蓖麻主茎节间数由基部到顶部的递增,CINLmaxn呈逐渐增加趋势,第1节间长度极值最小,顶部节间长度极值最大,第1节间至第5节间节间长度极值增加缓慢,第5节间至第11节间的节间长度极值增加较快,第12节间之后,节间长度极值增加又趋于缓慢(图1)。因此,蓖麻主茎各节间长度极值可用式(2)计算。
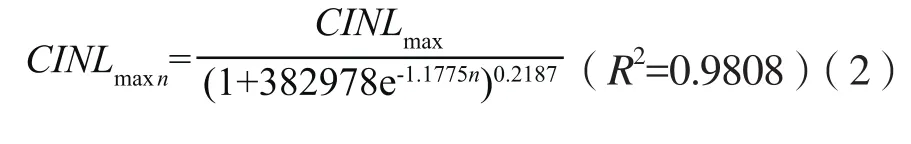

图1 节间长度极值变化Fig.1 Internode length variation
模型参数bnl与CINLmaxn呈指数函数关系,可通过式(3)计算。

模型参数knl随CINLmaxn呈“S”曲线变化,可用Logistic方程表达,见式(4)。

mnl与CINLmaxn亦呈指数函数关系,可通过式(5)计算。

2.1.2 各节直径预测模型 根据文献[19]的计算方法,运用本试验的观测数据计算可知,蓖麻主茎各节直径开始变粗的时间间隔为1.7个生理日,从开始变粗到粗度固定时间跨度为22.1个生理日。各节直径变化动态由式(6)计算:

式(6)中,CINT(PDT)n代表蓖麻主茎某一生理发育时间第n节直径(m),CINTmaxn代表蓖麻主茎第n节直径极值,是品种遗传参数。bnt、knt和mnt为模型参数。
随着蓖麻主茎节间数由基部到顶部递增,CINTmaxn呈逐渐降低的趋势,第1节直径极值最大,最顶部节直径极值最小(图2)。各节直径极值用式(7)计算。

图2 各节直径极值变化Fig.2 Change in diameter maximum of each section

式(7)中,CINTmax为顶部节间极值,是品种遗传参数。式(7)表明,某一节间极值可以根据顶部节间极值,运用Richards方程来计算。
bnt、knt和mnt等模型参数分别是CINTmaxn的指数函数、线性函数和对数函数关系,见式(8)、(9)和(10)。
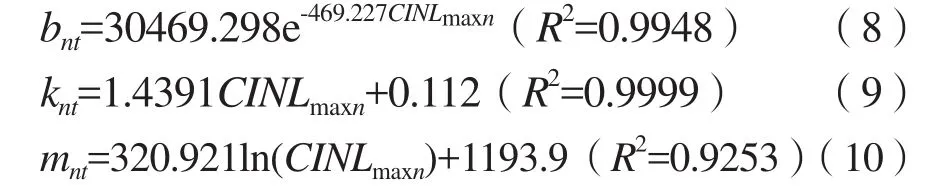
2.2 叶片扩展的预测模型
2.2.1 叶片伸长的动态模型 叶片伸长动态见图3,其增长动态可用式(11)表达。

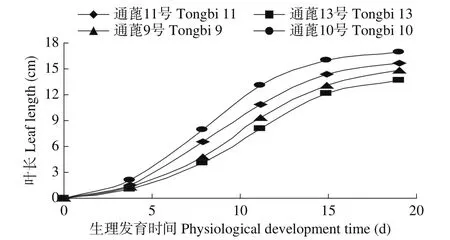
图3 叶长极值的动态Fig.3 Dynamic of the maximum of leaf length
式中,CLLEN(PDT)代表某一生理发育时间蓖麻主茎叶长(m),CLLENmax代表蓖麻主茎第3复叶的长度极值(m),定为品种遗传参数。根据文献[19]的计算方法,运用本试验的观测数据计算可知,第3复叶出现的生理发育时间是18.55个生理日,停止生长的生理发育时间是37.56个生理日。式(11)表明,在18.55~37.56个生理日的时间范围内,蓖麻主茎第3复叶的长度可用Richards方程来计算,并受氮肥效应的影响。
2.2.2 叶片增宽的动态模型 叶片增宽动态见图4,叶片宽度极值在不同PDT阶段变化不同,当PDT<5或者PDT>15时,其增加较为缓慢,当5<PDT<15时,其增加的幅度较大。

图4 叶宽极值的动态Fig.4 Dynamic of the maximum of leaf width
这种变化趋势可用式(12)表达。
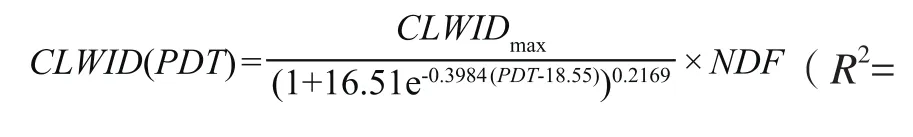

其中,CLWID(PDT)为某一生理发育时间蓖麻主茎第3复叶的宽度(m),CLWIDmax表示蓖麻主茎第3复叶的宽度极值(m),可定为品种遗传参数。式(12)表明,在18.55~37.56个生理日的时间范围内,蓖麻主茎第3复叶的宽度可用Richards方程来计算,并受氮肥效应的影响。
2.3 果穗生长的预测模型
蓖麻主茎果穗长变化的动态见图5,主茎果穗长在前期(PDT≤15)增加幅度不大,后期增加幅度较大,可用式(13)表达。

图5 穗长的生长动态Fig.5 Dynamic of ear length

式中,CBLEN(PDT)为某一生理发育时间蓖麻主茎果穗长(m),CBLENmax为蓖麻主茎果穗长极值(m),可定义为品种遗传参数。根据文献[19]的计算方法,运用本试验的观测数据计算可知,主茎果穗开始生长的生理发育时间是33.48个生理日,停止生长的生理发育时间是57.41个生理日。式(13)表明,在33.48~57.41个生理日的时间范围内,蓖麻主茎第3复叶的宽度可用Richards方程来计算,并受氮肥效应的影响。
2.4 蒴果生长的预测模型
2.4.1 蒴果长变化的动态模型 蓖麻蒴果长动态变化(图6)显示,其增加的时期主要在生育前期(PDT<15),生育后期几乎没有变化,可用方程(14)表示:


图6 蒴果长极值的动态Fig.6 Dynamic of the maximum of capsule length
式中,CBWID(PDT)为某一生理发育时间蓖麻主茎蒴果长(m),CBWIDmax为蒴果长极值(m),是品种遗传参数。根据文献[19]的计算方法,运用本试验的观测数据计算可知,蒴果开始增长的生理发育时间是33.48个生理日,停止增长的生理发育时间是57.41个生理日。式(14)表明,在33.48~57.41个生理日的时间范围内,蓖麻蒴果长变化呈不对称“S”曲线,分渐增、激增和缓增3个阶段,可用Richards方程来计算,并受氮肥效应的影响。
2.4.2 蒴果直径变化的动态模型 蒴果直径动态变化见图7,可用式(15)表达。

图7 蒴果直径极值的动态Fig.7 Dynamic of the maximum of capsule diameter

式中,CBTHI(PDT)为某一生理发育时间蓖麻蒴果直径(m),CBTHImax为蓖麻蒴果直径极值(m),是品种遗传参数。根据文献[19]的计算方法,运用本试验的观测数据计算可知,蒴果开始增粗的生理发育时间是33.48个生理日,停止增粗的生理发育时间是57.41个生理日。
2.5 品种遗传参数的确定及模型的检验
2.5.1 品种遗传参数的确定 表1显示,4个供试蓖麻品种遗传参数中,顶部节间长在18.68~26.52cm,第1节间直径在2.96~3.38cm,叶长在13.72~17.00cm,叶宽在12.32~14.04cm,果穗长在44.00~56.84cm,蒴果长在2.30~2.54cm,蒴果直径在1.90~2.04cm。通蓖10号各项参数值最高,通蓖13号最低。
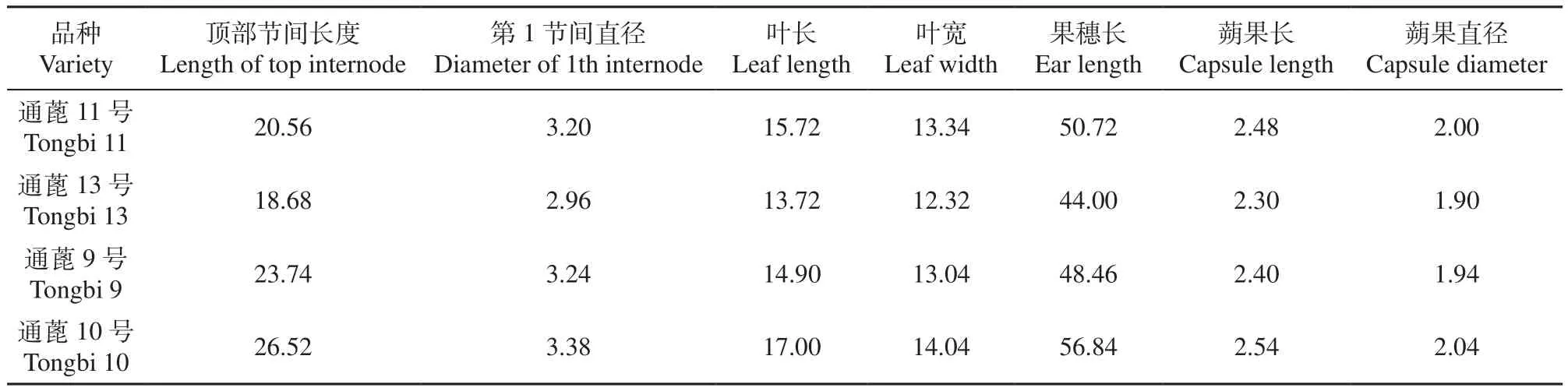
表1 4个蓖麻品种遗传参数Table 1 Genetic parameters of four castor varieties cm
2.5.2 模型的检验 选择通辽市农业科学研究院试验点现蕾期的观测数据,采用根均方差RMSE方法[18]对蓖麻主茎器官形态建成的预测模型进行检验。

式(16)中,OBSi为观测值,SIMi为模拟值,n为样本容量。
选择各品种蓖麻主茎第3、5、7和9节间,对其长度和直径预测模型进行检验,结果(表2)显示,所检验节间长度预测绝对误差在0.10~0.72cm,RMSE在0.14~0.60cm;各节直径的模拟误差在0.03~0.16cm,RMSE在0.06~0.11cm。

表2 节间长和主茎直径预测误差(n=16)Table 2 Prediction errors of internode length and main stem diameter on castor (n=16) cm
选择苗期和灌浆期的观测数据对蓖麻主茎第3复叶的生长动态模型和果穗长、蒴果长和蒴果直径生长动态模型进行检验,结果(表3)显示,叶长的预测误差在0.19~0.73cm,RMSE为0.47cm;叶宽的预测误差在0.30~0.60cm,RMSE为0.38cm;果穗长的模拟误差在0.89~1.85cm,RMSE为1.42cm;蒴果长的模拟误差在0.11~0.21cm,RMSE为0.16cm;蒴果直径的模拟误差在0.05~0.12cm,RMSE为0.07cm。

表3 蓖麻叶长、叶宽、果穗长度、蒴果长和蒴果直径预测误差(n=20)Table 3 Prediction errors of leaf length, leaf width, ear length, capsule length and capsule diameter of castor (n=20) cm
运用各品种蓖麻主茎茎秆长度、粗度、叶长、叶宽、果穗长、蒴果长和蒴果直径的观测值和模拟值数据,绘制两者之间的1∶1关系图(图8),各观测点均集中在1∶1直线附近,表明模拟值与观测值间误差较小,显示模型具有一定的可靠性。

图8 蓖麻主茎茎秆、叶片及果实观测值与模拟值比较Fig.8 Comparison of simulated values with observed values on stem, leaf and fruit of castor
3 讨论
3.1 关于模型的建模思路
目前,作物器官形态建成模拟模型的构建有以下4种思路:一是以积温(生长度日,GDD)为驱动变量,构建积温与器官生长的动态关系。如周娟等[21]构建了棉花主茎叶长和叶宽、主茎叶柄长、主茎节间长和粗、果枝叶长和宽、果枝叶柄长、果节长和粗以及棉铃高度和直径等随GDD变化的动态模型;孙爱珍等[22]构建了水稻叶片几何形态参数与有效积温之间的动态关系模型。二是以辐热积为驱动变量,构建其与器官生长的动态关系,这类模型多见于蔬菜和鲜花等温室作物。如常毅博等[23]建立了番茄株高、单株展开叶数及单叶面积随辐热积的变化模型;倪纪恒等[24]构建了基于辐热积的黄瓜单果果长、果径、开花节位的动态变化模型;李刚等[25]构建了温室切花百合出叶数、株高、第1花蕾长度和第1花蕾直径随幅热积的动态变化模型。三是以生物量为驱动变量,构建其与作物器官生长的动态关系,这是目前大田作物器官形态建成模型中应用最多的一类,通过系统观测作物器官生物量、体积、密度和直径等指标数据与器官生长的动态趋势来建立模型。如宋有洪等[26]构建了基于器官生物量的植株形态的玉米虚拟模型;陈超等[27]在系统分析生物量增加与器官增长关系的基础上,构建了棉花叶片和棉铃生长动态模型;展志岗等[28]描述了冬小麦叶片、茎秆和穗等器官随其生物量变化的动态方程;张伟欣等[29]构建了基于生物量的油菜越冬前植株叶片空间形态结构模型。四是以生理发育时间为驱动变量,构建其与作物器官生长的动态关系。如徐寿军等[30-31]以生理发育时间为模型尺度,构建了大麦和大豆地上部主要器官形态建成预测模型;杨恒山等[32]构建了基于生理发育时间的玉米器官形态建成的模拟模型。
3.2 关于模型的机理性
本研究预测了常规栽培条件下蓖麻地上部叶片、茎秆和蒴果等器官的生长过程,检验结果表明所建模型具有较好的预测性和可靠性。与既有模型比较,本研究在模拟尺度确定和模型参数确定方面有所差异。以生理发育时间而非作物实际生长天数作为模拟尺度,消除了以实际天数作为模拟尺度造成的温光差异对蓖麻生育进程的影响,统一了各器官开始生长和结束生长的时间节点以及器官间叠次生长的时间间隔。在品种遗传参数确定方面,对不同器官采取了不同的处理方法。对于节间生长的模型参数处理较为系统,探明了各节间同伸规律,量化了各节间同步生长及其与生理发育时间的关系,找出了各节间长度和粗度的变化规律,确定了最优条件下各品种节间长度和粗度极值,在此基础上,以节间长极值和主茎直径极值作为品种遗传参数来计算其他模型参数,较好地体现了节间生长过程中的同步关系。对于叶片、果穗和蒴果的模型参数处理则相对简化,将叶长极值、叶宽极值、果穗长极值和蒴果直径极值直接确定为品种遗传参数,经验性和机理性有效地结合,使模型更加简便实用。
蓖麻分枝多,形态复杂,本研究构建了地上部主茎器官形态建成模型。如何描述分枝器官的生长动态,是下一步要进行的工作,模型也需更多品种和更大地域范围的检验。
4 结论
以生理发育时间为模拟尺度,构建了蓖麻地上部主茎器官形态建成的预测模型,检验结果表明,模型表现出较好的预测性、可靠性和一定的机理性。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29 08:55:54
安徽农业大学学报(2022年3期)2022-10-25 12:32:36
甘肃农业大学学报(2021年4期)2021-09-22 06:58:50
种子(2021年7期)2021-08-19 01:46:58
扬子江诗刊(2018年2期)2018-11-13 13:08:03
扬子江(2018年2期)2018-03-24 09:29:02
中国马铃薯(2017年1期)2017-03-02 09:15:46
创新作文(3-4年级)(2015年8期)2015-09-15 11:19:46
中国粮油学报(2015年5期)2015-02-06 01:47:26
作文周刊·小学二年级版(2013年9期)2013-04-29 00:44:03
