
紫外处理和热激处理调控芥菜芽苗主要生理生化及褪黑素富集研究
2021-04-19 03:27:52耿钰涵吴悦豪方维明杨正飞尹永祺
核农学报 2021年5期
耿钰涵 程 超 吴悦豪 方维明 杨正飞 尹永祺
(扬州大学食品科学与工程学院,江苏 扬州 225127)
褪黑素是一种胺类激素,具有改善睡眠、延缓衰老、抗击肿瘤及提高免疫力等多种生理功效[1-3]。目前,市面上以褪黑素为主要功能成分的保健品和膳食补充剂大多为化学合成的[4]。研究发现褪黑素广泛存在于植物组织中,且其含量与植物种类、生长阶段、组织器官及外界环境密切相关[5]。褪黑素生物合成活性在植物的幼嫩组织中较高,且植物在不同的环境条件下,尤其是在逆境条件下其体内褪黑素含量显著增加。Byeon 等[6]发现,将水稻幼苗置于55℃的黑条件下1 h,其褪黑素含量由2.95 ng·g-1提高到4.90 ng·g-1。绿藻经高温和重金属处理褪黑素含量有所升高,且外源性褪黑素的添加缓解了镉胁迫对藻类的影响[7]。向日葵在盐胁迫条件下,褪黑素含量上升[8]。因此,通过调控植物生长发育以促进其体内褪黑素积累,进而开发富含褪黑素的营养食品以补充人体所需的褪黑素是一种环保、绿色的保健方式,符合未来的发展方向。
芥菜(Brassica junceaL.)是十字花科芸薹属一年生或二年生草本植物,分布广泛,是我国重要的鲜食和加工蔬菜[9]。研究发现,相较于其他植物源食品原料,芥菜中褪黑素含量最高[10]。本研究通过前期试验发现,芥菜芽苗经紫外和热激处理可显著提高其褪黑素含量,但其作用机制尚不明确。此外,芥菜中硫代葡萄糖苷被催化水解得到的异硫氰酸酯具有抗氧化、杀菌、抗癌等多种生理功效[10-11]。在高等植物中,色氨酸经色氨酸脱羧酶(tryptophan decarboxylase,TDC)、色氨酸5-羟化酶(tryptophan 5-hydroxylase,T5H)、血清素N-乙酰转移酶(serotonin N-acetyltransferase,SNAT)、乙酰基血清素甲基转移酶(acetyl serotonin methyl transferase,ASMT)催化形成褪黑素[12]。研究紫外处理和热激处理下这些褪黑素关键合成酶的基因表达变化有助于分析褪黑素富集的作用机制。
1 材料与方法
1.1 材料与试剂
芥菜种子(长合甜脆大肉芥菜),广州长合种子有限公司,-20℃储存。乙腈(色谱纯),美国Sigma 公司;总抗氧化能力检测试剂盒(A015-1)、谷胱甘肽过氧化物酶检测试剂盒(A005-1),南京建成生物工程研究所;植物RNA 提取试剂盒(R6827),美国OMEGA公司;RNA 反转录试剂盒(RR047A)、TB Green Premix Dimer Eraser(RR091A),日本TaKaRa 公司;其余试剂均为国产分析纯。
1.2 仪器与设备
pGX-150 型智能光照培养箱,宁波海曙赛福实验仪器厂;DICO 型台式离心机,上海Thermo Fisher 有限公司;UV-7504C 型紫外可见分光光度计,上海欣茂仪器有限公司;Agilent 1200 液相色谱仪、荧光检测器系统,美国Agilent 科技有限公司;Stepone plus 荧光定量PCR,美国ABI 公司。
1.3 试验方法
1.3.1 样品制备 取芥菜种子10 g,按料液比1 ∶10(g·mL-1)置于1 %(v/v)次氯酸钠水溶液中浸泡消毒15 min,之后用蒸馏水冲洗pH 值至中性,然后置于30℃蒸馏水中浸泡4 h。浸泡后的芥菜种子置于铺有蛭石的小盒中,喷施60 mL 蒸馏水,分别设置以下处理组:
(1)对照(CK):置于30℃培养箱内发芽,每天16 h 光照/8 h 黑暗,每天喷施20 mL 去离子水。
(2)紫外(ultraviolet,UV)处理:置于30℃培养箱内发芽,每天16 h 光照/4 h 黑暗/4 h UV 照射,每天喷施20 mL 去离子水。
(3)热激(Heat)处理:置于30℃培养箱内发芽,每天16 h 光照/7.5 h 黑暗,并于每天13:30 放入50℃避光恒温箱30 min,每天喷施20 mL 去离子水。
芽苗共发芽7 d,于发芽4 d 和7 d 取样,液氮速冻后置于-20℃保存,测定相关指标。
陈少平:2019年,广东垦区上下将全面贯彻党的十九大和十九届二中、三中全会精神,以习近平新时代中国特色社会主义思想为指导,深入贯彻习近平总书记对农垦和广东重要讲话精神,高举新时代改革开放旗帜,抓住粤港澳大湾区建设这一大机遇、大文章,立足农垦定位,发挥垦区优势,持续推进“走出去”发展,让广东农垦的发展搭上新一轮改革发展的快车,为实施乡村振兴战略、质量兴农战略、健康中国战略做出广垦贡献。
1.3.2 指标测定
1.3.2.1 形态特征 各处理组随机选取芥菜芽苗,用相机拍摄其形态特征。
1.3.2.2 芽长 各处理组随机选取30 株芽苗,用游标卡尺测量其根尖到顶端的长度,取平均值。
1.3.2.3 过氧化氢含量 采用分光光度法测定[13]。
1.3.2.4 可溶性蛋白含量 采用考马斯亮蓝G250法测定[14],以牛血清白蛋白为标准。
1.3.2.5 总抗氧化能力 采用总抗氧化能力检测试剂盒测定。
1.3.2.6 谷胱甘肽过氧化物酶活性 采用谷胱甘肽过氧化物酶检测试剂盒测定。
1.3.2.7 异硫氰酸酯含量 参照Jiao 等[15]的方法测定。
1.3.2.8 褪黑素含量 参照Arnao 等[16]的方法,稍作修改。取0.2 g 芥菜芽苗组织切片(3~5 mm),不进行均质化处理,置于装有3 mL 氯仿∶甲醇(30 ∶1,v ∶v)的棕色小瓶中,于4℃恒温箱中振荡15 h。然后于12 000×g离心10 min,将下层清液避光真空干燥,再用100 μL 35%甲醇溶解,取20 μL 样本采用高效液相色谱法测定褪黑素含量。色谱柱:ZORBAX SB-C18 柱(5 μm 粒径,4.6 mm×250 mm);梯度洗脱(42%~50%甲醇梯度洗脱27 min,50%甲醇等体积洗脱);流速:0.3 mL·min-1; 分离时间:31 min;检测器温度:30℃。在280 nm 激发光和348 nm 发射光下测定褪黑素含量。
1.3.2.9 关键酶基因表达量 参照吴艳迪[17]的方法,获得褪黑素合成酶关键基因TDC、T5H、SNAT、ASMT的基因序列。根据NCBI 上报道的其他植物褪黑素合成酶的同源蛋白,并与芥菜基因组数据库(http:/ /brassicadb. org/brad/index. php) 比 较,作 为DNAMAN 软件的参考。选择具有保守功能域且与其他相应植物基因同源性超过30%的潜在同源基因作为候选基因。表1 为荧光实时定量聚合酶链式反应(quantitative real-time PCR,qRT-PCR)的引物序列。
1.4 数据统计与分析
试验设3 次重复,结果以平均值±标准差表示。采用SPSS 18.0 软件分析,采用Tukey 多重比较,在0.05 水平上进行显著检验,基因表达数据采用2-ΔΔCt法[18]处理。
2 结果与分析
2.1 芥菜芽苗形态特征、芽长和过氧化氢含量
由图1-A 可知,相较于CK,经紫外处理和热激处理的芥菜芽苗生长受到抑制,其叶片变小且芽长、根长均受到影响,而紫外和热激两处理间形态差异较小。由图1-B 可知,随着发芽时间的延长,紫外处理和热激处理的芥菜芽苗芽长呈增长趋势,两种处理的芽苗芽长均显著低于CK(P<0.05),但紫外处理和热激处理的芽苗芽长无显著差异(P>0.05)。由图1-C 可知,过氧化氢含量随发芽时间延长呈下降趋势。发芽7 d时,热激处理的芥菜芽苗过氧化氢含量较CK 显著下降(P<0.05),且紫外处理和热激处理后芽苗的过氧化氢含量均低于CK。表明紫外处理和热激处理抑制了芥菜芽苗的生长。

表1 qRT-PCR 的引物序列Table 1 Primers sequences used for qRT-PCR
2.2 可溶性蛋白含量
由图2 可知,发芽期间,紫外处理与热激处理的芥菜芽苗中可溶性蛋白含量显著高于CK(P<0.05),发芽4 d 时,热激处理的芽苗中可溶性蛋白含量较紫外处理也显著提高(P<0.05),此时,紫外处理与热激处理芥菜芽苗的可溶性蛋白含量分别是CK 的1.44 和1.95 倍,热激处理芥菜芽苗的可溶性蛋白含量是紫外处理的1.36 倍。说明紫外处理与热激处理可显著提高芥菜芽苗中可溶性蛋白的含量。
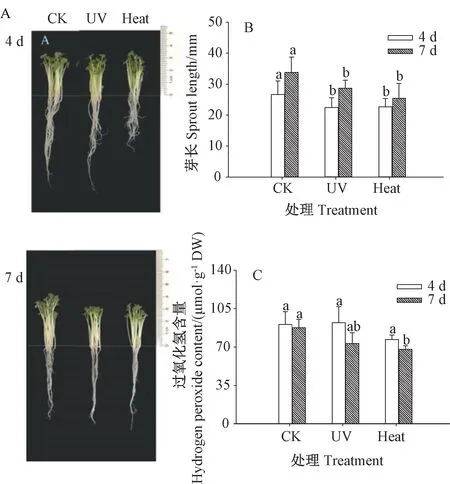
图1 紫外处理和热激处理对芥菜芽苗形态特征、芽长和过氧化氢含量的影响Fig.1 Effect of UV and heat stress treatments on morphological characterization,sprout length andhydrogen peroxide content of mustard sprouts
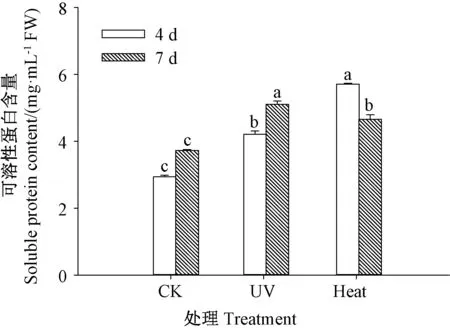
图2 紫外和热激处理对芥菜芽苗可溶性蛋白含量的影响Fig.2 Effect of UV and heat stress treatments on soluble protein content of mustard sprouts
2.3 总抗氧化能力
由图3 可知,发芽期间,紫外处理和热激处理的芥菜芽苗总抗氧化能力均显著高于CK(P<0.05)。发芽4 d 时,热激处理芥菜芽苗的总抗氧化能力与紫外处理差异不显著(P>0.05)。发芽7 d 时,热激处理芥菜芽苗的总抗氧化能力显著高于紫外处理(P<0.05)。表明芥菜芽苗经紫外处理和热激处理后总抗氧化能力显著提高。
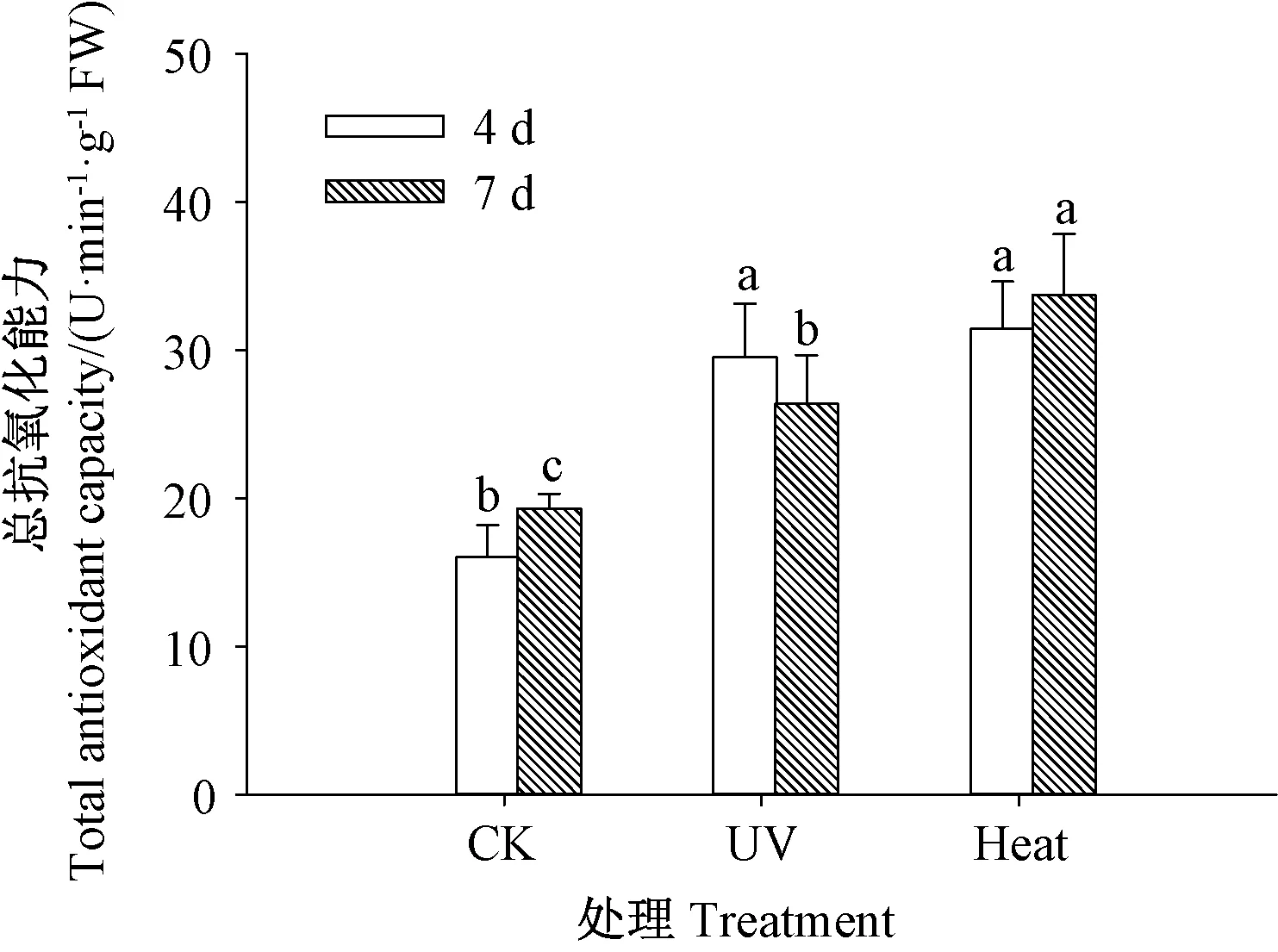
图3 紫外和热激处理对芥菜芽苗总抗氧化能力的影响Fig.3 Effect of UV and heat stress on total antioxidant capacity of mustard sprouts
2.4 谷胱甘肽过氧化物酶活性
由图4 可知,发芽4 d 时,热激处理芥菜芽苗的谷胱甘肽过氧化物酶活性显著高于紫外处理(P<0.05),紫外处理芥菜芽苗的谷胱甘肽过氧化物酶活性与CK间差异不显著(P>0.05)。发芽7 d 时,紫外处理和热激处理芥菜芽苗的谷胱甘肽过氧化物酶活性均显著高于CK(P<0.05)。表明芥菜芽苗经紫外和热激胁迫作用后,谷胱甘肽过氧化物酶活性提高。
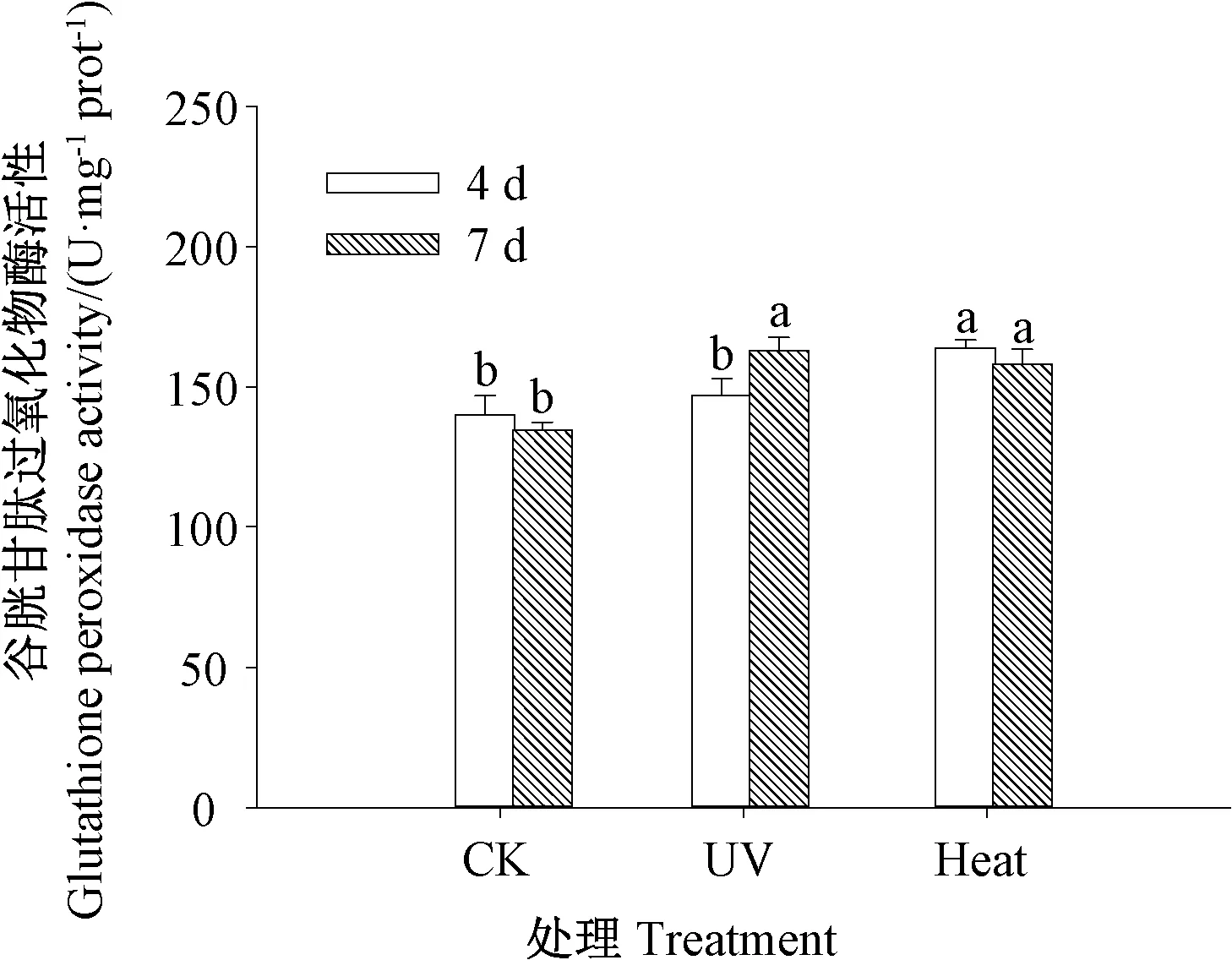
图4 紫外和热激处理对芥菜芽苗谷胱甘肽过氧化物酶活性的影响Fig.4 Effect of UV and heat stress treatment on glutathione peroxidase activity of mustard sprouts
2.5 异硫氰酸酯含量
由图5 可知,随发芽时间的延长,各处理芥菜芽苗的异硫氰酸酯含量均下降。发芽4 d 时,各处理芥菜芽苗的异硫氰酸酯含量无显著差异(P>0.05)。发芽7 d 时,热激处理芥菜芽苗的异硫氰酸酯含量较CK 显著提高(P<0.05),是CK 的1.78 倍。说明热激处理可提高芥菜芽苗中异硫氰酸酯的含量。
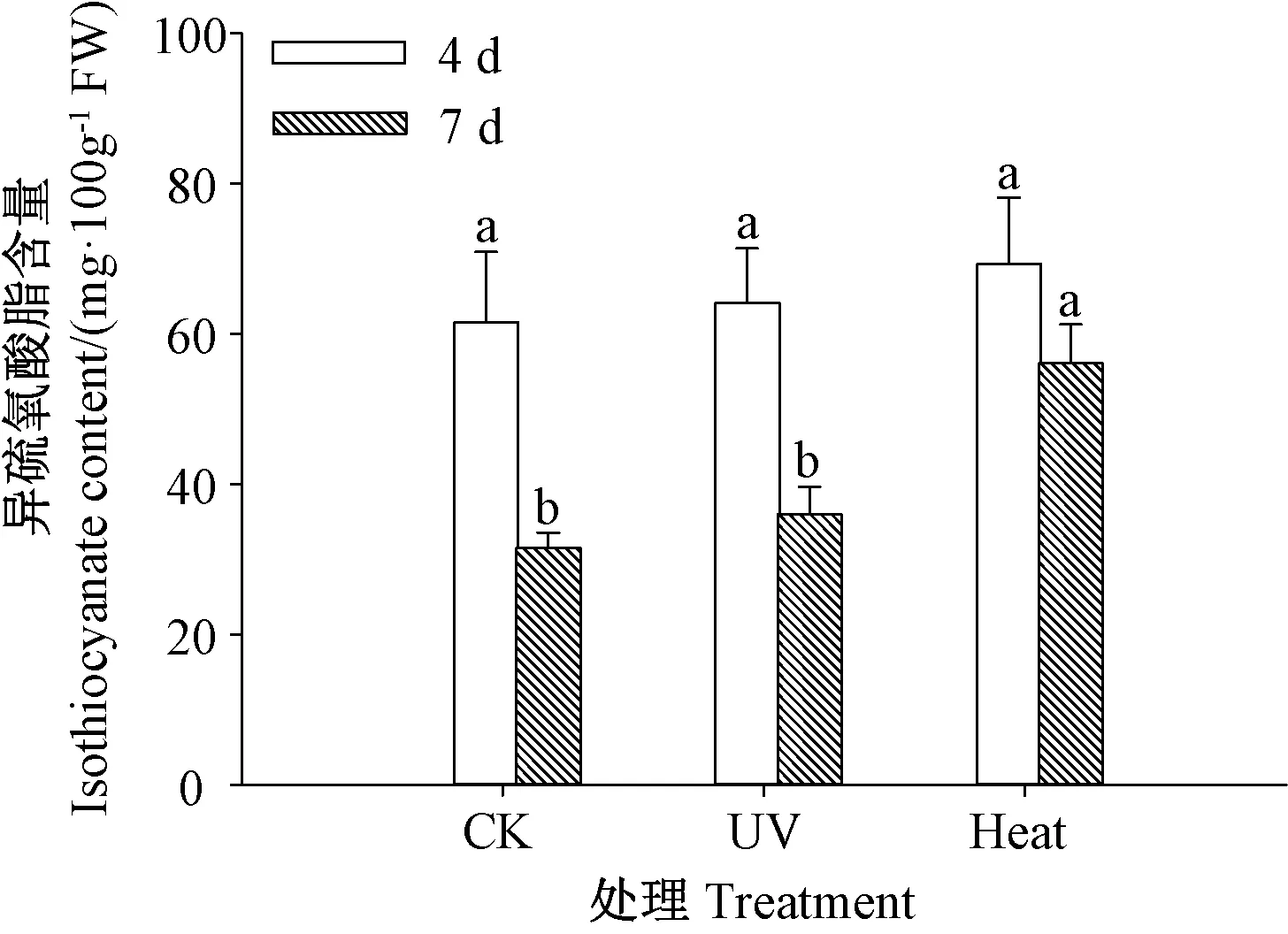
图5 紫外和热激处理对芥菜芽苗异硫氰酸酯含量的影响Fig.5 Effect of UV and heat stress treatments on isothiocyanate content of mustard sprouts
2.6 褪黑素含量
由图6 可知,发芽4 d 时,紫外处理芥菜芽苗的褪黑素含量与CK 无显著差异(P>0.05),紫外处理与热激处理芥菜芽苗的褪黑素含量分别是CK 的1.17 和2.89 倍,热激处理芥菜芽苗的褪黑素含量为紫外处理的2.47 倍。发芽7 d 时,紫外处理和热激处理芥菜芽苗的褪黑素含量均显著高于CK(P<0.05),分别是CK的2.16 和1.92 倍。表明热激处理4 和7 d、紫外处理7 d 可显著提高芥菜芽苗的褪黑素含量,且发芽4 d 时芥菜芽苗的褪黑素含量最高(65 ng·g-1FW)。
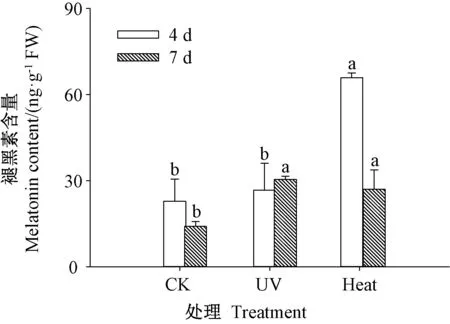
图6 紫外和热激处理对芥菜芽苗褪黑素含量的影响Fig.6 Effect of UV and heat stress treatments on melatonin content of mustard sprouts
2.7 关键酶基因表达量
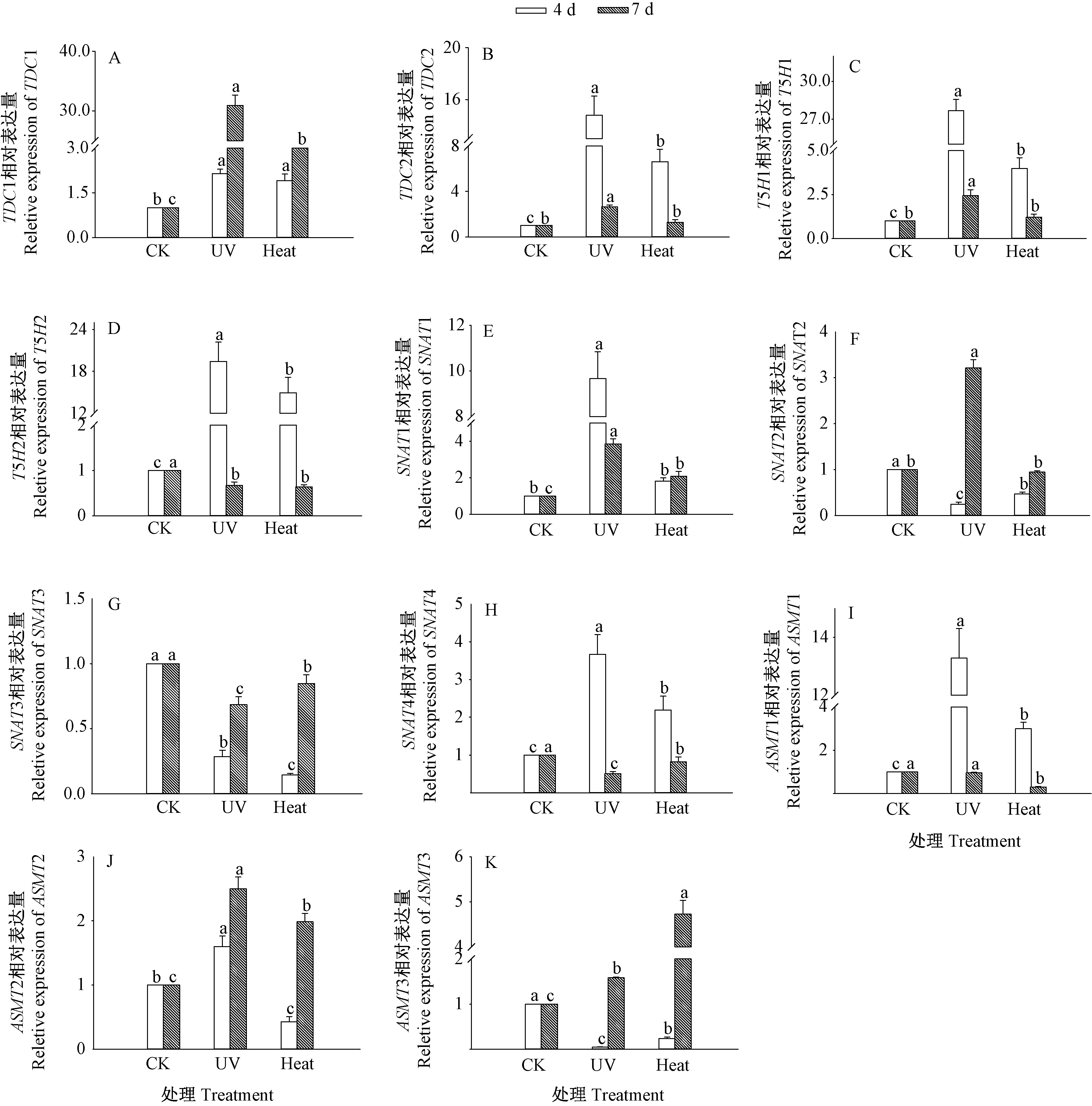
图7 紫外和热激处理对芥菜芽苗关键酶基因表达的影响Fig.7 Effects of UV and heat stress treatments on the expression of key enzyme gene in mustard shoots
由图7-A 可知,与CK 相比,紫外处理和热激处理显著提高了芥菜芽苗TDC1 的表达量(P<0.05),但发芽4 d 时紫外处理和热激处理芥菜芽苗TDC1 的表达量无显著差异。由图7-B 可知,发芽4 d 时,紫外处理与热激处理芥菜芽苗TDC2 的表达量分别为CK 的14.80 和6.61 倍。与CK 相比,经紫外处理和热激处理发芽4 d 芽苗中T5H1 和T5H2 表达量均显著上调(图7-C、D)。发芽4 d 时,芽苗中SNAT1 表达量经紫外处理显著上调,但热激处理对其无显著影响(图7-E)。发芽4 d 时,热激处理芥菜芽苗SNAT2 的表达量显著低于CK(P<0.05)(图7-F)。紫外处理和热激处理使SNAT3 的表达量显著下调(P<0.05)(图7-G)。发芽4 d 时,两种处理芥菜芽苗SNAT4 的表达量分别为CK 的3.66 和2.19 倍(图7-H)。发芽4 d 时,紫外处理和热激处理芥菜芽苗ASMT1 的表达显著上调(图7-I)。发芽4 d 时,热激处理芥菜芽苗ASMT2 的表达量相较紫外处理显著下调(图7-J),而ASMT3 的表达量显著上调(图7-K)。表明紫外处理和热激处理可通过影响芥菜芽苗中褪黑素合成关键酶基因的表达,进而调控芥菜芽苗中褪黑素的合成。
3 讨论
植物在盐[19]、高温[20]、干旱[21]、紫外[22]等胁迫作用下,细胞会产生大量自由基,造成膜脂过氧化损伤,影响植物的生长与发育[23]。本研究发现,芥菜芽苗经紫外处理和热激处理后,植株生长形态较小,芽长降低,表明芥菜芽苗生长受到抑制。植物器官衰老或在逆境下遭受伤害,过氧化氢与可溶性蛋白含量的变化是对逆境做出的有效生理反应,激活内源抗氧化防护系统,从而减轻或缓解逆境伤害。总抗氧化能力是反映机体抗氧化作用的综合性指标[24-26]。发芽期间,紫外处理和热激处理芥菜芽苗的总抗氧化能力和可溶性蛋白含量均显著高于CK,处理7 d 时,热激处理芥菜芽苗的可溶性蛋白含量显著高于紫外处理,这与经干旱与聚乙二醇胁迫处理的小麦幼苗中相关指标结果一致[27]。研究发现,谷胱甘肽可通过巯基与自由基结合,将自由基转化成易代谢的酸类物质,从而加速自由基代谢,因此谷胱甘肽过氧化物酶活性也是判断芽苗受胁迫程度的重要指标[28]。发芽7 d 时,激处理和紫外处理芥菜芽苗的谷胱甘肽过氧化物酶活性均显著高于CK,表明紫外处理和热激处理会抑制芥菜芽苗的生长,并可能通过自身的调控作用应对外界环境的胁迫。
Tan 等[29]发 现,在强中波紫外线(ultraviolet radiation B,UV-B)辐射条件下,水葫芦的褪黑素含量显著提高。Shi 等[30]研究表明,热胁迫处理可显著提高拟南芥叶片的褪黑素含量。本研究发现,发芽期间,热激处理4 和7 d、紫外处理7 d 时芥菜芽苗的褪黑素含量均显著高于CK,且发芽4 d 时芥菜芽苗中褪黑素含量最高;另外,紫外处理和热激处理提高了芥菜芽苗中异硫氰酸酯的含量,且热激处理的效果更明显。表明紫外处理和热激处理可富集芥菜芽苗中的褪黑素,同时提高芽苗中异硫氰酸酯的含量,进而提升功能性食品的品质[31]。植物体内的褪黑素是由色氨酸经色氨酸脱羧酶(tryptophan decarboxylase,TDC)的催化转化为色胺,然后经过色氨酸5-羟化酶(tryptophan 5-hydroxylase,T5H)催化色胺形成5-羟色胺,再由血清素N - 乙酰转移酶(serotonin N-acetyltransterase,SNAT)催化5-羟色胺转化为N-乙酰基-5-羟色胺,最后经乙酰基血清素甲基转移酶(hydroxyindole-Omethyltransferase,ASMT)或羟基吲哚-O-甲基转移酶(hydroxyindole-O-methyltransferase,HOMT)作用将N-乙酰基-5-羟色胺形成褪黑素[8]。不同观察时间点(4 d 和7 d),褪黑素合成关键酶基因表达量变化趋势并不相同,这与不同关键酶在不同阶段发挥作用有关。本研究发现,与CK 相比,TDC的表达量升高时褪黑素含量也升高,说明其可以促进褪黑素的合成,这与Zhao 等[32]的研究结果一致。表明紫外处理和热激处理可显著影响褪黑素合成过程中关键酶基因的表达,从而提高芥菜芽苗中褪黑素的含量,实现褪黑素的富集。
4 结论
本研究发现,芥菜芽苗发芽期间,紫外处理和热激处理抑制了芥菜芽苗的生长发育,芽苗中过氧化氢含量降低,可溶性蛋白质含量、谷胱甘肽过氧化物酶活性和总抗氧化能力提高,异硫氰酸酯含量和褪黑素含量上升。在观察时间点(发芽4 d 和7 d),热激处理4 d时芥菜芽苗中褪黑素含量最高,为65 ng·g-1FW,说明热激处理是富集褪黑素的有效方式。
猜你喜欢
化学与粘合(2023年1期)2023-04-06 19:01:45
中国土壤与肥料(2021年5期)2021-12-02 01:05:40
广州化工(2021年5期)2021-03-17 01:34:16
现代园艺(2017年21期)2018-01-03 06:41:56
青年文学家(2017年28期)2017-11-28 15:41:54
分析科学学报(2016年6期)2016-10-16 01:17:40
中国塑料(2016年4期)2016-06-27 06:33:38
福建林业科技(2015年3期)2015-09-16 02:51:57
少儿科学周刊·儿童版(2014年1期)2014-03-10 19:55:19
快乐作文·中年级(2013年7期)2013-04-29 00:44:03
