
提高供镁水平对西洋参幼苗镉胁迫缓解作用研究
2021-04-13 09:21钱佳奇吴晨阮音音吴虎平孙海邵财张亚玉
特产研究 2021年2期
钱佳奇,吴晨,阮音音,吴虎平,孙海,邵财,张亚玉,2※
(1 中国农业科学院特产研究所,吉林 长春 130112;2 成都大学药学与生物工程学院,四川 成都 610106)
镉(Cd)是毒性最强、农田污染最严重的重金属之一[1],Cd胁迫会导致植物体内过多活性氧(如O2-、H2O2等)的产生与积累,从而引发或者加剧细胞膜脂过氧化、蛋白质变性以及核苷酸受损等,严重时导致细胞死亡[2]。土壤中的镉有着很强的生物迁移性,土壤镉污染已对我国农产品的安全及人体健康构成潜在威胁。
镁(Mg)为植物生长必需元素之一,也是叶绿素的重要组分[3]。镁离子(Mg2+)也是许多酶的激活剂或组分,尤其是转移磷酸基酶的活化剂,其所活化的酶类关系到糖类、脂肪、蛋白质和核酸等的物质代谢与能量转化[4],在植物的光合作用和植物抗逆过程中发挥着重要作用,近年来的研究表明,镁可以减轻重金属对植物的毒害作用,但关于镁是否能缓解镉对植物的毒害作用及其生理作用机制还不清楚。
西洋参(Panax quenquifolium L.)又称西洋人参、花旗参等,属五加科人参属名贵药材,以根部入药,有抗疲劳、提高免疫、清热解毒、滋阴补肾和口齿生津的功效[5],被医药及保健行业广泛应用。就国产西洋参而言,目前并未发现镉含量超标现象[6],但在西洋参主要栽培区的部分散户依然进行着过量使用农药、化肥等不规范的生产方式,这给西洋参及其土壤安全带来了极大的隐患,目前依然存在西洋参产品安全及镉超标的潜在风险[7]。而国内外对镉胁迫下提高供镁水平对西洋参生理特性及镉积累的影响尚未报道。鉴于此,通过水培试验,研究镉胁迫条件下提高供镁水平对西洋参幼苗的缓解作用,为探索缓解植物镉胁迫的途径提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为健康无病、大小一致的1 年生西洋参幼苗。
1.2 试验设计与方法
试验开始之前设置两个预实验确定西洋参幼苗最适镉胁迫浓度及最适镁培养浓度:1、设置0、10、20、30、40、50mol L-1的CdCl2,培养12 d,最终结果表明30mol L-1的CdCl2 胁迫效果最佳(即胁迫症状较10、20mol L-1更明显,生长状态更差,叶片萎蔫更严重,植株发育更迟缓;40、50mol L-1的胁迫浓度幼苗死亡率较高,无法完成试验的样品采集);2、设置0、1、2、4、8、16 mmol L-1的Mg2+浓度,通过衡量生长及生理指标,最终确定Mg2+的最佳培养浓度为2 mmol L-1。
试验于2020 年7 月12 日至2020 年7 月24 日在中国农业科学院特产研究所智能光照培养间内进行。温度范围在23~28 ℃,湿度控制在60%~70% 之间,光照强度光强为105mol m-2s-1,光谱信息见图1。将西洋参种子经多菌灵处理后栽培至营养土中,萌发35 d 后(株高为5~6 cm),于2020 年7 月12 日用直径为2 cm、高为2.5 cm的圆柱状海绵夹住茎部插入预先准备好的带孔(直径1.7 cm)的塑料板(59 cm 11 cm)上,每板插苗30 株,将其放入盛有不同镁浓度的改良Hoagland 营养液的水培箱(60 cm 12 cm 20 cm)中,水培箱内含循环系统,潜水泵(Atman 创星AT-301)提供动力完成水培箱内营养液循环,以提高营养液中溶解氧含量,保障在试验过程中西洋参根系溶解氧的供应。每箱盛有7 L 营养液,开始胁迫处理。营养液配方pH 值为6.0。在此基础上加施的并设置Mg2+浓度为2、3、4、5 mmol L-1,记为Cd、CM1、CM2、M3,另设置Mg2+浓度为2 mmol L-1但不加CdCl2 记为CK,共5 组,见表1,每组3 次重复,共15 箱。当Mg2+浓度大于2 mmol L-1时,用MgCl2来增加Mg2+的供应,为防止盐分积累,每3 d 更换1 次营养液。为探究镉含量的动态变化,每3 d 进行1 次破坏性取样,将根、茎、叶分离,105℃杀青30 min,75 ℃烘干至恒重备用,其余各项指标在培养12 d 后进行测定:保护性酶样品采集后分装,液氮速冻,80 ℃冰箱保存;根系活力、生长指标及荧光参数均在取样当天完成。
图1 培养所用LED 灯的光谱信息Fig.1 Spectral information of LED lamps for culture
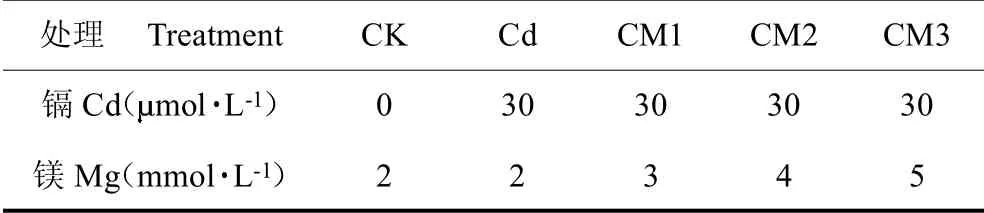
表1 试验设计Table 1 Design of this experiment
1.3 生长指标的测定
株高、茎粗分别用直尺及游标卡尺测量,叶面积使用叶面积仪YMJ-D(托普云农)进行测量,均6 次重复;每组选取10 株长势一致的植株,根用蒸馏水冲洗5 min,将根、茎、叶分离,分别测量其鲜重,后105 ℃杀青30 min,75 ℃烘干6 h 至恒重,使用精确度为0.000 1的电子天平称量其干重,备用。
1.4 根系活力的测定
根系活力采用TTC 还原法测定,用TTC 还原量表示根系活力[8]。
1.5 镉含量的测定
样品烘干后进行研磨,过100 目筛,准确称量0.100 0 g 样品,使用浓HNO3-HCIO4 法进行消解后上机(ICP-OES)检测[9]。
1.6 保护性酶的测定
破坏性取样,将每组叶片去掉主叶脉,使用手术刀将叶片切成正方形小块,混匀,准确称量0.1 g 叶片,液氮速冻,80 ℃冰箱保存,按照试剂盒(苏州科铭)说明书操作进行测定。
1.7 叶绿素含量的测定
选取大小一致的叶片,擦拭干净,使用取样器取样后剪成细丝,混匀并称取0.2 g,采用分光光度计法测定叶绿素含量[10]。
1.8 荧光参数的测定
使用超便携式调制叶绿素荧光MINI-PAM(WALZ,Effeltrich,Germany),用叶夹暗适应30 min后,PAR设定为10mol m-2s-1。测定光系统II (PSII)最大量子产额Fv/Fm=(Fm Fo)/Fm 及相对电子传递速率ETR。利用内置自动光源测定荧光诱导曲线,计算PSII实际量子产量Y(II)=(Fm' Fs)/Fm',Y(NPQ)=F/Fm' F/Fm。式中,Fm'、Fs 分别为光适应过程中的饱和荧光值和实时荧光值,Fm 和Fo 分别为暗适应后最大荧光值和最小荧光值[11]。
2 结果与分析
2.1 提高供镁水平对镉胁迫下西洋参幼苗生长的影响

图2 提高供镁水平对镉胁迫下西洋参幼苗生长的影响(平均值±标准偏差)。Fig.2 Effect of increased magnesium level on the growth of Ginseng seedlings under cadmium stress(mean±SD).
由图2 可知,镉胁迫下西洋参幼苗的生长受到显著抑制,株高、茎粗和叶面积分别较CK降低了10.03%、8.69%和3.86%,镉胁迫对株高的抑制最明显。在此基础上提高供镁水平可不同程度地缓解镉胁迫。其中CM2 组的缓解效应较Cd 组佳。其株高和茎粗分别较Cd 增加了9.41%和9.38%,差异显著,叶面积虽然增加了4.35%,但差异不显著。
2.2 提高供镁水平对镉胁迫下西洋参幼苗根系活力的影响
由图3 可知,镉胁迫下西洋参幼苗根系活力受到显著抑制,较CK 降低了17.50%。随着供镁水平的增加,根系活力逐渐增加,分别较Cd 增加了7.89%、23.12%和23.67%,当Mg2+为5 mmol L-1时增幅最显著,但与Mg2+为4 mmol L-1时差异并不显著。
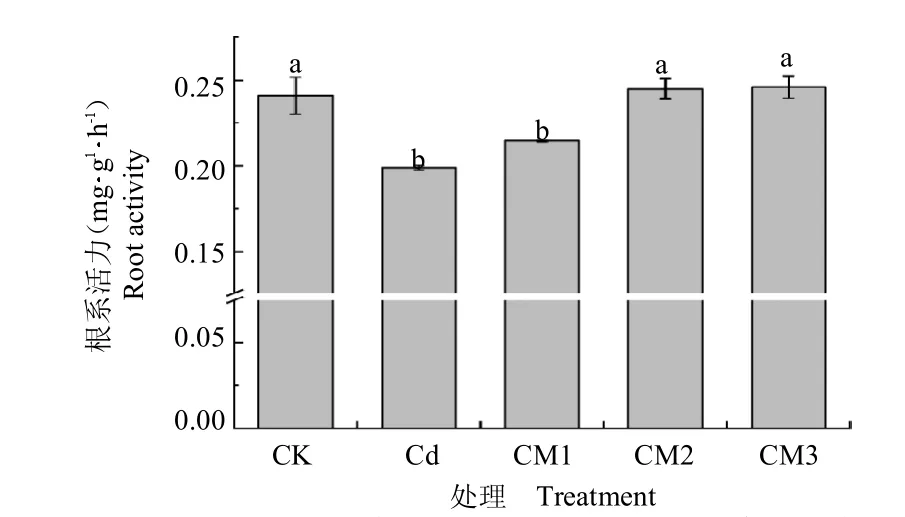
图3 提高供镁水平对镉胁迫下西洋参幼苗根系活力的影响(平均值±标准偏差)Fig.3 Effect of increased magnesium supply level on root activity of Panax quinquefolium seedlings under cadmium stress(mean±SD).
2.3 提高供镁水平对镉胁迫下西洋参幼苗镉元素积累的影响
由图4 可知,随着胁迫天数的增加,各实验组各部位的镉含量呈增加趋势;且各实验组各部位的镉含量呈现出Cd >CM1 >CM2 >CM3 >CK 的趋势;同时出现根中镉含量>茎中镉含量>叶中镉含量的现象。
2.4 提高供镁水平对镉胁迫下西洋参幼苗叶片保护性酶活性的影响
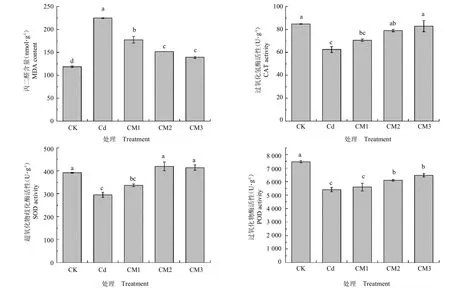
图5 提高供镁水平对镉胁迫下西洋参幼苗叶片保护性酶活性的影响(平均值±标准偏差)Fig.5 Effect of increased magnesium supply on the protective enzyme activity of panax quinquefolium seedling leaves under cadmium stress(mean±SD)
由图5 可知,在镉胁迫条件下,西洋参叶片保护性酶CAT、SOD 和POD 活性均显著降低,较CK 分别降低了26.39%、24.60%和27.64%,膜脂过氧化产物MDA含量呈显著升高的趋势,较CK升高了89.45%。随着供镁水平的提升,保护性酶活性有所升高,其中CAT 与POD 在Mg2+浓度为5 mmol L-1时增幅最显著,较Cd 增加了35.54%和19.77%,SOD 在Mg2+浓度为4 mmol L-1时增幅最显著,较Cd增加了42.00%,膜脂过氧化产物丙二醛(MDA)含量在Mg2+浓度为4 mmol L-1时降幅最显著,较Cd 降低了38.06%。但实验组CM2 与CM3 的各保护性酶活性及膜脂过氧化产物含量差异不显著。
2.5 提高供镁水平对镉胁迫下西洋参幼苗叶绿素含量的影响
由图6 可知,镉胁迫可显著降低西洋参幼苗叶绿素a 及叶绿素a+b 的含量,分别较CK 降低了12.03%和10.23%;叶绿素b 含量虽然有所降低,但各组之间差异不显著。随着供镁水平的提高,叶绿素a 及叶绿素a+b 的含量也随之升高,但与Cd 差异不显著,且CM2、CM3 与CK 差异也不显著。
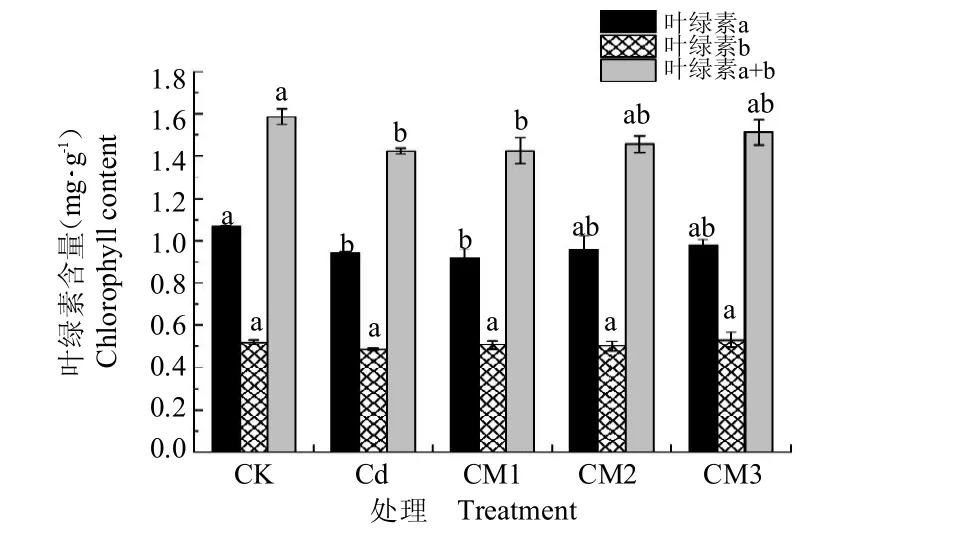
图6 提高供镁水平对镉胁迫下西洋参幼苗叶绿素含量的影响(平均值±标准偏差)Fig.6 Influence of increased magnesium supply level on chlorophyll content of Panax quinquefolium seedlings under cadmium stress(mean±SD)
2.6 提高供镁水平对镉胁迫下西洋参幼苗荧光参数的影响

图7 提高供镁水平对镉胁迫下西洋参幼苗荧光参数的影响(平均值±标准偏差)Fig.7 Influence of increased magnesium supply level on fluorescence parameters of Panax quaeneas seedlings under cadmium stress(mean±SD)
由图7 可知,镉胁迫下Fv/Fm、Y(II)和ETR 参数显著降低,分别较CK 降低了21.82%、33.92%和34.82%,Y(NPQ)显著上升,较CK 升高了38.20%。随着供镁水平的提高,Fv/Fm、Y(II)和ETR 均呈上升趋势,在Mg2+浓度为5 mmol L-1时增幅最显著,分别较Cd 升高了18.02%、14.20%和18.42%;随着供镁水平的提高Y(NPQ)呈降低趋势,在Mg2+浓度为5 mmol L-1时降幅最显著,较Cd 降低了23.11%。但实验组CM2 与CM3 的各参数差异不显著。
3 讨论
镁对植物的逆境生理至关重要,适当增加镁元素的用量可显著增强植物抵御外界胁迫的能力,从而促进植物生长,在水稻[12]、药用菊花[13]和柑橘[14]上均已证明适当地增加镁水平,可显著提高植株的抗逆能力。在30mol L-1的镉胁迫条件下,西洋参幼苗体内O2-产生速率加快,导致单线态氧、H2O2 等氧化分子积累,引起膜脂过氧化,细胞膜透性增大,细胞内重要的物质大量外渗、外界的有毒物质进入细胞,细胞内新陈代谢发生紊乱,西洋参幼苗的生长发育受到抑制。本试验中,提高Mg2+的用量,西洋参幼苗的根系活力会得到提高。增加适量镁能维持细胞膜的完整性和稳定性,可降低质膜的透性,减少细胞内的外渗物,为细胞抵御不良外界环境提供了良好的生理基础[15],有效提高了西洋参幼苗受到镉胁迫时的根系活力。
由试验结果可知,外源镁抑制了西洋参幼苗根系中的Cd2+向地上部运输及各部位镉含量的积累,对地上部镉毒害的缓解作用大于根系,这可能是因为镁提高了植物螯合态合成酶的活性,植物螯合态合成酶在细胞质中聚合形成植物螯合肽(phytochelatin,PC)[16],PC具有很强的重金属亲和力,与西洋参幼苗从根部中吸收的Cd2+螯合后形成无毒的化合物Cd-S4-complex[17],这些化合物随即被转运到西洋参幼苗根系细胞液泡中,并且在其他酶的作用下被排出细胞外,从而降低了西洋参幼苗细胞内游离的Cd2+浓度,且防止了对Cd2+敏感酶变性失活[18]。通过图4 可知随着供镁水平的提高,西洋参各部位的镉含量降低,这可能是因为Cd2+、Mg2+同为二价阳离子,提高供镁水平,即增加了根系周围的Mg2+的量,这可能与降低了Cd2+通过根系Mg2+通道进入植物体内的概率有关。
植物在受到镉胁迫后,细胞膜透性增加、合成代谢能力下降、线粒体形态改变,同时SOD、POD 和过氧化氢酶(CAT)等保护性酶的活性降低,清除自由基及过氧化物的能力减弱[19];同时,活性氧自由基伤害加重,膜脂过氧化程度增强也会引起膜脂过氧化产物MDA 含量的增加[20]。本试验中,西洋参幼苗叶片在30mol L-1的镉胁迫条件下POD、SOD 和CAT 活性显著升高、MDA 含量显著降低,表明镉胁迫导致西洋参幼苗植株的脂质过氧化水平增加,对西洋参植株造成了过氧化伤害。与单独镉胁迫处理相比,增加Mg2+供应显著提高了镉胁迫下西洋参幼苗叶片中POD、SOD 和CAT 活性,并降低了MDA 含量,这可能是因为植物体内活性氧的清除由某些酶系统及抗氧化物质来完成,随着外源镁的增加,致叶片中的镁含量增加,镁通过提高酶及抗氧化物质的活性,达到提高POD、SOD 和CAT 的活性及降低MDA 含量的效果。
重金属对植物生长和代谢的影响是多方面的,其中对光合作用的抑制尤为显著,叶绿素含量的高低直接影响植物光合作用水平[21]。本试验中,西洋参幼苗叶片的光合色素含量在镉胁迫下显著下降,随着镁的加入得到缓解,与生物量变化规律相似,这是因为镁是构成叶绿素分子的中心元素,随着外源镁的加入,叶片中的镁含量增加,进而促进叶绿素的合成[22]。且叶绿素a 比叶绿素b 对Cd2+更加敏感,这是因为Cd2+会使类胡萝卜素等捕光色素复合体受损,破坏叶绿素酶系统,阻碍叶绿素a 的合成,导致光合系统Ⅱ比光合系统Ⅰ更敏感,进而表现出叶绿素a 比叶绿素b 变化得更为显著[23]。
叶绿素荧光参数反应光合机构对光能吸收、传递和转化的能力,对逆境反应十分敏感[24]。在镉胁迫下,西洋参幼苗的PSII 潜在活性Fv/Fm、Y(II)及相对电子传递速率显著降低、非光化学淬灭系数Y(NPQ)显著升高,这表明叶片受到了严重的光抑制,造成叶片的量子产量降低、电子传递减缓,进而降低光合速率[25],在加入外源镁后,Fv/Fm、Y(II)及相对电子传递速率显著升高,说明镁可缓解镉胁迫对光合机构的破坏,但当Mg2+浓度为5 mmol L-1时,较4 mmol L-1增幅并不显著,说明对于缓解西洋参幼苗镉毒胁迫,镁的用量不需要过高;非光化学淬灭系数Y(NPQ)显著降低,这说明植株受到的胁迫有所缓解,但CM3 较CM2 差异不显著,又进一步说明了缓解西洋参幼苗镉毒胁迫,镁的用量不可过高。
4 结论
猜你喜欢
少儿科学周刊·少年版(2021年17期)2021-01-17
金桥(2020年11期)2020-12-14
阅读(科学探秘)(2020年8期)2020-11-06
家庭百事通·健康一点通(2020年9期)2020-10-09
金桥(2020年7期)2020-08-13
家庭科学·新健康(2020年7期)2020-08-06
绿色科技(2019年2期)2019-05-21
食品界(2018年8期)2018-09-03
现代养生·下半月(2018年8期)2018-03-12
女性天地(2016年10期)2017-04-25
