
金钱松群落优势种群种间联结及群落稳定性研究
2021-04-12 07:41谢春平刘大伟南程慧李浩黄晨杨张健
生态科学 2021年1期
谢春平, 刘大伟, 南程慧, 李浩, 黄晨杨, 张健
金钱松群落优势种群种间联结及群落稳定性研究
谢春平1, 2, 刘大伟2, *, 南程慧2, 李浩2, 黄晨杨2, 张健2
1. 广东海洋大学滨海农业学院, 湛江 524088 2. 野生动植物物证技术国家林业和草原局重点实验室, 南京 210023
种间联结反映了植物群落中的种间关系和群落稳定程度, 对群落恢复和目标物种的保护具有重要意义。为了解金钱松群落的种间关系及群落稳定性, 采用方差比率法()、2检验、联结系数(AC)、共同出现百分率(PC)、Pearson相关系数、Spearman秩相关系数以及M. Godron稳定性测定等数量分析方法, 对浙江长兴金钱松群落中的18个优势种群、153个种对的种间相关性及群落稳定性进行分析。结果表明: 金钱松群落内的18个优势种群总体联结性呈不显著的正联结, 植物种类累积倒数百分比与累积相对频度交点坐标为(40, 60), 远离稳定交点坐标(20, 80), 说明群落处于不稳定状态。卡方检验表明绝大多数种对之间的联结性均不显著, 只有2个种对存在显著关联。联结系数和共同出现百分率也表明, 群落各种对相关性较弱, 各物种间呈独立分布格局。Pearson相关检验和Spearman秩相关检验发现, 金钱松与水竹之间呈显著负相关。结论: 金钱松群落种间关联性弱, 群落处于不稳定阶段。为保护和促进金钱松种群的恢复, 应进行适当的人为干扰和限制竹类植物在群落中的生长。
种间联结; 相关系数; 群落稳定性; 金钱松
0 前言
任何一种植物在群落中的存在都不是孤立的, 它是与环境及其他物种长期相适应的结果, 由此导致植物在群落空间上的分布呈现出一定的关联性[1]; 因此种间联结是植物群落重要的数量和结构特征之一, 是群落结构形成和演化的重要基础[2]。一方面, 种间联结分析是探讨森林群落稳定性和演替过程中种间关系变化情况的有效手段[3]; 另一方面, 物种之间相互依存、竞争或随机出现等综合情况可通过种间联结得到充分的体现[4]。对植物群落种间联结的研究不仅有利于对群落结构、功能的清楚认识, 亦可为森林经营、种群恢复、生物多样性保护等提供科学依据[5]。对于珍稀濒危植物而言, 种间关系的研究对于理解群落中物种的共存机制尤其是了解目标物种在群落中的地位具有重要意义, 也是成功实现就地保护的重要基础[6]。因此, 研究群落植物种间关系对植被管理和物种保护具有重要的指导意义。
地球气候环境正经历重大变化, 这种变化对珍稀濒危植物的影响显得尤为突出[7]; 如何科学有效地评估珍稀濒危植物在群落中的现状, 是保护工作的基础。种间联结性可用于揭示濒危机制, 了解群落竞争状况及演替趋势, 这对于正确指导濒危物种保护和引种栽培具有积极作用[8]。国内已就珍稀濒危物种生存群落的种间关系做了大量的研究, 如桫椤()[9]、云南红豆杉()[10]、华顶杜鹃()[11]等。这些研究均指出了目标物种在群落中与其他物种的关系及其对资源的利用情况, 研究成果对珍稀濒危植物的保护工作的开展具有较高的实践价值。
金钱松((Nelson) Rehd.)隶属于松科(Pinaceae)金钱松属(), 是我国特有的单属种植物, 现已被列为国家II级重点保护植物。根据化石证据表明, 该物种曾广泛分布于我国许多地区, 在经历了第四纪大冰期后, 现仅存活于长江中下游的华中和华东地区[12]; 因此, 该物种在研究松科及裸子植物进化史上具有重要的意义[13]。本研究在野外调查的基础上, 基于2×2 联列表, 采用方差比率()、2检验、联结系数AC、共同出现百分率PC、Pearson 相关系数检验和Spearman轶相关系数检验等方法对分布于浙江长兴县的金钱松群落种间联结及相关性进行分析, 同时结合M. Godron群落稳定性分析, 以期揭示该区域金钱松群落的种间关系和群落稳定性, 为维护金钱松群落的健康发展, 进而有针对性对该物种提出科学的保护策略、种群恢复等提供理论参考依据。
1 材料与方法
1.1 研究地概况
研究区位于浙江省北部的湖州市长兴县(30° 43′—31°11′ N, 119°33′—120°06′ E), 属低山丘陵向太湖西岸平原过渡地区, 全域面积约1430 km2。该县属亚热带海洋季风气候, 气候整体表现为四季分明、雨热同期、气候温和、降雨充沛等特点。该区域年均温15.8 ℃, 最低温和最高温一般出现在1月和7月。年均降水1347 mm, 其中75 %的降雨量集中在5月—10月期间, 同时年均蒸发量为800— 900 mm。年均日照时数约1810 h, 历年平均日照百分率为41 %, 光照分配较均匀, 年均无霜期250 d[14]。长兴县土壤以红壤土类、水稻土类、潮土类和岩性土类等4种类型为主[15]。根据实地调查, 研究地的金钱松群落乔木层优势种以金钱松、毛竹()、冬青()、盐肤木()、苦槠()、栗()、黄檀()、水竹()等为主; 灌木层以茶()、野蔷薇()及其他乔木层树种的幼树幼苗居多。
1.2 研究方法
1.2.1 样地调查与设置
在金钱松分布较为典型的区域随机设置半径为6 m的样圆21个。对样圆内胸径≥2 cm的乔木(包括竹类植物)进行每木调查, 记录种名、胸径、高度、冠幅等。在样圆内垂直四个方向各设置1个2 m×2 m的灌木调查样方, 记录灌木的种名、高度、冠幅等。同时, 记录每个设样地地理坐标、群落郁闭度、干扰情况及环境因子(海拔、土壤、坡向、坡度、凋落物厚度等)。根据调查结果, 最终18个重要值大于1 %的物种被选为研究对象。
1.2.2 分析方法
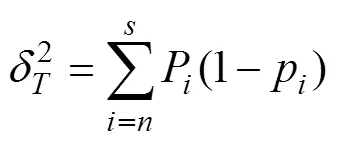
(2)2检验。利用2检验来判断种间关联与否: 假设种A和B相互独立, 通过2值来检验这一假设, 其Yates的连续纠正公式为:2=[|-|-0.5]2/[(+) (+)(+)(+)]; 式中,、、、分别表示A、B两个种在样圆中出现的情况, 即为同时出现,为只有物种B出现,为只有A出现,为物种A与物种B均不出现; 样方总数=+++[19]。2检验仅得出了种间联结性显著与否的结论, 没有显著联结性的种对不代表没有联结性, 而联结系数(AC)和共同出现百分率(PC)则可对联结程度加以反映[8]。
(3)联结系数AC。当≥时, AC=(–)/ [(+)(+)]; 当>且≥时, AC=(–)/[(+)(+)]; 当>且<时, AC=(–)/[(+) (+)][20]。
(4)共同出现百分率PC。为避免因值所产生的AC偏差, 利用PC来测定种间联结程度, 其表达式为: PC=/(++)[21]。
联结系数AC及共同出现百分率PC中的、、、与2检验公式的含义相同; 同时, 为避免分母为0而出现无法计算的情况, 将和值加权为1, 以获得较为客观的效果[4]。
(5)种间相关性。根据Pearson相关系数和Spearman轶相关系数对金钱松群落内2个物种间的相关性进行分析[3, 11, 21]。其中:
重要值计算公式为: [相对多度()+相对频度()+相对优势度()]/3[22]。
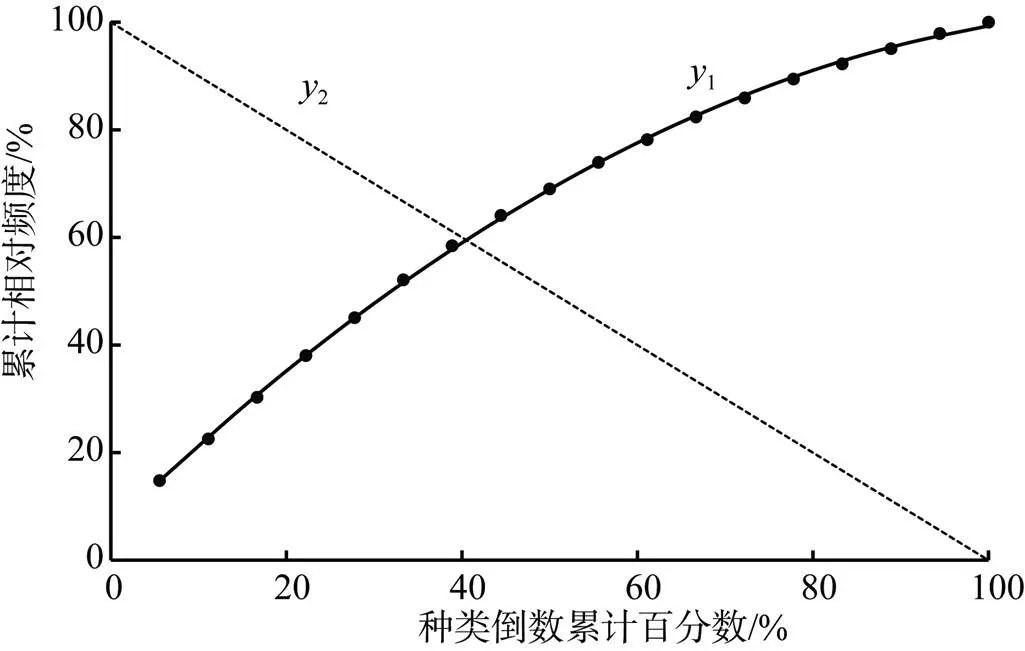
(6)群落稳定性分析。根据改良的M. Godron稳定性测定方法, 利用本研究中的18个优势种群的相对频度的累计值和物种数倒数的累计值建立数学模型; 通过拟合方程=2++和= -+ 100获得交点值和, 由此推断群落演替过程中主要物种变化趋势。若交点值越接近(20, 80)则表示群落越稳定[8, 23, 24]。
2 结果与分析
2.1 总体联结性


表 1 金钱松群落主要种群总体关联性检验
2.2 x2检验、联结系数与共同出现百分率分析
Yates的连续校正公式的2检验是基于2×2列表中种对在样方中出现的次数关系所构建的(自由度为1), 因此当3.841≤2<6.635和2≥6.635时分别表示种对之间具有显著和极显著的关系, 同时亦可通过>、=和<反映种对之间的正相关、不相关和负相关的关系[25, 26]。由表2的2检验统计的结果可知, 在金钱松群落内153个优势种对中有76个种对呈现为正相关, 75个种对为负相关, 2个种对表现为不相关。其中, 仅枸骨()-朴树()表现为极显著正相关(6.52), 及化香树()-小叶栎()表现为显著正相关(5.52); 金钱松-苦槠, 化香树-黄檀表现为不相关。因此,2检验结果表明在金钱松群落中相关性不显著的种对占到整体的97.39 %。这一结果较好地反映出群落中物种之间的关联性较弱。
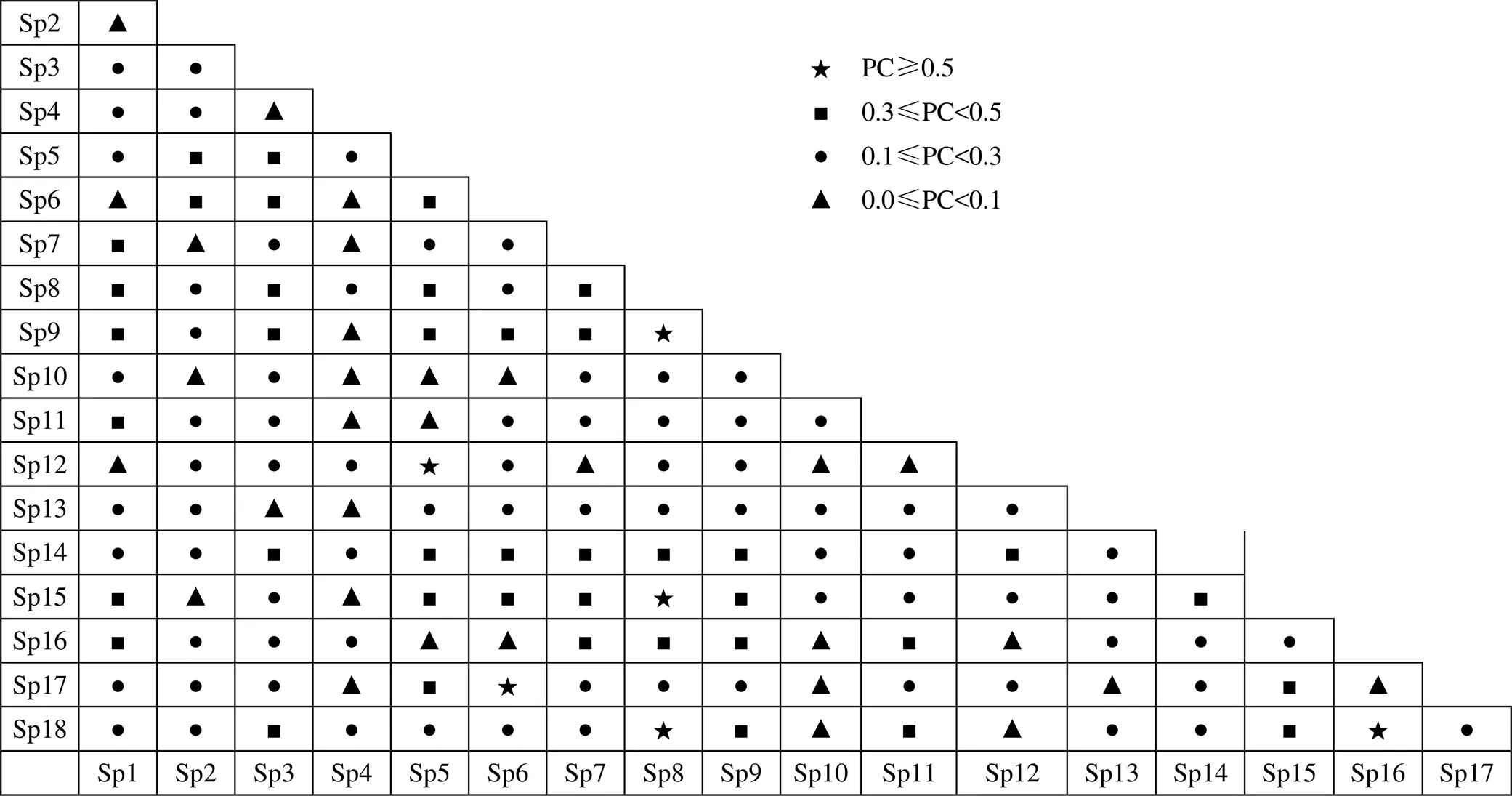
2统计量虽能客观、精确地反映出种对间联结的显著程度, 但仅能作定性判断, 不能区分联结强度的大小, 这在一定程度上模糊了种间关联性之间的差异性[2, 4]; 因此应结合AC、PC等系数加以判读。从金钱松群落优势种间联结系数的半矩阵图(图1)可知, 群落中AC≥0.5的种对有8对, 占总对数的5.23 %, 其中茶条槭()与枸骨、化香树、山胡椒(), 山胡椒与朴树、青榨槭(), 盐肤木与牡荆(var.)、白杜()等种对间的正联结程度较高。0.3≤AC<0.5的种对共有17对, 占总对数的11.11%, 如枫香树()-化香树、黄檀-栗、苦槠-枸骨、小叶栎-茶条槭等。-0.3≤AC<0.3共计有92对, 超过总数的60 %, 在整个优势种对间占有较大的部分, 如黄檀-盐肤木、冬青-牡荆、金钱松-朴树等。-0.5≤AC<-0.3共计有17对, 占总对数的11.11 %, 如冬青-水竹、枫香树-苦槠、山胡椒-白杜、毛竹-水竹等。AC<-0.5的种对有19对, 占总数的12.42 %, 如构骨-毛竹、毛竹-白杜、化香树-栗等。目标树种金钱松与其他优势种的AC值多数在[-0.4, 0.01]的区间, 说明它与其他物种的正联结性较弱。AC的值域为[-1, 1], 其值越趋近于1, 表明种间正联结性越强; 其值越趋近于-1, 表明种间负联结性越强; AC=0, 表示物种间相对独立[19]。因此通过上述AC结果可知, 该群落中种对间的正联结性或负联结性并不明显, 各种对间趋于独立或随机分布; 这一结果与2检验是相互验证的。
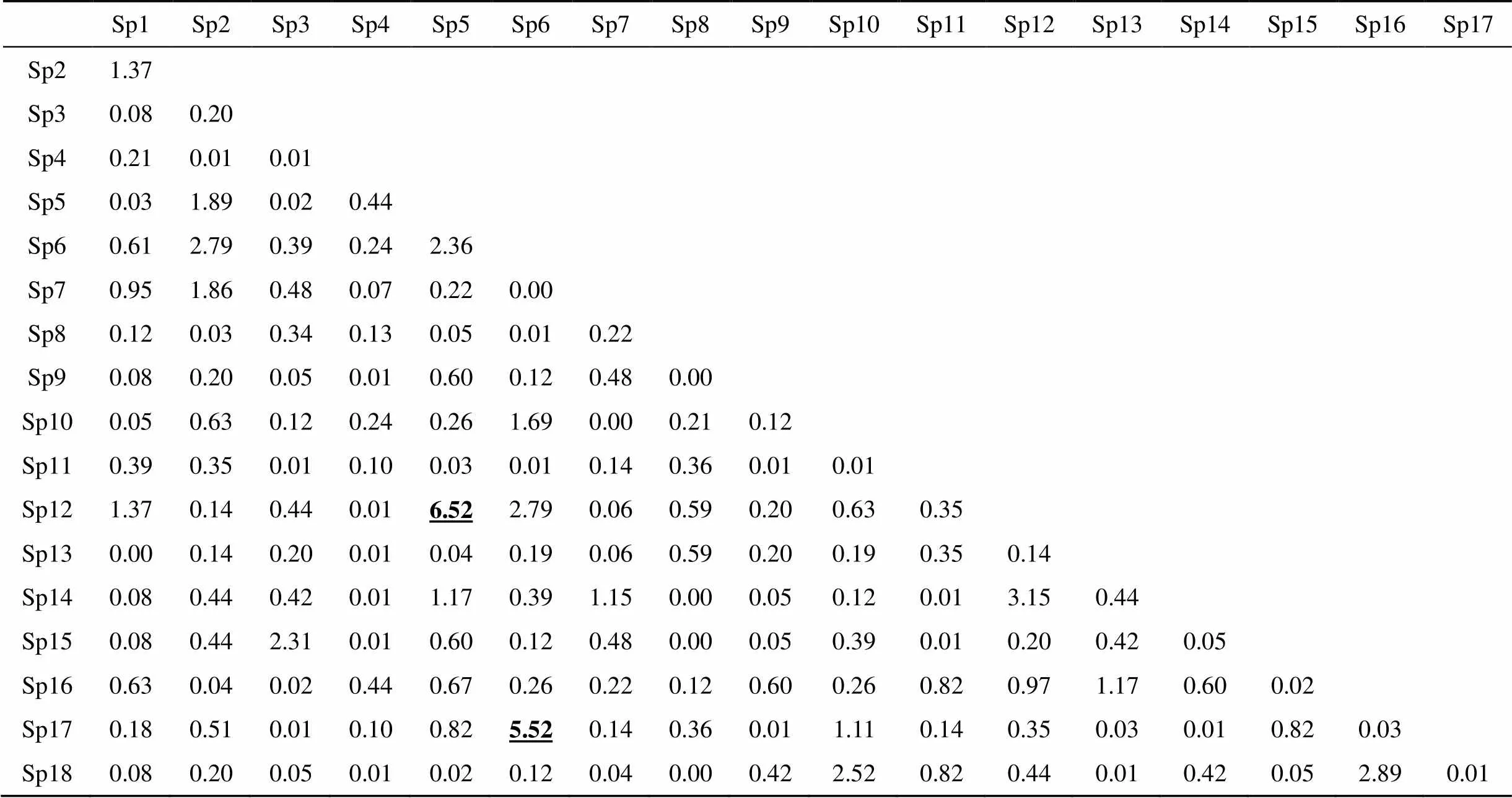
表2 金钱松群落优势种群x2统计量检验
注: 表2中Sp1—Sp18分别表示金钱松群落内的主要优势树种: 栗()、茶条槭(、冬青()、枫香树()、枸骨()、化香树()、黄檀()、金钱松()、苦槠()、毛竹()、牡荆(var.)、朴树()、青榨槭()、山胡椒()、水竹()、白杜()、小叶栎()、盐肤木()。

注: 物种编号含义见表2。
Figure 1 Semi-matrix diagram of association coefficients (AC) for dominant population in the community of
共同出现百分率(PC)可解释种间正联结程度的高低, 因为两物种共同出现的可能性越大, 则两物种的生态习性和对环境的需求越一致[18, 27]。根据金钱松群落主要种群共同出现百分率半矩阵图(图2)可知, PC值大于0.5的仅有6对, 占总对数的约4 %, 如金钱松-盐肤木、化香树-小叶栎、苦槠-金钱松等, 它们的值均在0.55左右。余下0 根据Pearson相关系数的结果显示(表3), 62个种对的相关系数为正, 占所有种对的40.52 %; 其中有7个种对呈现出极显著正相关(<0.01), 6个种对呈现显著相关(<0.05)。小叶栎-茶条槭(0.81)、朴树-化香树(0.64)、枸骨-枫香树(0.69)等均表现出极显著相关。此外, 有89个种对的相关系数为负, 占所有种对的58.17 %, 其中仅水竹-金钱松为显著负相关(-0.61), 其他种对间均无显著性。无关联的种对有2个, 占总体的1.31 %。 注: 物种编号含义见表2。 Figure 2 Semi-matrix diagram of co-occurence rate for dominant populations in the community of 对Spearman轶相关系数统计分析表明(表3), 79个种对的相关系数为正, 占总体的51.63 %; 其中有3个种对呈现出极显著正相关(<0.01), 6个种对呈现显著正相关(<0.05)。如小叶栎-化香树(0.59)、朴树-枸骨(0.72)及朴树-化香树(0.57)均表现出极显著的正相关性。74个种对的相关系数为负, 占总体的48.37%, 仅1个种对呈极显著负相关(<0.01), 即金钱松-水竹(-0.60)。 表3 金钱松群落主要种群Pearson相关系数和Spearman轶相关系数半矩阵 注: 表的右上部分为皮尔森相关系数, 左下部分为轶相关系数; 物种编号含义见表2。 Pearson相关系数和Spearman轶相关系数虽然在相关系数正负比例上有一定的差别, 但在整体上均表现出种间显著相关的占比均较低, 它们的结果与2检验、AC等参数相接近。 根据种类倒数累计百分数与累计相对频度制作散点图(图3), 其拟合曲线为:1= -0.00652+ 1.5772+ 6.206(R² = 0.9998)。曲线1与曲线y=100-的交点大致为(40, 60), 这与稳定群落的交点值(20, 80)相距甚远; 由此可知, 金钱松群落仍处于不稳定状态。本研究是利用群落内重要值大于1%的物种参与模拟, 它们重要值的总和已超过80%, 具有较好的代表性。学者指出, 以线性演替模型为基础的稳定性研究, 乔木树种对群落具有决定性的支配作用, 影响和决定着群落的最终发展趋势[23]; 因此, 本研究以金钱松群落内主要乔木树种为基础所构建的群落稳定性模型, 较好地反映了群落不稳定的特征。 种间联结性反映了物种间的相互影响和群落动态, 种群间总体关联性体现了群落演替的阶段及其稳定性, 而群落稳定性是种间联结性的具体表现[28]。对于地带性顶级群落, 由于群落已对当地的气候环境形成了高度的适应性, 同时群落内的物种之间的相互作用和对环境利用关系已趋于平稳, 因此其总体关联性检验的方差比率值应远大于1[8]; 同时亦表现出正负联结种对数量比值较高的情况。如有学者对浙江天台山常绿阔叶林优势种群的种间联结研究表明, 乔木层和灌木层主要种类间在总体上均呈显著的正相关, 正联结的种对多于负联结的种对[29]。反之, 群落若在总体上呈现负联结趋势, 呈极显著和显著正联结的种群所占比率较低, 则表明群落仍然处于不断的发展演替过程之中[30]。一般来说, 处于演替早期的群落, 物种间的联结程度较低, 正负联结比值偏小, 群落处于不稳定阶段[31]。如热带海岸香蒲桃()天然次生林群落, 乔木层正关联不显著, 说明优势种间存在一定的联系, 但未能形成稳定的搭配关系, 群落的稳定性不高[32]。与香蒲桃群落相似, 本研究总体联结性值大于1, 但偏离1的程度不高, 同时统计量值表明这种正联结无显著性。改进后的M. Godron群落稳定性交点值为(40, 60), 与理想的群落稳定值(20, 80)仍有较大的差距, 与群落总体联结性不显著的结果相印证; 这也说明了本群落的稳定性不高的特点。 图3 金钱松群落稳定性分析图 Figure 3 Stability graph of the community of 利用2检验与联结系数AC较好地阐释了金钱松群落的种间关联性。一方面,2检验表明呈显著或极显著的种对较少, 而无显著性的正相关与负相关的种对基本持平; 另一方面, 联结系数AC也显示出>0.5的正相关系数和<-0.5的负相关系数的种对均较少; 因此, 种对之间的关联性并不高。这种情况也在其它濒危植物群落内也有所体现[6, 8, 20, 21, 25], 多数可能是因为群落处于演替的初期或受外界干扰较大。在本研究中, 一方面诸如金钱松、枫香树等早期先锋树种在群落乔木上层已占据有利地位, 其对光、热、水等资源的利用具有较大的优势。另一方面, 青冈、苦槠、冬青、枸骨等常绿树种开始逐渐渗透入群落; 由于它们具有较好的耐阴等生态学特性, 在群落亚乔木层或灌木层的种群数量也开始逐渐增多。这一判断通过Pearson相关系数检验时, 由枸骨-枫香树呈极显著相关的结果得到验证。当群落处于远离顶极群落的不稳定阶段时, 组成群落的物种是松散的、偶然的聚合, 其总体关联性趋于无关联或显著负关联[33]。因此, 这就较好地说明了金钱松群落处于演替的初期, 群落种对间的关系仍较复杂。 一般情况下, 随着植物群落的顺行演替, 群落内各种类的种内竞争和种间竞争均趋于稳定, 群落的种间联结性以正关联为主, 使得群落中各种群能够和谐共存[34]。因为随着群落演替进展, 物种与群落之间相互选择和适应, 种间正关联程度逐渐增大; 当群落演替到顶级阶段时, 群落的种类之间常常会达到某种协调, 正关联优势明显, 生态位彼此分化, 最终达到稳定共存状态[3]。由共同出现百分率PC值可知, 整个群落内相似性较高的种对仅有4 %, 这说明现存优势种进入群落的随机性与松散性。种对间对群落内的资源利用虽存在一定的竞争, 但可能由于存在生态学和生物学的差异、或是群落垂直结构差异等原因, 它们仍可在一定程度上互补, 种对间还不至于排斥; 例如金钱松与盐肤木、化香树、苦槠等物种的PC值均大于0.5。 结合2检验、联结系数AC和共同出现百分率PC的分析结果发现, 金钱松与其他17个物种间均不存在显著的正负相关性性。但利用Pearson和Spearman两个相关系数检验结果发现, 金钱松与水竹之间存在显著的负相关; 这一结果应引起关注。因为, 竹类植物若在群落形成优势局面后, 不仅可导致群落结构简单化、生物多样性降低, 而且群落植物的空间分布格局也会发生改变[35-37]。这是因为竹类植物特殊的繁殖方式以及其对环境的耐受性要强于其他物种, 而更新层金钱松幼树幼苗的缺乏可较好地解释这种显著负相关情况出现的原因。 利用2检验、联结系数AC和共同出现百分率PC可对群落复杂的种间关系进行科学阐释, 对揭示群落演替阶段、功能结构、系统发育等方面具有重要作用, 同时为目标物种的保护和植被恢复具有重要的指导意义。总体联结性表明金钱松群落总体上的正联结不显著, 方差比率偏离1的程度较小; 而群落稳定性的结果也表明群落处于不稳定的阶段。2检验表明种间关联呈显著性的仅2对, 多数种对无显著性。多数种对的AC值表现出不明显的关联特性, 种对间PC值较低的结果也支持了上述结论。Pearson相关系数和Spearman轶相关系数的检验结果支持上述种间关系分析。综上所述, 金钱松群落种间关系较为松散, 种对间的关联性较弱,群落处于不稳定演替阶段; 该群落的物种组成、结构功能等均处于动态调节过程中。因此, 应利用种间关系研究的结果对群落进行科学维护, 促进群落趋于稳定, 以实现金钱松种群的恢复。 金钱松作为国家Ⅱ级重点保护植物在华东地区的种群数量和分布面积呈严重缩减的态势, 加大对其野生种群的保护工作已刻不容缓。因此, 对该地区金钱松的保护笔者建议: 首先, 注意梳理群落内与金钱松呈负相关的物种, 尤其是当前群落内出现的水竹已严重影响了金钱松种群的更新。其次, 由于金钱松属阳性树种, 应对郁闭度较高的群落采取适度的人工抚育措施, 如林窗开辟、倒木清理、减少其他灌木物种等。再者, 应注重金钱松幼树幼苗的抚育, 尤其是提高幼苗的成活率, 并通过建立保护小区等措施实现种群的稳定发展。 致谢:黔南民族师范学院俞筱押教授在数据处理方面给予了指导与帮助, 在此表示诚挚谢意。 [1] 唐愽言, 操国兴, 蒋万平. 武陵山大木山自然保护区红豆杉群落优势乔木种群生态位和种间联结[J]. 植物分类与资源学报, 2014, 36(5): 668–674. [2] 徐满厚, 刘敏, 翟大彤, 等. 植物种间联结研究内容与方法评述[J]. 生态学报, 2016, 36(24): 8224–8233. [3] 朱德意, 龙佳峰, 李远发, 等. 南盘江流域细叶云南松次生林群落主要木本种群的种间联结[J]. 西北植物学报, 2019, 39(3): 534–542. [4] 江焕, 张辉, 龙文兴, 等. 金钟藤入侵群落的种间联结及生态位特征[J]. 生物多样性, 2019, 27(4): 388–399. [5] 刘润红, 陈乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J]. 生态学报, 2020, 40(6): 1–15. [6] 俞筱押, 田华林, 郭治友. 贵州南部伯乐树群落特征及其种间关系研究[J]. 四川农业大学学报, 2016, 34(1): 29– 33. [7] 吕佳佳, 吴建国. 气候变化对植物及植被分布的影响研究进展[J]. 环境科学与技术, 2009, 32(6): 85–95. [8] 周赛霞, 彭焱松, 丁剑敏, 等. 珍稀植物狭果秤锤树群落木本植物种间联结性及群落稳定性研究[J]. 广西植物, 2017, 37(4): 442–448. [9] 李丘霖, 宗秀虹, 邓洪平, 等. 赤水桫椤群落乔木层优势物种生态位与种间联结性研究[J]. 西北植物学报, 2017, 37(7): 1422–1428. [10] 李帅锋, 刘万德, 苏建荣, 等. 滇西北云南红豆杉群落物种生态位与种间联结[J]. 植物科学学报, 2012, 30(6): 568–576. [11] 蔡鑫, 陈波, 陈锋, 等. 珍稀特有植物华顶杜鹃的种群结构和种间联结[J]. 浙江大学学报(理学版), 2019, 46(3): 354–363. [12] 魏学智, 胡玉熹, 林金星, 等. 中国特有植物金钱松的生物学特性及其保护[J]. 武汉植物学研究, 1999, 17(S1): 73–77. [13] 吴毅, 曹基武, 周国英等. 湖南衡山优势种群金钱松群落研究[J]. 中南林业科技大学学报, 2012, 32(11): 85–88. [14] 白炳书. 长兴县坡地开发的水土流失防治措施[J]. 中国水土保持, 2014(4): 36–37. [15] 刘政. 长兴县毛竹高效培育技术集成与示范[D]. 杭州: 浙江农林大学, 2010. [16] SCHLUTER D. A Variance Test for Detecting Species Associations, with Some Example Applications[J]. Ecology, 1984, 65(3): 998–1005. [17] 岳鹏鹏, 周静静, 王华. 毛乌素沙地东缘黑沙蒿群落种间联结性分析[J]. 生态科学, 2017, 36(6): 147–152. [18] 谢春平, 赵浩彦. 城市近郊构树群落主要种群种间联结及物种多样性研究[J]. 中南林业科技大学学报, 2017, 37(7): 85–91. [19] 高浩杰, 袁佳, 高平仕. 舟山群岛次生林优势种群的生态位与种间联结[J]. 热带亚热带植物学报, 2017, 25(1): 73–80. [20] 王博, 王亮, 王立龙, 等. 濒危植物裸果木()与其伴生种种间联结性及群落稳定性[J]. 中国沙漠, 2017, 37(1): 86–92. [21] 张滋芳, 毕润成, 张钦弟, 等. 珍稀濒危植物矮牡丹生存群落优势种种间联结性及群落稳定性[J]. 应用与环境生物学报, 2019, 25(2): 291–299. [22] 邱靖, 伊贤贵, 汤庚国, 等. 黄山水榆花楸群落结构分析[J]. 四川农业大学学报, 2016, 34(3): 304–311. [23] 郑元润. 森林群落稳定性研究方法初探[J]. 林业科学, 2000, 36(5): 28–32. [24] 王林林, 唐坚强, 张光富, 等. 南京朴树林竞争强度及其群落稳定性[J]. 生态与农村环境学报, 2012, 28(6): 654– 660. [25] 林永慧, 何兴兵, 胡文勇, 等. 濒危植物缙云卫矛群落种间联结分析[J]. 植物科学学报, 2011, 29(1): 58–63. [26] 林伟强, 刘惠明, 张璐. 广州帽峰山次生林主要种群种间联结性研究[J]. 生态科学, 2004, 23(1): 42–46. [27] CÁRDENAS RE, VALENCIA R, KRAFT NJB, et al. Plant traits predict inter-and intraspecific variation in susceptibility to herbivory in a hyperdiverse Neotropical rain forest tree community[J]. Journal of Ecology, 2014, 102(4): 939–952. [28] 刘润红, 姜勇, 常斌, 等. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析[J]. 生态学报, 2018, 38(19): 6881–6893. [29] 金则新. 浙江天台山常绿阔叶林优势种群结构及种间联结性研究[J]. 广西植物, 2002, 22(3): 203–208. [30] 刘淑燕, 余新晓, 陈丽华. 北京山区天然林乔木树种种间联结与生态位研究[J]. 西北林学院学报, 2009, 24(5): 26–30. [31] 郭忠玲, 马元丹, 郑金萍, 等. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究[J]. 应用生态学报, 2004, 15(11): 2013–2018. [32] 杨青青, 杨众养, 陈小花, 等. 热带海岸香蒲桃天然次生林群落优势种群种间联结性[J]. 林业科学, 2017, 53(9): 105–113. [33] 林勇明, 洪滔, 吴承祯, 等. 桂花次生林群落主要树种种间关联及其对混交度的响应[J]. 应用与环境生物学报, 2007, 13(3): 327–332. [34] 俞筱押, 余瑞, 黄娟, 等. 贵州茂兰国家级自然保护区四药门花群落各层次优势种群的种间联结性分析[J]. 植物资源与环境学报, 2018, 27(2): 24–32. [35] 欧阳明, 杨清培, 陈昕, 等. 毛竹扩张对次生常绿阔叶林物种组成、结构与多样性的影响[J]. 生物多样性, 2016, 24(6): 649–657. [36] 白尚斌, 周国模, 王懿祥, 等. 天目山保护区森林群落植物多样性对毛竹入侵的响应及动态变化[J]. 生物多样性, 2013, 21(3): 288–295. [37] 黄茹, 齐代华, 陶建平, 等. 竹类入侵干扰对桫椤种群空间分布格局的影响[J]. 四川师范大学学报(自然科学版), 2009, 32(1): 106–111. Study on community stability and interspecific correlations among dominant populations incommunities XIE Chunping1, 2, LIU Dawei1, 2,*, NAN Chenghui1, 2, LI Hao1, HUANG Chenyang1, ZHANG Jian1 1. College of Coastal Agricultural sciences, Guangdong Ocean University, Zhanjiang 524088, China 2.Key Laboratory of State Forest and Grassland Administration on Wildlife Evidence Technology, Nanjing 210023, China Interspecific correlations of plants reflect relationships among species in the community as well as community stability, and it has critical theoretical significance and application values for the restoration of vegetation and the conservation of aim species. In order to illustrate the interspecific correlations and the stability of communities for, 18 dominant species, 153 species pair ofcommunities were studied by using quantitative analysis methods including the variance ratio (), Chi-square test, association coefficient (AC), percentage co-occurrence (PC), Pearson correlation coefficient, Spearman rank correlation coefficient, and M. Godron stability analysis in Changxing county, Zhejiang province. Results indicated that there were positive associations between dominant species totally without significant. The coordinate (40, 60) between the inverse accumulative percentage of species and their accumulative relative frequency was far from the stability point (20, 80), which showed that these communities were in an unstable state. The chi square test interpreted that the association most of the species pairs was not significant, and only a two species pairs of the community had a significant association. A vast majority of the species pairs were a weak association and performed an independent distribution pattern reflected by the results of AC and PC. Based on the result of Pearson correlation coefficient and Spearman rank correlation coefficient, there was significant negative associations betweenand. Therefore, according to the analyses mentioned above, we came to conclusion that the correlation among the species pair for this community was relatively weak and the community was still in an unstable succession stage. To protect and improve the population restoration, not only the aim communities should be interfered by artificial measure, but also bamboos should be cleaned in the communities. Interspecific correlations; correlation coefficient; community stability; 谢春平, 刘大伟, 南程慧, 等. 金钱松群落优势种群种间联结及群落稳定性研究[J]. 生态科学, 2021, 40(1): 62–70. XIE Chunping, LIU Dawei, NAN Chenghui, et al. Study on community stability and interspecific correlations among dominant populations incommunities[J]. Ecological Science, 2021, 40(1): 62–70. 10.14108/j.cnki.1008-8873.2021.01.009 S718.542 A 1008-8873(2021)01-062-09 2020-02-03; 2020-03-23 国家林业和草原局珍稀濒危物种野外救护与人工繁育项目(2019)资助 谢春平(1980—), 男, 海南儋州人, 副教授, 主要从事森林生态学研究。E-mail: xcp@gdou.edu.cn 刘大伟(1983—), 男, 安徽阜阳人, 讲师, 主要从事野生动植物保护与利用研究。E-mail: 272756332@qq.com2.3 Pearson相关系数与Spearman轶相关系数分析

2.4 群落稳定性分析
3 讨论
4 结论与保护建议
猜你喜欢
今日农业(2022年15期)2022-09-20
当代水产(2022年8期)2022-09-20
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
湖南电力(2021年1期)2021-04-13
河南科学(2020年3期)2020-06-02
红土地(2018年7期)2018-09-26
蔬菜(2018年5期)2018-05-17
杂草学报(2015年2期)2016-01-04
