
崇明岛内河夏季鱼类群落组成及分布特征
2021-04-07 03:19从婷婷童春富赵成建陈振涛贺湛斐刘毛亚
生态学报 2021年5期
从婷婷,童春富,*,赵成建,陈振涛,贺湛斐,刘毛亚
1 华东师范大学河口海岸学国家重点实验室,上海 200241 2 崇明生态研究院,上海 202162
鱼类作为水域生态系统健康的重要指示类群,其种类组成和空间分布对区域生态系统物质循环和能量流动起重要作用[1- 2]。近年来,生态学家逐渐认识到,鱼类群落结构通常会受到多尺度、多种环境因子的共同影响[3- 4],而河流应当被看作是一种“树状网络体”[5- 6],鱼类等水生生物的个体、种群、群落等的空间分布特征,不仅与特定河段的栖息地环境有关,还受到该河段在整个河流网络中的空间位置的影响[7- 8]。鱼类群落空间分布特征及其影响因素是水生生态学研究的重要问题之一[9]。
岛屿与大陆水系联系较少,这种独特的地理位置和有限的地域空间使得岛内水域生态系统,包括河流生态系统,具有明显的特殊性[10- 11],因此其鱼类具有重要的研究价值。目前国内对岛屿河流鱼类群落的研究还相对较少,仅见于海南、台湾等地区,研究内容主要包括鱼类区系的组成特点及其形成原因等[11- 13]。崇明岛地处长江河口,是我国的第三大岛,也是世界最大的河口冲积沙岛,岛内水网密布、河流众多。崇明岛地区关于鱼类群落的研究主要集中于滩涂及邻近水域鱼类的种类组成和多样性特征等方面[14- 15],而对广泛分布于整个岛屿内部水系的鱼类群落至今还没有系统的研究。
崇明岛目前的发展定位是世界级生态岛。在建设过程中,要以生态优先为基本原则,融自然、经济、社会于一体,彰显其显著的示范意义和国际综合竞争力[16]。崇明岛内河是贯穿全岛的生态脉络,是生态岛的重要组成部分。本文通过对崇明岛内河鱼类群落的调查和分析,初步阐明了崇明岛内河夏季鱼类群落组成和分布特征,探讨了影响岛屿内河鱼类群落组成及分布的主要因素,在丰富岛屿内河鱼类群落研究成果的同时,可以为崇明生态岛的建设提供基础资料。
1 研究区域与研究方法
1.1 区域概况
研究区域位于崇明岛。该岛呈长条形,东西向长约110km,南北向长约13—18km。该区域气候温和湿润,夏季盛行东南风,冬季盛行偏北风,四季分明,属于典型的亚热带季风气候,年平均气温15.2℃,年平均降雨量1025mm[17]。岛屿三面环江,一面临海。岛内河网水系完全由人工开挖而成,南横引河和北横引河2条市级河道横贯东西,多条县级竖河贯穿南北,基本形成了南引北排,西引东排的河网输水和调蓄系统。各河道河面平均控制水位在2.6—3m[18],水域面积约占全岛总面积的14%,地表水补给约90%来自长江引水,水质状况同时受到长江淡水径流和海水的影响[19]。在枯水期,受潮流作用和长江径流减少的影响,岛内咸水倒灌现象频发。近年来,随着生活污水、城镇废水和农业面源污染加剧,岛内河流出现了水体富营养化的问题,以崇明岛北横引河中下游地区最为典型[20]。河网水质具有明显的空间分异特征,整体水质西部优于东部,南部优于北部。北横引河全长约84km,现状河道两岸多为自然岸坡,受河道和雨水冲刷后可能造成一定的水土流失,以致河道淤浅,其分布区域周边主要为农田。南横引河是岛内水上运输的主航道,贯穿崇明的南部地区,全长约78km,河宽平均约50m,深约4m,承担着重要的防汛除涝功能[21- 22],其分布区域主要为人口集中分布的建城区。县级河道均为南北走向,河水南进北出,河宽20—30m,水深1.5—3.0m[23],为南、北横引河的连接、贯通通道。
1.2 站位布设与样品采集
本研究选取2条市级河道(南横引河与北横引河)和3条县级竖河(庙港、堡镇港、八滧港)布设16个调查站位。其中,南横引河和北横引河各布设5个调查站位,庙港、堡镇港、八滧港3条竖向河道各布设2个调查站位。根据调水方式、河网分布特征以及地形水文等条件将调查水系分为南横引河、北横引河、竖向河流3种不同的河段类型。站位布设如图1所示。
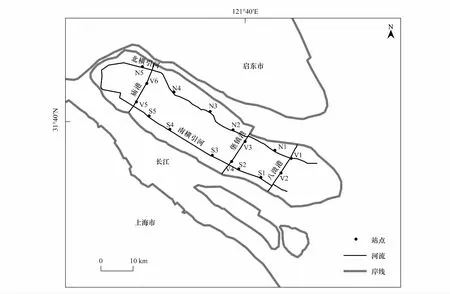
图1 崇明河道采样站位示意图Fig.1 Map of sampling sites in the rivers of Chongming Island
调查于2018年6月进行。调查网具为单拖网,网袋长度为45m,网口高度3.5m,有效宽度11m,网袋网目3.5cm;取鱼部(网囊)长度8m,网目为2cm。调查时,采用渔船进行拖网作业,船速控制在2—3节。每个调查站位拖1网,约15min,用GPS记录拖行距离和时间。所有渔获物带回实验室进行鉴定、计数,测定体长、生物量等指标。鱼类的命名参考《上海鱼类志》[24]。基于拖行距离和网口有效宽度算出每网的扫底面积,在此基础上将所记录的各站点渔获量(个体数、生物量)换算为单位面积(hm2)的渔获量(CPUE标准化)以便后续计算、比较。
1.3 数据处理
采用相对多度(%N)、相对生物量(%B)和Pinkas相对重要性指数(IRI)[25]反映鱼类群落的组成及优势度特征,将IRI>20的定为优势种。
相对多度:
%N=ni/N×100
相对生物量: %B=bi/B×100
相对重要性指数: IRI=(%N+%B)×fi/F
式中,N为各河段渔获物的总个体数;B为各河段渔获物的总生物量(鲜重);F为各河段的站位数;ni为各河段第i种鱼类的个体数;bi为各河段第i种鱼类的生物量;fi为各河段第i种鱼类出现的站位数。
使用Primer 5.2.8软件对鱼类丰度数据进行四次方根转换后,进行各站点间的Bray-Curtis相似性系数计算,构建相似性矩阵。采用等级聚类(Cluster)分析和非度量多维标度排序(nMDS)法,分析不同站点鱼类群落组成的相似性[26]。用胁强系数(Stress)来判断nMDS分析结果的可信度。当0
利用SPSS Statistics 22统计软件进行方差分析。采用Kruskal-Wallis H的非参数检验方法,研究不同河段共有优势鱼类的体长差异,并进行两两比较,以P<0.05表示显著差异,以P<0.01表示极显著差异[29]。作图在Origin 8.6中完成。
2 结果与分析
2.1 区系组成
调查期间共采集鱼类样本4417尾,分属6目9科30种,其中鲤形目鲤科鱼类种类最丰富,达19种,占总物种数的63.3%;鲇形目次之,有5种,占总数的16.7%;鲈形目3种,鲱形目、鳗鲡目、鮭形目各1种。从生态类型来看,以淡水鱼类为主,有27种,另外洄游性鱼类和近海鱼类分别为2种和1种,如表1所示。
调查期间,北横引河共记录鱼类4目6科20种,其中18种为淡水鱼类,其余2种各为洄游鱼类及近海鱼类。鲢Hypophthalmichthysmolitrix、鳙Aristichthysnobilis、鲤Cyprinuscarpio、鲇Silurusasotus、子陵栉鰕虎鱼Ctenogobiusgiurinus仅在北横引河有记录。南横引河共记录鱼类6目7科17种,其中14种为淡水鱼类,2种为洄游性鱼类,1种为近海鱼类。鳡Elopichthysbambusa、麦穗鱼Pseudorasboraparva、寡鳞飘鱼Pseudolaubucaengraulis、沙塘鳢Odontobutisobscurus、太湖新银鱼Neosalanxtaihuensis仅在南横引河有记录。三条竖河共记录到鱼类5目5科18种,其中15种为淡水鱼类,2种为洄游性鱼类,1种为近海鱼类,如表1所示。
2.2 丰度、生物量和优势种
从北横引河、南横引河和竖河各站位的平均物种数、丰度和生物量来看,北横引河各项指标值均高于南横引河和竖河,如图2所示。就物种数而言,南横引河最少,它与北横引河的共有种有8种,与竖河的共有种有9种,有5种鱼类仅在南横引河有记录。但单因素方差分析的结果则表明,物种数、丰度、生物量在不同河段间均不存在显著差异(P>0.05)。
在北横引河中,鲫Carassiusauratus、似鳊Pseudobramasimoni和翘嘴红鲌Erythroculterilishaeformis作为主要优势种(IRI>20),其相对多度和相对生物量亦占优势。其中,似鳊的优势度最高,是北横引河的绝对优势种。另外,鳊Parabramispekinensis、青梢红鲌Erythroculterdabryi和鳙也具有一定的优势特征(5

表1 崇明岛内河鱼类群落组成特征

图2 不同河段鱼类物种数、丰度、生物量(平均值+标准误)Fig.2 Number of species, abundance and biomass of the fish assemblages in the different rivers (Mean+Standard Error)柱子上方相同小写字母表示无显著性差异(P>0.05)
鲫、翘嘴红鲌和似鳊作为三个河段的共有优势种,它们的个体体长在不同河段存在明显差异,且不同种类有所不同,如图3所示。鲫的体长在南横引河显著小于北横引河和竖河(P<0.01),但在北横引河和竖河间差异不显著(P>0.05);翘嘴红鲌、似鳊在任意两河段间的体长均存在极显著差异(P<0.01),翘嘴红鲌体长排序为北横引河>竖河>南横引河,似鳊体长排序为北横引河>南横引河>竖河。

图3 主要鱼类优势种在不同河段的体长分布特征Fig.3 Body length distribution characteristics of the main dominant fish species in the different rivers
2.3 群落相似性
鱼类组成的等级聚类和nMDS分析结果如图4所示。聚类分析结果显示:崇明内河的鱼类可以分为2组:组Ⅰ包括所有南横引河站点(S1、S2、S3、S4、S5)和部分竖河站点(V3、V4、V5);组Ⅱ包括所有北横引河站点(N1、N2、N3、N4、N5)和部分竖河站点(V1、V2、V6),两大类群的相似性接近60%。nMDS分析进一步证实了这一结果(Stress=0.17 < 0.2)。相似性分析结果表明:竖河与南横引河间鱼类群落结构存在显著差异(R=0.333,P=0.013),竖河与北横引河间鱼类群落结构差异不显著(R=0.181,P=0.11),南横引河与北横引河间存在极显著差异(R=0.524,P=0.008)。
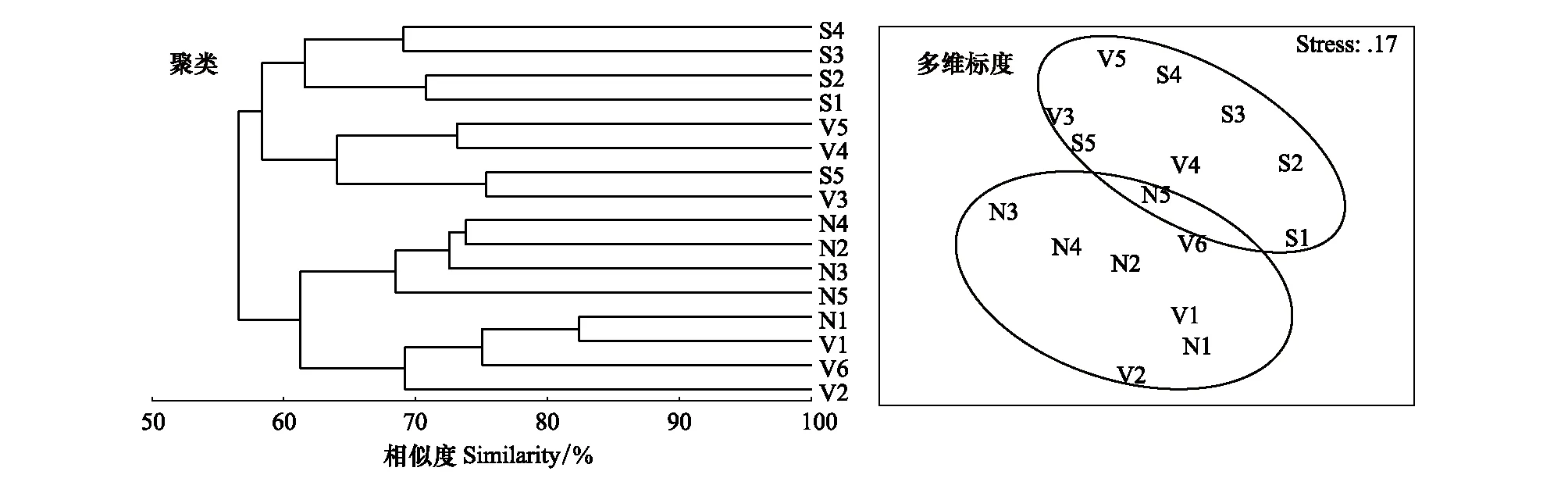
图4 各站点鱼类种类组成的等级聚类和非度量多维标度排序分析图Fig.4 Cluster and nMDS analysis on the fish assemblages in the different sites
根据以上结果,可对3类不同河道各站位物种丰度进行SIMPER分析,如表2所示。结果显示:北横引河与竖河两组间鱼类种类组成的平均相异性为39.96%,造成两组间差异的主要物种有10种。南横引河与竖河两组间鱼类种类组成的平均相异性为42.33%,造成两组间差异的主要物种有9种。南、北横引河两组间鱼类种类组成的平均相异性为43.92%,造成两组间差异的主要物种有10种。

表2 不同河道组间相异性贡献率较大的鱼种(平均贡献率>5%)
3 讨论
3.1 岛屿内河鱼类组成特征
鱼类是水域生态系统的重要组成部分,不同地区的鱼类组成往往存在一定差异。在内陆河流中,同一河流不同河段[30],或是不同的河流类型[31],大多数情况下主要鱼类均是淡水鱼,且以鲤形目鲤科鱼类居多;而在大型岛屿中,如海南岛内河的鱼类组成以鲈形目占显著优势,同时发现有多种河口鱼类[11];辽东半岛的内陆水域中,土著淡水鱼类居多,近海鱼类与半咸水鱼类约占调查区鱼类总数的1/4[32];地中海科西嘉岛河流鱼类群落的研究中,鲤科鱼类等主要的淡水鱼种在岛内较为稀缺,更多的是海洋性鱼类[33]。从本文研究结果来看,崇明岛内河鱼类组成兼具淡水种与咸水种,但淡水鱼类占优势,且以鲤科鱼类居多。
本次调查中,除了淡水鱼,还有2种洄游性鱼类和1种近海鱼类。洄游性鱼类为刀鲚和日本鳗鲡Anguillajaponica,近海鱼类为中国花鲈Lateolabraxmaculatus。一般认为,长江刀鲚包括2种生态类型,即溯河洄游型和淡水定居型[34]。刀鲚在长江口属于溯河洄游种类,是长江口水域传统的渔业捕捞对象之一,崇明岛内部水域也有相关记载[35]。日本鳗鲡为降海洄游性鱼类,秋冬季节于性成熟时顺流入海产卵,仔鱼随着洋流到达各大陆沿岸,而后在淡水环境中生活至性成熟,它们的整个生活史贯穿海洋、河口、淡水3种不同生境[36]。中国花鲈多栖息于沿海及河口区域,通常只进行深浅适温或产卵、索饵迁移, 而无长距离洄游习性;长江口是中国花鲈较佳的育肥场所,在未性成熟之前,均能在长江口育肥、生长[37]。3种不同生态型的鱼类同时分布在崇明岛内河,说明崇明岛内河环境不同于一般内陆淡水河流,而具有河口咸淡水过渡区的环境特征。
3.2 生境条件对内河鱼类分布的影响
生境是生物的个体、种群或群落生活地域的环境,包括必需的生存条件和其他对生物起作用的生态因素。崇明岛位于长江入海口,岛内水系的连通性较高,同时受长江径流和潮流作用影响,内河虽以淡水为主,但部分河段仍具备一定的盐度。而盐度是影响鱼类等水生生物的多样性及分布特征的重要因子[38- 39]。因此,崇明内河鱼类组成虽以淡水种为主,且各个河段主要优势鱼种大致相同,但也兼具近海和洄游性种类。在枯水期,咸潮入侵导致咸水包围崇明岛长达数月[40],北支倒灌盐水团的影响范围曾延伸至堡镇港[41],导致内河不同河段盐度差异较大,这也是南、北横引河鱼类分布差异较为明显的主要原因。
不同河段人为活动强度的差异对鱼类群落组成、物种多样性和分布会有明显影响[42]。研究表明,内河航运对鱼类生物多样性有直接或间接的影响。如船舶航行产生的水体扰动,会使通航水体的浊度上升,进而妨碍鱼类的生长、摄食、迁移等行为,最终影响鱼类的资源状况[43]。船舶通行过程中的物理作用如剪切力、船行波等也会导致鱼类的伤亡[44]。在本文中,不同河道鱼类种类组成和优势种的体长分布差异明显,其中,北横引河与南横引河差异最大,竖河介于二者之间。优势种似鳊对造成南、北横引河鱼类组成差异的贡献率最高,优势种鲫和翘嘴红鲌对组内平均相似性的贡献率较高。可以看出,优势鱼种是影响不同河段鱼类种类组成中的重要因素。南横引河贯穿整个崇明南部,沿途坐落多个相对发达的城镇,是崇明岛引水排涝、水土输送的大动脉。作为岛内最主要的航运通道,繁忙的航运带来的水文变化势必会影响河中鱼类的生长和发育。北横引河同样也承担着引清调水、水源调度和航运的作用,但其通航强度较低,流经区域以水产养殖区和种植区为主,所受到的人为干扰相对较小。竖向中小河道两边主要分布着农田和村镇,通行船只以小型农用船为主,作为南、北横引河的过渡和联通区,所受干扰强度介于两者之间,鱼类组成也表现出了类似特点。这表明,不同河段所处区域条件、人类干扰活动强度的差异是造成鱼类分布差异的因素之一。
有研究表明,鱼类的空间分布格局与生境异质性和稳定性密切相关[42,45]。通常,鱼类群落空间结构的完整性在相对自然的生境中表现更高,此类自然生境适宜更多的鱼类生存,其鱼类的物种数、资源量均能保持在相对较高的水平。南横引河作为岛内水运的主航道,人工岸段以石驳岸为主,护岸结构多使用安全耐久、抗冲刷的硬质材料,因此河岸硬质化明显,河道原有的底质生境被破坏,进而影响鱼类栖息地的多样性和宜居性,群落结构发生变化。而北横引河自然岸段占比较高,河道主要流经农业种植区和水产养殖区,河岸生境自然化程度高,鱼类自然栖息地保留相对完整。竖向河道生境特征介于南、北横引河之间。本文研究表明,北横引河无论在鱼类丰度还是生物量上均大于南横引河和竖向河道,主要优势种体长分布特征在不同河道也有明显差异。因此,人为干扰和生境条件在很大程度上能够改变和影响河流鱼类的群落结构。
崇明岛内河夏季鱼类群落组成特征,既遵循一般河流的规律,即生境异质性能够影响生物对空间资源的利用,同时也具备其独特性,如鱼类群落组成兼具淡水和咸水特征。除此之外,温度、电导率、DO、氨氮浓度等水环境因子也会影响鱼类群落组成及分布[46- 47],而环境因素往往会综合作用于鱼类群落结构。李艳利等人在探究影响浑太河流域鱼类群落结构的不同环境因子时发现,DO、总氮、PH和栖息地质量等显著影响了鱼类在不同河段的空间分布特征[48]。因此,未来需要在长期系统监测基础上,综合多种环境因子,对崇明岛内河鱼类组成、分布、变化趋势及内在机理开展深入研究。
参考文献(References):
[1] Karr J R. Assessment of biotic integrity using fish communities. Fisheries, 1981, 6(6): 21- 27.
[2] 叶少文, 李钟杰, 曹文宣. 牛山湖两种不同生境小型鱼类的种类组成、多样性和密度. 应用生态学报, 2007, 18(7): 1589- 1595.
[3] Sarpedonti V, Da Anunciação É M S, Bordalo A O. Spatio-temporal distribution of fish larvae in relation to ontogeny and water quality in the oligohaline zone of a North Brazilian estuary. Biota Neotropica, 2013, 13(3): 55- 63.
[4] Moss T. The governance of land use in river basins: prospects for overcoming problems of institutional interplay with the EU Water Framework Directive. Land Use Policy, 2004, 21(1): 85- 94.
[5] Grant E H C, Lowe W H, Fagan W F. Living in the branches: population dynamics and ecological processes in dendritic networks. Ecology Letters, 2007, 10(2): 165- 175.
[6] Benda L, Poff N L, Miller D, Dunne T, Reeves G, Pess G, Pollock M. The network dynamics hypothesis: how channel networks structure riverine habitats. BioScience, 2004, 54(5): 413- 427.
[7] Osborne L L, Wiley M J. Influence of tributary spatial position on the structure of warmwater fish communities. Canadian Journal of Fisheries and Aquatic Sciences, 1992, 49(4): 671- 681.
[8] Grenouillet G, Pont D, Hérissé C. Within-basin fish assemblage structure: the relative influence of habitat versus stream spatial position on local species richness. Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(1): 93- 102.
[9] Nakagawa H. Contribution of environmental and spatial factors to the structure of stream fish assemblages at different spatial scales. Ecology of Freshwater Fish, 2014, 23(2): 208- 223.
[10] 王晓丽, 王嫒, 石洪华, 郑伟, 周然. 海岛陆地生态系统固碳估算方法. 生态学报, 2014, 34(1): 88- 96.
[11] 余梵冬, 王德强, 顾党恩, 胡隐昌, 朱海, 佟延南, 李芳远, 申志新. 海南岛南渡江鱼类种类组成和分布现状. 淡水渔业, 2018, 48(2): 58- 67.
[12] 李红敬, 张凤瑞. 海南森林溪流淡水鱼类区系研究. 信阳师范学院学报: 自然科学版, 2001, 14(2): 192- 194.
[13] 陈宜瑜, 何舜平. 海峡两岸淡水鱼类分布格局及其生物地理学意义. 自然科学进展, 2001, 11(4): 337- 342.
[14] 张涛, 庄平, 刘健, 章龙珍, 冯广朋, 侯俊利, 赵峰, 刘鉴毅. 长江口崇明东滩鱼类群落组成和生物多样性. 生态学杂志, 2009, 28(10): 2056- 2062.
[15] 刘凯, 徐东坡, 张敏莹, 段金荣, 施炜纲. 崇明北滩鱼类群落生物多样性初探. 长江流域资源与环境, 2005, 14(4): 418- 421.
[16] 王敏, 熊丽君, 黄沈发. 崇明生态岛建设生态环境指标体系研究. 上海环境科学, 2010, 29(2): 47- 51.
[17] 池源, 石洪华, 丰爱平. 典型海岛景观生态网络构建——以崇明岛为例. 海洋环境科学, 2015, 34(3): 433- 440.
[18] 张颖纯, 姜德刚, 李建华. 崇明岛“闸控型”河网水体富营养化特征及其影响因素. 湖泊科学, 2013, 25(3): 366- 372.
[19] 林发永. 崇明岛水系改造的几点设想. 水利水电快报, 2003, 24(8): 15- 16.
[20] 姜德刚, 李建华, 徐金燕, 张琳婷. 崇明岛富营养化河道溶解有机质的三维荧光光谱特征. 水生态学杂志, 2019, 40(3): 33- 40.
[21] 钱壁君. 崇明岛南横引河多种功能综合利用河道设计. 山西水利, 2018, (11): 19- 21.
[22] 徐东. 浅谈崇明南横引河综合整治. 吉林水利, 2005, (2): 34- 35.
[23] 朱英杰, 李建华, 姜德刚. 崇明河网水质污染的空间差异性分析. 安徽农业科学, 2010, 38(22): 11906- 11908, 11912- 11912.
[24] 中国水产科学研究院东海水产研究所, 上海市水产研究所. 上海鱼类志. 上海: 上海科学技术出版社, 1990: 111- 310.
[25] 安传光, 赵云龙, 林凌, 吕敢堂, 陈亚瞿. 长江口九段沙潮间带大型底栖动物季节分布特征的初步研究. 水产学报, 2007, 31(S1): 52- 58.
[26] Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Applied and Environmental Microbiology, 2005, 71(12): 8228- 8235.
[27] Kruse M, Taylor M H, Muhando C A, Reuter H. Lunar, diel, and tidal changes in fish assemblages in an East African marine reserve. Regional Studies in Marine Science, 2016, 3: 49- 57.
[28] 张衡, 朱国平. 长江河口潮间带鱼类群落的时空变化. 应用生态学报, 2009, 20(10): 2519- 2526.
[29] 祁海萍, 申希平. Kruskal-Wallis H检验平均秩多重比较在SPSS软件中的实现. 兰州工业学院学报, 2015, 22(2): 76- 78.
[30] 林鹏程, 李淑贞, 秦祥朝, 程卫习, 刘焕章. 黄河伊洛河中下游鱼类多样性及群落结构. 湖泊科学, 2019, 31(2): 482- 492.
[31] 张春光, 赵亚辉, 邢迎春, 郭瑞禄, 张清, 冯云, 樊恩源. 北京及其邻近地区野生鱼类物种多样性及其资源保育. 生物多样性, 2011, 19(5): 597- 604.
[32] 牟秀林. 辽东半岛及其沿海岛屿的淡水鱼类区系. 大连海洋大学学报, 1990, 5(1): 22- 32.
[33] Changeux T. Insular characteristics of freshwater fish communities in the island of Corsica, comparison with French continental coastal rivers. Italian Journal of Zoology, 1998, 65(S1): 305- 311.
[34] Liu D, Li Y Y, Tang W Q, Yang J Q, Guo H Y, Zhu G L, Li H H. Population structure ofCoilianasusin the Yangtze River revealed by insertion of short interspersed elements. Biochemical Systematics and Ecology, 2014, 54: 103- 112.
[35] 管卫兵, 陈辉辉, 丁华腾, 宣富君, 戴小杰. 长江口刀鲚洄游群体生殖特征和条件状况研究. 海洋渔业, 2010, 32(1): 73- 81.
[36] 汪德海, 龚小玲, 鲍宝龙, 刘利平. 长江口降海洄游日本鳗鲡的生长特性. 上海海洋大学学报, 2016, 25(1): 25- 33.
[37] 孙帼英, 朱云云, 周忠良, 陈建国. 长江口及浙江沿海花鲈的繁殖生物学. 水产学报, 1994, 18(1): 18- 23.
[38] Gain A K, Uddin M N, Sana P. Impact of river salinity on fish diversity in the south-west coastal region of Bangladesh. International Journal of Ecology and Environmental Sciences, 2008, 34(1): 49- 54.
[39] 吴逢润, 童春富. 近30年长江口北支演变及其对物种多样性的影响. 海洋学报, 2017, 39(2): 72- 85.
[40] 孔亚珍, 贺松林, 丁平兴, 胡克林. 长江口盐度的时空变化特征及其指示意义. 海洋学报, 2004, 26(4): 9- 18.
[41] 恽才兴. 长江河口近期演变基本规律. 北京: 海洋出版社, 2004: 103- 105.
[42] 杜龙飞, 徐建新, 李彦彬, 渠晓东, 刘猛, 张敏, 余杨. 北京市主要河流鱼类群落的空间格局特征. 环境科学研究, 2019, 32(3): 447- 457.
[43] Zajicek P, Wolter C. The effects of recreational and commercial navigation on fish assemblages in large rivers. Science of the Total Environment, 2019, 646: 1304- 1314.
[44] Gutreuter S, Dettmers J M, Wahl D H. Estimating mortality rates of adult fish from entrainment through the propellers of river towboats. Transactions of the American Fisheries Society, 2003, 132(4): 646- 661.
[45] 王晓宁, 彭世贤, 张亚, 张远, 钱昶, 丁森. 滦河流域鱼类群落结构空间异质性与影响因子分析. 环境科学研究, 2018, 31(2): 273- 282.
[46] Furlan N, Esteves K E, Quináglia G A. Environmental factors associated with fish distribution in an urban neotropical river (Upper Tietê River Basin, São Paulo, Brazil). Environmental Biology of Fishes, 2013, 96(1): 77- 92.
[47] 王伟, 王冰, 何旭颖, 渠晓东, 张远. 太子河鱼类群落结构空间分布特征. 环境科学研究, 2013, 26(5): 494- 501.
[48] 李艳利, 李艳粉, 徐宗学. 影响浑太河流域鱼类群落结构的不同尺度环境因子分析. 环境科学, 2014, 35(9): 3504- 3512.
猜你喜欢
化工管理(2022年13期)2022-12-02
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
中华建设(2020年5期)2020-07-24
石油沥青(2020年1期)2020-05-25
安徽农业科学(2017年18期)2017-07-10
钟山风雨(2017年1期)2017-03-12
绿色科技(2014年1期)2014-11-28
青年文摘·上半月(1984年12期)1984-11-01
