
6种杀线剂对重茬山药土壤微生物数量、酶活性和养分含量的影响
2021-04-01 07:58:14范琳娟刘子荣徐雪亮王奋山彭德良姚英娟
浙江农业学报 2021年3期
范琳娟,刘子荣,徐雪亮,王奋山,彭德良,姚英娟,*
(1.江西省农业科学院 农业应用微生物研究所,江西 南昌330200; 2.中国农业科学院 植物保护研究所,北京 100094)
山药具有较高的食药两用价值,综合种植效益远高于很多经济作物[1]。然而,近些年山药上植物寄生线虫的发生越来越严重,在河南、山东、河北、江苏、江西等地的山药上均发现有植物寄生线虫严重为害的报道,导致许多山药主产区出现了明显的连作障碍[2-5]。研究表明,为害山药的植物寄生线虫主要为根结线虫和短体线虫[3,6-8]。为克服植物寄生线虫对山药的为害,缓解连作障碍效应,施用杀线剂是目前生产中常采用的防治方式[2,9-11];但杀线剂在杀死土壤中植物寄生线虫的同时,也可能会杀死土壤中的有益生物,破坏土壤的生态平衡[12-13]。
土壤微生物在土壤生态系统的物质循环和能量流动过程中起着重要作用,其数量变化可作为评价土壤质量、维持土壤肥力和作物生产力的重要指标之一[14]。土壤酶主要来自微生物、动植物活体的分泌或动植物残体、遗骸分解释放,不仅可用来评价土壤的肥力状况,也是衡量土壤生态系统土壤质量变化的敏感指标[15];因此,土壤微生物数量和酶活性常被用来判断土壤生态环境是否受到干扰或污染[16]。目前,大多数关于杀线剂的研究主要关注其对植物寄生线虫的防治效果,鲜有研究重视其对土壤生态环境的影响。时立波等[12]发现,阿维菌素、涕灭威和克线丹对土壤微生物的组成和多样性均有不同程度的影响;邹小明等[17]研究表明,三唑磷对土壤细菌、真菌和放线菌在培养初期均有不同程度的抑制作用;武贺等[18]研究发现,施用呋喃丹和涕灭威会降低土壤中真菌的种类,影响土壤真菌的动态平衡。上述杀线剂除阿维菌素和三唑磷外均为高毒性农药,而目前关于常用的阿维·噻唑膦和氟吡菌酰胺等低毒性化学杀线剂和淡紫拟青霉等生物杀线剂对土壤环境的影响尚未见报道。基于此,本研究以山药苗期和成熟期的重茬土壤为研究对象,探讨2种化学杀线剂(阿维·噻唑膦和氟吡菌酰胺)和4种生物杀线剂(厚孢轮枝菌、淡紫拟青霉、辣根素和茶枯抑线生物菌肥)对土壤微生物种群数量、酶活性和土壤肥力的影响,旨在明确其对土壤生态环境的影响,以期为上述6种杀线剂的合理采用和环境毒性评价提供依据。
1 材料与方法
1.1 供试材料
供试土样采自瑞昌市范镇镇范镇村重茬多年山药田。
供试药品:10.5%阿维·噻唑膦颗粒剂(AG)/10%阿维·噻唑膦微乳剂(AM),燕化永乐(乐亭)生物科技有限公司;41.7%氟吡菌酰胺悬浮剂(Fl),拜耳作物科学(中国)有限公司;2.5亿孢子·g-1厚孢轮枝菌颗粒剂(Vc),广东真格生物科技有限公司;50亿·g-1淡紫拟青霉颗粒剂(Pl),湖北启明生物工程有限公司;20%辣根素水剂(Ho),北京亚戈农生物药业公司;10亿·g-1茶枯抑线生物菌肥(解淀粉芽孢杆菌)(tc),广东真格生物科技有限公司。
1.2 试验设计
试验地山药播种密度为8株·m-2,株行距(20~30)cm×(45~55)cm。小区面积10 m2(2 m×5 m),每小区种4行,每行20株,每小区种80株。每处理重复3次。按照正常操作进行田间农事管理。
试验共设置7个处理,各处理的药剂分别于2019年3月11日和2019年7月18日(第一次取样前)按照推荐剂量施用(表1)。第一次用药时,颗粒剂拌入适量细土撒施;其他剂型按照0.75 L·m-2兑水稀释后沟内浇施;辣根素沟内稀释浇施后,迅速盖土并用塑料薄膜封闭,熏蒸处理7 d并散气4 h后再进行播种。除辣根素外,其他处理均于第一次施药后当天播种。第2次用药时,均按照0.80 L·m-2兑水稀释浇施。空白对照(CK)不用药。
1.3 试验方法
1.3.1 土样采集与处理
试验设计2次田间采样,分别在山药苗期(2019年7月18日)和成熟期(2019年11月20日),其中第一次采样时土壤含水量在20%左右(7月11日曾降雨),第二次采样与第一次采样期间降雨极少,至第二次采样时土壤含水量在10%左右。采用随机5点取样法,采集5~20 cm深度土样,每个处理3个重复,每个重复取约500 g放入自封袋,带回实验室。一部分放于4 ℃保存,用于土壤微生物数量测定分析;另一部分自然风干研磨后过2 mm筛,用于测定土壤酶活性和理化性质。
1.3.2 测定指标与方法
土壤pH值,按水土体积质量比2.5∶1的比例浸提后,用酸度计法测定;有机质含量采用重铬酸钾法测定;铵态氮含量采用靛酚蓝比色法测定;速效钾含量采用火焰光度法测定;速效磷含量采用钼锑抗比色法测定[19]。
土壤微生物种群数量测定:采用稀释平板计数法,计算每克干土中微生物的数量。其中,细菌培养用牛肉膏蛋白胨培养基,真菌培养用马铃薯葡萄糖培养基,放线菌培养用改良高氏一号培养基。
土壤酶活性测定:土壤脲酶活性采用苯酚钠-次氯酸钠比色法测定,酸性磷酸酶活性采用磷酸苯二钠比色法测定,蔗糖酶活性采用3,5-二硝基水杨酸比色法测定[19]。
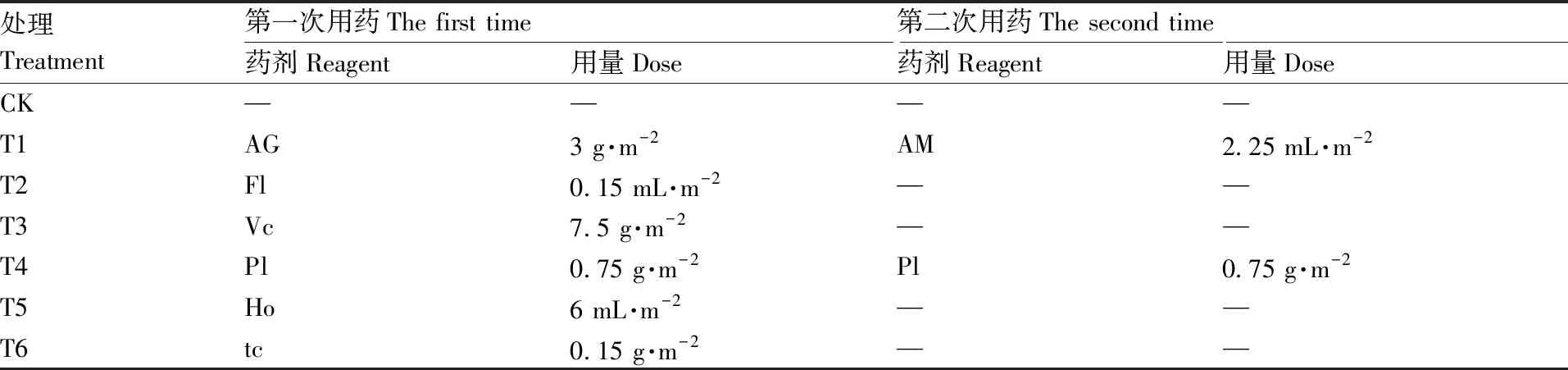
表1 各处理的供试药剂与施药量
1.4 数据处理
所得数据用Excel 2013进行处理,用SPSS 20.0进行单因素方差分析(ANOVA)。
2 结果与分析
2.1 不同处理对重茬山药土壤微生物种群数量的影响
如图1-A所示:施用2种化学杀线剂对苗期土壤的细菌数量表现出显著(P<0.05)的促进作用,分别较CK提高182.35%和152.94%;而至成熟期时,仅施用茶枯抑线生物菌肥的土壤细菌数量较CK显著(P<0.05)提高,增幅达233.33%,其他处理均与CK无显著差异。
如图1-B所示:施用2种化学杀线剂和茶枯抑线生物菌肥的苗期土壤真菌数量显著(P<0.05)高于CK,增幅分别为180.00%、130.00%和90.00%;成熟期时,施用氟吡菌酰胺的土壤真菌数量显著(P<0.05)低于CK,降幅为61.54%,而施用辣根素的土壤真菌数量显著(P<0.05)高于CK,增幅达84.61%,其他处理与CK无显著差异。
由图1-C可知:苗期时,施用淡紫拟青霉的土壤细菌真菌比(细菌/真菌)显著(P<0.05)高于CK,增幅达72.86%,而施用茶枯抑线生物菌肥的土壤细菌/真菌较CK显著(P<0.05)降低87.54%;成熟期时,施用氟吡菌酰胺和茶枯抑线生物菌肥的土壤细菌/真菌较CK显著(P<0.05)提高,增幅分别达61.29%和186.29%。
如图1-D所示:苗期时,不同杀线剂对土壤放线菌数量无显著影响;成熟期时,施用辣根素的土壤放线菌数量显著(P<0.05)高于CK,增幅达141.67%。
2.2 不同处理对重茬山药土壤酶活性的影响
2.2.1 不同处理对土壤脲酶活性的影响
如图2所示:不同杀线剂对重茬土壤脲酶活性在山药苗期和成熟期均无显著影响,推测施用杀线剂并不会显著影响山药土壤氮素的转化过程。
2.2.2 不同处理对土壤酸性磷酸酶活性的影响
如图3所示:在山药苗期时,施用阿维·噻唑膦、氟吡菌酰胺、厚孢轮枝菌、淡紫拟青霉和辣根素后,土壤酸性磷酸酶活性均较CK显著(P<0.05)升高,幅度分别为2.04%、2.98%、98.35%、104.79%和3.14%,但施用茶枯抑线生物菌肥后,土壤酸性磷酸酶活性与CK无显著差异;在山药成熟期时,施用6种杀线剂后,土壤酸性磷酸酶活性均显著(P<0.05)低于CK,且降幅均在40%以上。
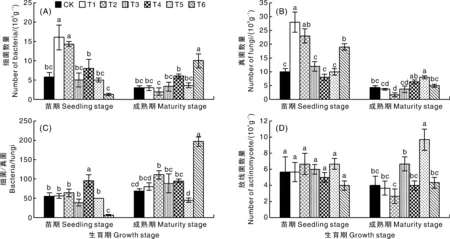
同一生育期柱上无相同字母的表示处理间差异显著(P<0.05)。下同。Bars marked without the same letters at the same growth stage indicated significant difference within treatments at P<0.05. The same as below.图1 不同处理对山药土壤微生物数量的影响Fig.1 Effects of different treatments on quantity of soil microorganisms in replanted Chinese yam field

图2 不同处理对重茬山药土壤脲酶活性的影响Fig.2 Effects of different treatments on soil urease activity in replanted Chinese yam field
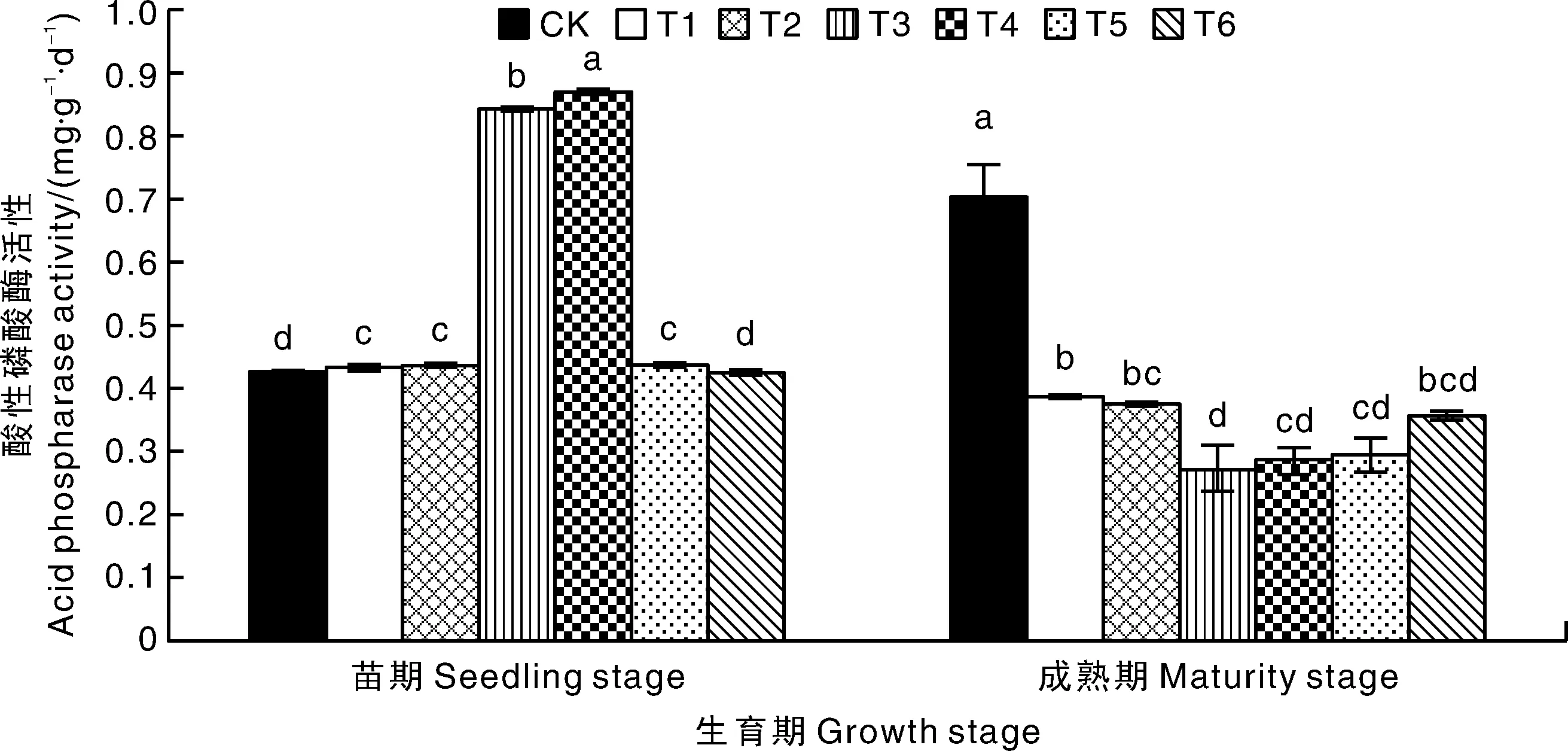
图3 不同处理对重茬山药土壤酸性磷酸酶活性的影响Fig.3 Effects of different treatments on soil acid phosphatase activity in replanted Chinese yam field
2.2.3 不同处理对土壤蔗糖酶活性的影响
如图4所示,在山药苗期时,施用6种杀线剂后,土壤蔗糖酶活性均较CK显著(P<0.05)降低,降幅在23.04%~70.99%;在山药成熟期,施用茶枯抑线生物菌肥的土壤蔗糖酶活性较CK显著(P<0.05)提高121.65%,但施用其他5种杀线剂的土壤蔗糖酶活性均较CK显著(P<0.05)降低,且降幅均在30%以上。

图4 不同处理对山药重茬土壤蔗糖酶活性的影响Fig.4 Effects of different treatments on soil sucrase activity in replanted Chinese yam field
2.3 不同处理对重茬山药土壤养分的影响
由表2、表3可知,在山药苗期和成熟期,不同杀线剂处理的土壤pH值和有机质含量与CK均无显著差异。与CK相比,不同杀线剂对苗期土壤铵态氮和速效钾含量均无显著影响,但6种杀线剂处理的苗期土壤速效磷含量均显著(P<0.05)低于CK;至成熟期后,阿维·噻唑膦、氟吡菌酰胺、厚孢轮枝菌、淡紫拟青霉和辣根素处理的土壤铵态氮含量均显著(P<0.05)高于CK,增幅均在140%以上,6种杀线剂处理的土壤速效钾含量也均大幅显著(P<0.05)提高,且增幅均在30%以上。

表2 不同处理对苗期山药土壤养分的影响

表3 不同处理对成熟期山药土壤养分的影响
3 结论与讨论
土壤细菌、真菌和放线菌是土壤微生物区系的重要组成成分,其数量直接影响着土壤生化活性,以及土壤养分的组成与转化,且大多数土壤真菌是重要的病原菌[20]。研究表明,土壤中细菌和放线菌数量越多,真菌数量越少,细菌/真菌越大,土壤越有利于植物生长[21-22]。本研究结果表明,2种化学杀线剂施用后对苗期土壤的细菌和真菌数量均表现出显著的促进作用,而对苗期土壤的放线菌数量、细菌/真菌,以及成熟期的土壤细菌、放线菌数量无显著影响。研究显示,氟吡菌酰胺对土壤细菌、真菌和放线菌数量的抑制作用达60 d左右,阿维·噻唑膦在土壤中的持效期不会超过90 d[23-24]。本研究中,2次取样时间与施药的间隔期均超过了4个月。据此分析,本研究中2种化学杀线剂在试验的过程中可能被土壤中的一些微生物当作碳源利用[25],降解产物在土壤中积累,由此导致苗期土壤细菌和真菌数量大幅提升,至成熟期后,由于降解产物基本已被消耗,因此又降至CK水平或更低。4种生物杀线剂中,茶枯抑线生物菌肥对土壤微生物的影响相对较大,淡紫拟青霉和厚孢轮枝菌的影响相对较小。茶枯抑线生物菌肥(解淀粉芽孢杆菌)是近年新研发出的集药、肥于一体的全新生物型颗粒肥,具备双重防线机理(茶枯中的茶皂素和解淀粉芽孢杆菌),并添加有多种微量元素。一般情况下,微生物菌剂施入土壤后可迅速繁殖,然后通过分解土壤中的矿物质、增加土壤中的抗菌物质等,在一定程度上抑制土传病害,并降低其种群数量。本研究结果表明,土壤中的细菌在山药苗期很难适应茶枯抑线生物菌肥(解淀粉芽孢杆菌)的毒性干扰,导致其种群数量受到了限制,而为真菌的生长提供了生存空间;但至成熟期后,土壤中的细菌已经逐渐适应了这种干扰,细菌种群数量大幅度提升,真菌种群数量逐渐恢复到CK水平。也正因如此,土壤中的细菌/真菌在苗期和成熟期先后表现出抑制和促进的作用。这表明,茶枯抑线生物菌肥(解淀粉芽孢杆菌)可在一定程度上改善土壤环境和质量,促使连作引起的“真菌型”土壤向“细菌型”转变,缓解作物的连作障碍。胡基华等[26]研究发现,解淀粉芽孢杆菌TF28施入土壤8周对土壤细菌数量无显著影响,而对放线菌和真菌数量却表现出促进作用。这与本研究的结果略有不同。考虑到不同作物的根际都有其特定的微生物群落,即使是同一作物在不同生育期和营养状态下,其根际土壤微生物群落也不会完全相同[27];因此,不同研究的确可能会得到不完全一致的结果。总的来看,在本研究中,无论是化学杀线剂还是生物杀线剂,对土壤微生物群落结构均会产生一定的影响,其中,茶枯抑线生物菌肥(解淀粉芽孢杆菌)的处理可在一定程度上起到调节土壤微生物群落结构、改善土壤环境的作用。
土壤酶可参与土壤中一切复杂的生化过程,土壤微生物代谢是其重要来源之一;因此,影响土壤微生物的因子也会影响土壤酶活性的变化,且不同微生物和土壤酶活性对不同药剂的敏感性也不尽相同[28]。土壤脲酶主要催化酰胺化合物水解,生成的氨是植物氮素营养的重要来源之一。纪春涛等[29]研究表明,100 d时不同剂量噻唑膦处理区的土壤脲酶活性均基本恢复到对照水平。这与本研究结果基本一致,表明土壤脲酶一般情况下对外来干扰不敏感。
土壤酸性磷酸酶是评价土壤磷元素生物转化强度和方向的指标,主要促进土壤中有机磷转化为植物可直接吸收利用的无机磷。山药苗期时,阿维·噻唑膦、氟吡菌酰胺、厚孢轮枝菌、淡紫拟青霉和辣根素对土壤酸性磷酸酶活性均表现为促进作用,茶枯抑线生物菌肥对其无显著影响,但至成熟期后,6种杀线剂均对土壤酸性磷酸酶活性表现出显著的抑制作用。纪春涛[30]研究发现,不同剂量噻唑膦施用50 d后,对室内土壤磷酸酶活性的抑制作用逐渐减弱。本研究结果与其不同,这可能与不同作物根际微生物群落的差异有关。氟吡菌酰胺、厚孢轮枝菌、淡紫拟青霉、辣根素不仅对植物线虫具有毒杀作用,对一些土传病害也具有较好的防治效果。据此分析,这4种杀线剂施用后破坏了土壤中原有微生物群落的平衡,对土壤中的一些病原菌产生了毒杀作用,为可代谢产生土壤酸性磷酸酶的微生物种群提供了生存空间,抑或是氟吡菌酰胺或辣根素产生的降解物质给可代谢产生土壤酸性磷酸酶的微生物种群提供了食物来源,因此提高了苗期土壤酸性磷酸酶的活性。这与傅丽君等[31]的研究结果一致。至山药成熟期后,6种杀线剂对土壤酸性磷酸酶活性表现出强烈的抑制作用,其原因不详,有待进一步研究。
土壤蔗糖酶主要参与水解蔗糖,生成的葡萄糖和果糖可被植物和微生物直接利用。蔗糖酶活性高,表明土壤生物活性较高,土壤状况良好。本研究结果表明,除茶枯抑线生物菌肥外,其他5种杀线剂对土壤蔗糖酶活性均表现出显著的抑制作用。仉欢等[32]研究表明,高浓度的磷化铝对土壤蔗糖酶活性的抑制作用较大。贾红梅等[33]发现,34株丛枝菌根(AM)真菌在施用150 d后,虽然大多数AM真菌对土壤蔗糖酶活性表现出显著的促进作用,但仍有12株表现出显著的抑制作用。这表明土壤蔗糖酶对不同药剂的敏感性不同。阿维·噻唑膦等5种杀线剂对土壤蔗糖酶均具有一定的毒性效应;而对于茶枯抑线生物菌肥(解淀粉芽孢杆菌)来说,土壤蔗糖酶活性在苗期时似乎很难适应其干扰,酶活性受到了抑制,但至成熟期后不仅适应了这种影响,还能在一定程度上利用菌肥的影响激活土壤蔗糖酶的活性[32]。综上,这6种杀线剂施用后主要影响土壤酸性磷酸酶和蔗糖酶活性,干扰土壤中有机磷矿化和碳循环,但并不会影响作物对土壤中氮元素的吸收。
合理的土壤养分是保证作物良好生长的前提。本研究结果表明,6种杀线剂主要影响土壤铵态氮、速效钾和速效磷含量。铵态氮和速效钾含量在苗期时与对照相比均无显著变化,至成熟期后均在一定程度上高于对照,而速效磷含量在苗期表现为显著的抑制作用。有研究表明,药剂施入土壤后,可通过改变土壤微生物的代谢途径影响土壤酶活性,从而对土壤正常的养分循环过程造成干扰[34]。由此分析,可能是这6种杀线剂施用后对土壤微生物群落和酶活性产生了影响,由此改变了土壤中铵态氮、速效钾和速效磷的含量。
综上,本试验所选择的6种杀线剂施用后对土壤微生物、土壤脲酶和土壤养分的影响随着时间的延长均可逐渐恢复,但其对土壤酸性磷酸酶和蔗糖酶活性的影响持续时间较长,可能会干扰土壤中有机磷矿化和碳循环,影响土壤中有效养分的释放,对土壤生态环境产生不利影响。山药不同生育期对土壤养分的需求不同,其中,苗期和成熟期对氮肥和钾肥的需求量较大,对磷肥需求不多,据此推测酸性磷酸酶活性和蔗糖酶活性的降低可能对山药产量影响较小。对比发现,茶枯抑线生物菌肥(解淀粉芽孢杆菌)虽然会在一定程度上抑制土壤酶的活性,但其也可在一定程度上调节土壤微生物群落结构,改善连作土壤以”真菌型”为主的状态,促使土壤向“细菌型”转变。
本文仅从土壤微生物数量、酶活性和土壤养分这3个方面初步分析了这6种杀线剂对土壤生态环境的影响,但土壤微生态环境非常复杂,土壤微生物多样性对药剂的安全性评估具有重要作用,这方面的研究还有待进一步深入,以便更全面地评价这6种杀线剂对土壤生态环境的影响。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:28
房地产导刊(2020年9期)2020-10-28 08:37:24
中国果树(2020年2期)2020-07-25 02:14:18
今日农业(2019年12期)2019-08-15 00:56:32
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
新疆农垦科技(2016年2期)2016-08-21 13:50:28
广西林业科学(2016年1期)2016-03-20 05:33:00
浙江柑橘(2016年4期)2016-03-11 20:13:01
现代农业(2016年4期)2016-02-28 18:42:15
医学研究杂志(2015年8期)2015-06-22 14:00:57
