
云顶山不同人工林林下植物多样性及其与土壤理化性质的关系
2021-03-31 06:44李婷婷唐永彬周润惠余飞燕董洪君郝建锋
生态学报 2021年3期
李婷婷, 唐永彬, 周润惠, 余飞燕, 董洪君, 王 敏, 郝建锋,3,*
1 四川农业大学林学院, 成都 611130 2 四川省资阳市自然资源和规划局, 资阳 641300 3 四川农业大学水土保持与荒漠化防治重点实验室, 成都 611130
物种多样性能反映物种水平上的生物多样性[1- 2],其维持机制一直是生态学的研究热点[3- 4]。林下植被作为森林生态系统的重要组成部分,在促进物种多样性发展、维持系统稳定性、调控物质能量交换等方面发挥着重要作用[5- 6]。如Wu等[7]对中国南方桉树人工林的研究发现,人为去除林下灌草层会导致土壤微生物群落结构改变、凋落物分解减缓,最终必将影响人工林的养分循环及土壤碳排放。Nilsson等[8]对瑞典北方森林的研究表明,林下植被是生态系统的主要驱动力,其短期内影响着森林树种的组成,长期来看则是土壤肥力的主要驱动因素,影响着养分有效性及植物生长。
我国人工林面积居于世界首位[3,9- 10],其在我国森林可持续经营管理中发挥着重要作用[11]。设计完善、多目标经营管理的人工林能够恢复部分天然林所具有的生态系统服务功能[12]。而林分树种的正确选择则是实现人工林近自然林经营的重要基础。因为林分类型不同会使林冠结构、养分循环、林下微环境等产生差异,最终将导致林下植物组成及其多样性格局变异[13- 14]。因此研究不同人工林林下植物多样性差异及其维持机制,对维持人工林可持续经营具有重要意义。
植物-环境关系一直是生态学研究的重要方面[15]。环境因子如何影响林下植物分布并导致植物多样性格局变异是生态学家们长期致力研究的问题[16]。影响林下植物多样性的环境因子包括生物因素与非生物因素两个方面,生物因素如林龄[11]、林分密度[9]、人为干扰[1]等,非生物因素如气候条件[16]、地形条件[17]、土壤条件[16]等。其中土壤因素被广泛认为是中小尺度范围内制约植物多样性的主要方面[11,18- 19]。如Weigel等[16]对欧洲山毛榉林下植物的研究发现,相较于气候因素,土壤因素对林下植物分布的影响力更大。并且随着现代排序方法的成熟可以定量研究各土壤因子对植物多样性的单独影响[11,19],但不同地区制约多样性的土壤因子可能不同[4],因此在区域尺度上揭示影响林下植物多样性的土壤驱动因子对理解森林生态系统结构与功能具有重要意义。
目前关于人工林林下植物多样性及其与土壤理化性质关系的研究还较少,且已有研究多集中于林分密度[9]、林龄[11]、林窗[10]等对林下植物多样性及土壤理化性质的影响,而不同人工林类型方面的研究还少有报道。有鉴于此,以成都云顶山广泛分布的5种人工林为研究对象,试图解决以下几个问题:(1)不同人工林林下植物组成;(2)不同人工林林下植物多样性;(3)不同人工林土壤理化性质;(3)影响林下植物多样性的主要土壤因子有哪些?通过以上问题的解决,以期为相关理论研究提供一定数据支撑,并为当地及类似区域人工林经营管理与物种多样性保育提供理论参考。
1 材料与方法
1.1 研究区域概况
图1 云顶山地理位置Fig.1 The geographic location of Yunding Mountain
云顶山(30°29′―30°57′N,104°20′―104°52′E)属成都平原龙泉山脉中段(图1),位于成都市金堂县境内,面积约67 km2,平均海拔982 m。该区属亚热带湿润季风气候,年均日照1295.5 h,年均气温16.6℃,年均无霜期285 d,常年雨量充沛,年均降水量920.5 mm。土壤较肥沃,以黄壤与紫色土为主。植被属人工种植的生态公益林。
20世纪50年代末,受“大办钢铁”、“大办公共食堂”等影响,民众“大砍大伐”、“毁林开荒”等行为使云顶山森林植被受到持续严重破坏,到60年代初期,其森林覆被已很少。1963—1965年,为贯彻执行“调整、巩固、充实、提高”方针,四川省委、省政府多次强调必须植树造林,因此金堂县地方政府组织民众开始了大规模的植树造林活动。云顶山树种栽植的初始密度较高,约为1100株/hm2,主要树种包括:柏木(Cupressusfunebris)、银杏(Ginkgobiloba)、枫杨(Pterocaryastenoptera)、桤木(Alnuscremastogyne)、光皮梾木(Swidawilsoniana)、灯台(Cornuscontroversa)等,多以不同树种搭配混合栽植。后期由当地林业站组织人员进行管护,属生态公益林,禁伐。
经长期自然演替,云顶山植被类型丰富,以不同树种混交而成的常绿阔叶林、常绿针叶林、常绿针阔混交林为主,同时乔木层出现了构树(Broussonetiapapyrifera)、珊瑚朴(Celtisjulianae)、枇杷(Eriobotryajaponica)、八角枫(Alangiumchinense)等对当地环境适生良好的乡土树种,人工林群落在向次生林群落方向演替。区内林下植物丰富,主要灌木有荚蒾(Viburnumdilatatum)、石海椒(Reinwardtiaindica)、蚊母树(Distyliumracemosum)、五叶地锦(Parthenocissusquinquefolia)等;主要草本植物有韩信草(Scutellariaindica)、细野麻(Boehmeriagracilis)、莩草(Setariachondrachne)、过路黄(Lysimachiachristinae)等。
1.2 植被调查、采样及样品分析
2018年9月,依据方精云等[20]的研究方法,对云顶山进行实地全面踏查,采用典型抽样法选取研究区广泛分布的5种人工林(图2):柏木枫杨林(Ⅰ)、银杏楠木林(Ⅱ)、光皮梾木香樟林(Ⅲ)、枫杨桤木林(Ⅳ)、柏木纯林(Ⅴ)。不同人工林各设置4个20 m×30 m的样地,共计样地20个(表1),总面积12000 m2,样地选择尽量遵循立地条件基本相似、林相整齐、林层丰富、具有代表性原则。灌木、草本样方设置方法及数据记录内容见张柳桦等[3]。
土壤理化性质测定指标包括:pH值、含水量、容重、有机质、全氮、全磷、全钾、速效磷、速效钾,土壤样品采集、处理及测定方法见王媚臻等[9]。
1.3 数据处理及分析
1.3.1重要值及物种多样性指数计算
植物多样性测定采用物种丰富度指数(D)、Shannon-wiener多样性指数(H)、Simpson优势度指数(H′)和Pielou均匀度指数(Jsw),灌草层重要值及物种多样性指数计算公式见王媚臻等[9]。
1.3.2数量分析方法
采用DPS 7.05中的单因素方差分析法(One-Way ANOVA)分析不同人工林物种多样性指数及土壤理化性质的差异(P=0.05),运用Excel 2010统计软件进行数据整理与统计分析。
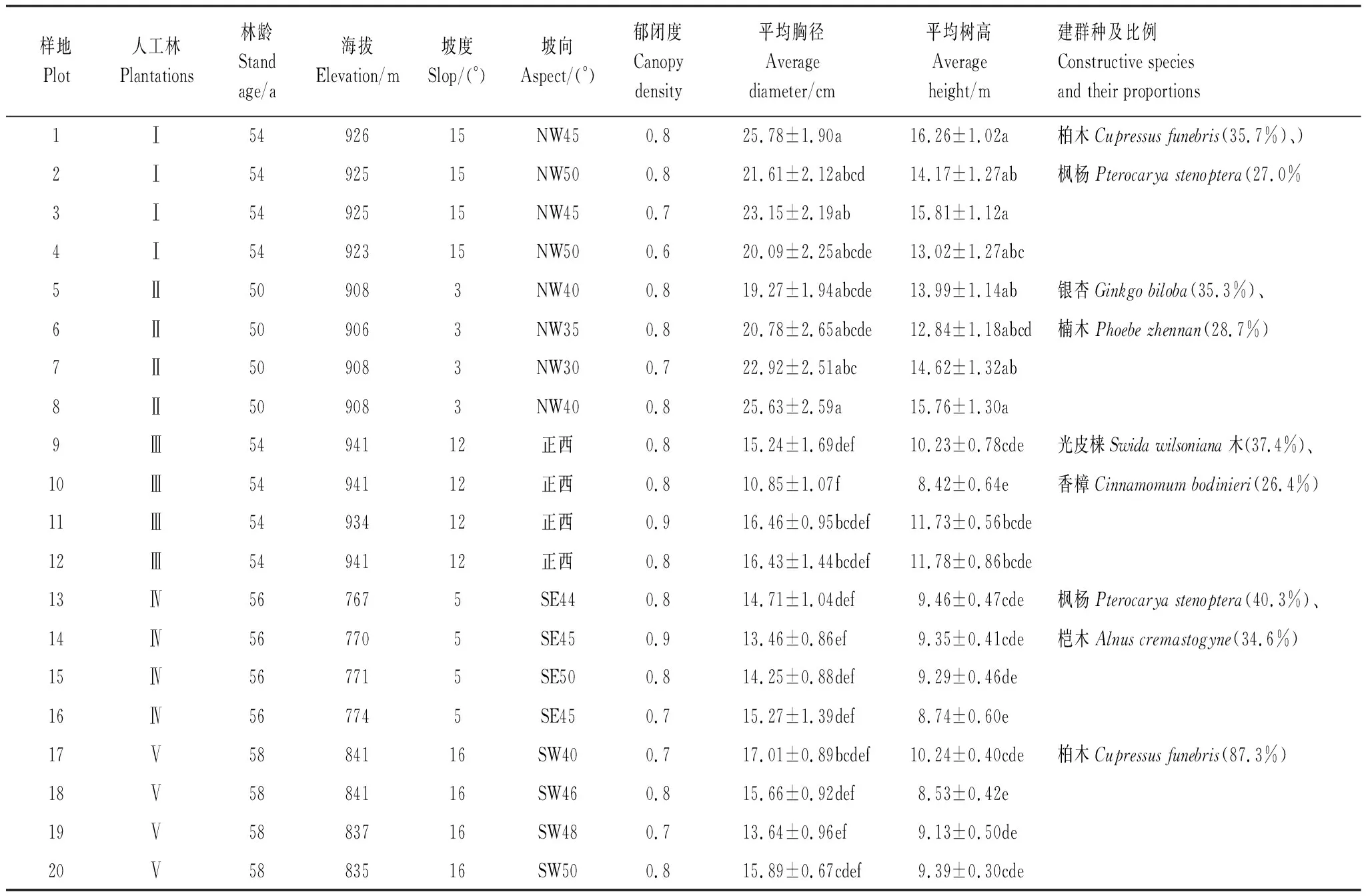
表1 样地概况
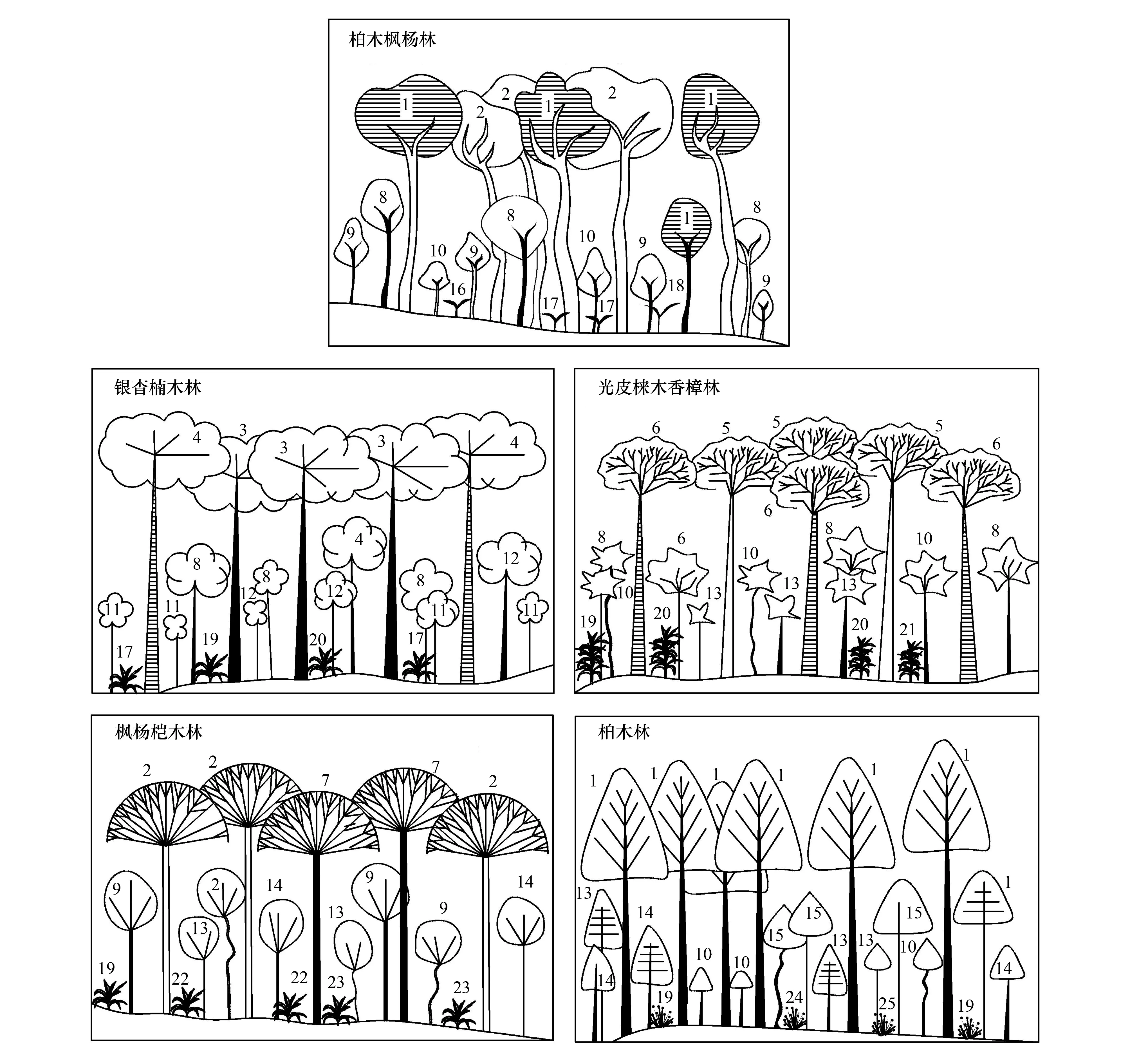
图2 不同人工林群落剖面示意图Fig.2 Illustration of vertical structure of different plantation communities1:柏木Cupressus funebris;2:枫杨Pterocarya stenoptera;3:银杏Ginkgo biloba;4:楠木Phoebe zhennan;5:光皮梾木Swida wilsoniana;6:香樟Celtis sinensis;7:桤木Alnus cremastogyne;8:蚊母树Distylium racemosum;9:箭竹Fargesia spathacea;10:蕊帽忍冬Lonicera pileata;11:五叶地锦Parthenocissus quinquefolia;12:棕榈Trachycarpus fortunei;13:石海椒Reinwardtia indica;14:荚蒾Viburnum dilatatum;15:黄荆Vitex negundo;16:扁竹兰Iris confusa;17:火麻Cannabis sativa;18:细野麻Boehmeria gracilis;19:莩草Setaria chondrachne;20:韩信草Scutellaria indica;21:舞花姜Globba racemosa;22:艾草Leonurus artemisia;23:土牛膝Achyranthes aspera;24:白接骨Asystasiella neesiana;25:三花莸Caryopteris terniflora
采用CANOCO 5.0中的冗余分析(Redundancy analysis,RDA)研究不同人工林灌木层、草本层物种多样性指数与土壤环境因子的关系,并通过蒙特卡洛置换检验确定影响灌草层物种多样性指数的主要因子。其中罗盘仪测量的0―360°坡向方位角由下列公式转换为0―1之间的坡向数值,数值越大表示越干热[11]:
TRASP=1-cos[(π/180)(aspect-30)]/2
2 结果与分析
2.1 不同人工林林下植物组成及重要值
5种不同人工林林下主要植物组成及重要值见表2。灌木层中,柏木枫杨林、银杏楠木林、光皮梾木香樟林分别记录灌木植物35、25、42种,蚊母树在这3种人工林中的重要值均为最大,分别为0.1302、0.3402、0.1427;枫杨桤木林与柏木林分别记录灌木植物34、35种,石海椒、黄荆(Vitexnegundo)分别是这2种人工林重要值最大的树种,分别为0.2859、0.3596。草本层中,柏木枫杨林共记录草本植物31种,扁竹兰重要值最大,为0.2861;银杏楠木林、光皮梾木香樟林、枫杨桤木林与柏木林分别记录草本植物27、45、41、41种,莩草在这4种森林类型中长势均较好,其重要值介于0.0844—0.2581之间。
2.2 不同人工林林下植物多样性
由表3可见,5种人工林灌木层的多样性指数H、优势度指数H′、丰富度指数D具有显著差异(P<0.05),且具有相似的变化趋势,表现为光皮梾木香樟林略大于柏木枫杨林,二者的H、H′、D显著大于其余3种人工林;不同人工林灌木层的均匀度指数Jsw无显著差异(P>0.05)。草本层的H、D、Jsw差异显著,H、Jsw均表现为柏木林>枫杨桤木林>光皮梾木香樟林>柏木枫杨林>银杏楠木林,D表现为柏木林>光皮梾木香樟林>枫杨桤木林>柏木枫杨林>银杏楠木林;不同人工林草本层H′无显著差异。
2.3 不同人工林土壤理化性质
由表4可见,不同人工林的土壤pH值、有机质、全氮、全磷、速效钾、速效磷具有显著差异(P<0.05),土壤含水量、土壤容重、全钾差异不显著(P>0.05)。土壤pH值均大于7,呈弱碱性,枫杨桤木林pH值显著低于其他4种人工林类型。有机质、全氮变化趋势相似,表现为银杏楠木林最高,柏木枫杨林与枫杨桤木林稍低于银杏楠木林,光皮梾木香樟林与柏木林最低。全磷表现为柏木枫杨林>枫杨桤木林>银杏楠木林>柏木林>光皮梾木香樟林。速效磷与速效钾变化规律一致,表现为银杏楠木林显著高于其余4种人工林,且均呈银杏楠木林>枫杨桤木林>柏木枫杨林>光皮梾木香樟林>柏木林规律分布。

表2 不同人工林林下主要植物组成及重要值
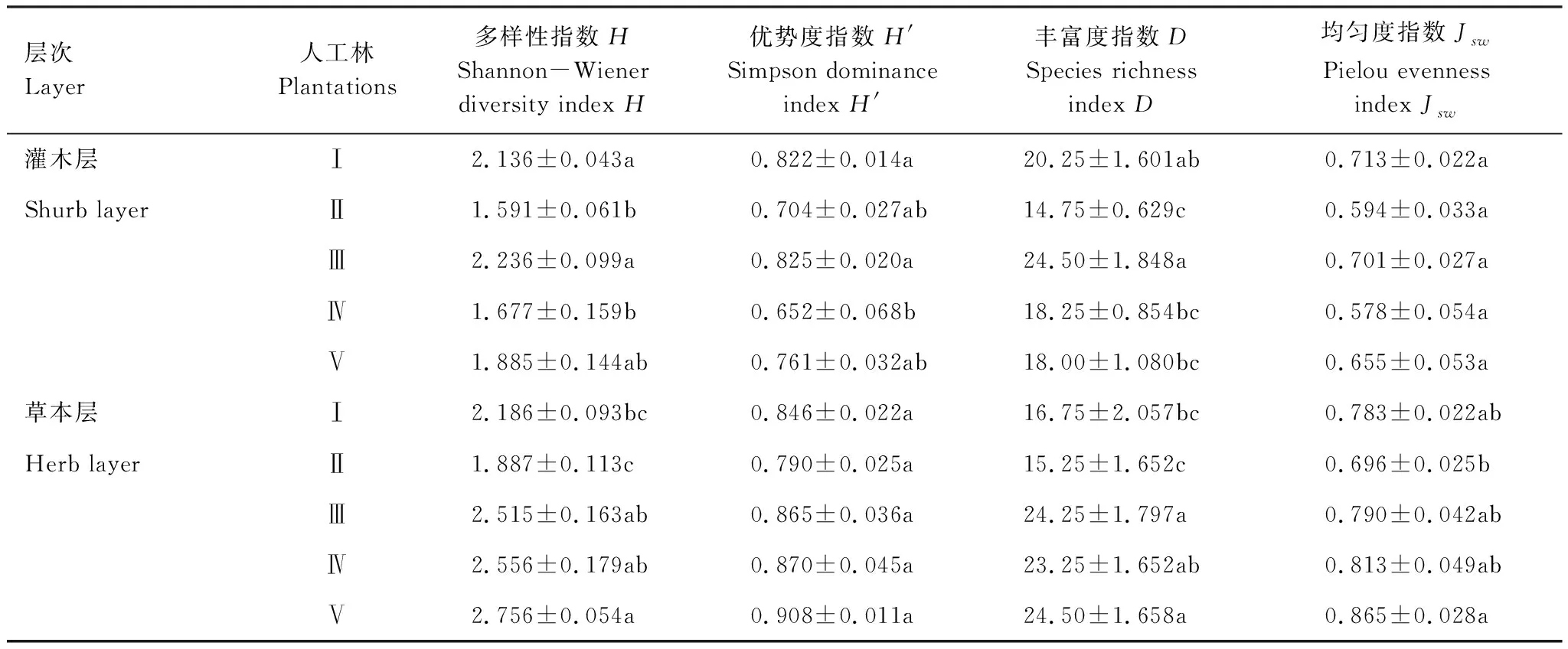
表3 不同人工林灌木层与草本层物种多样性指数比较
2.4 物种多样性指数与土壤环境因子的RDA分析
由表5及图3可知,灌木层中,RDA1轴主要反映有机质(Organic matter,OM)、速效钾(Available potassium,AK)、速效磷(Available phosphorus,AP)、坡度(Slop)的变化,在RDA1轴上,随着OM、AK、AP减少及Slop增加,多样性指数H、优势度指数H′、丰富度指数D、均匀度指数Jsw均增加。RDA2轴主要反映土壤含水量(Soil water content,SWS)的变化,在RDA2轴上,随着SWS减少,D减小,而H、H′、Jsw均增加。蒙特卡洛置换检验结果表明,OM、AP是显著影响灌木层物种多样性的土壤因子。
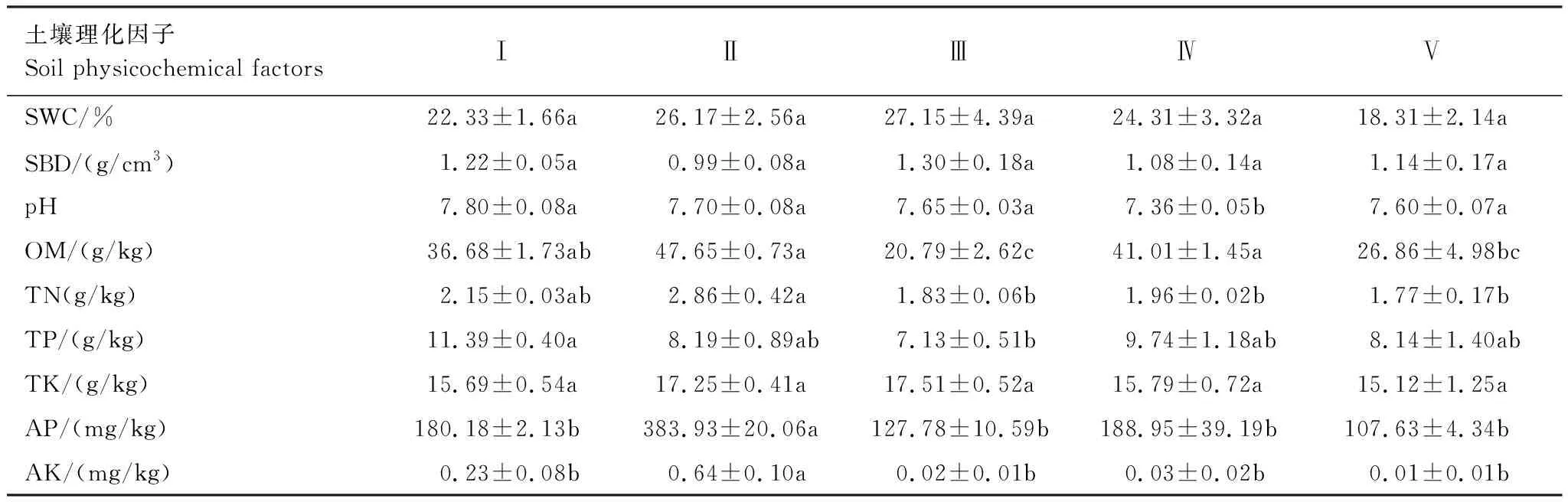
表4 不同人工林土壤理化性质
由表5及图4可知,草本层中,RDA1轴主要反映AP、pH、AK、全氮(Total nitrogen,TN)、坡向(Aspect)的变化,在RDA1轴上,随着AP、pH、AK、TN增加及Aspect减小,H、H′、D、Jsw均减小。RDA2轴主要反映全钾(Total potassium,TK)的变化,在RDA2轴上,随着TK减少,D减小,而H、H′、Jsw均增加。AP、pH、TP是显著影响草本层物种多样性的土壤因子。
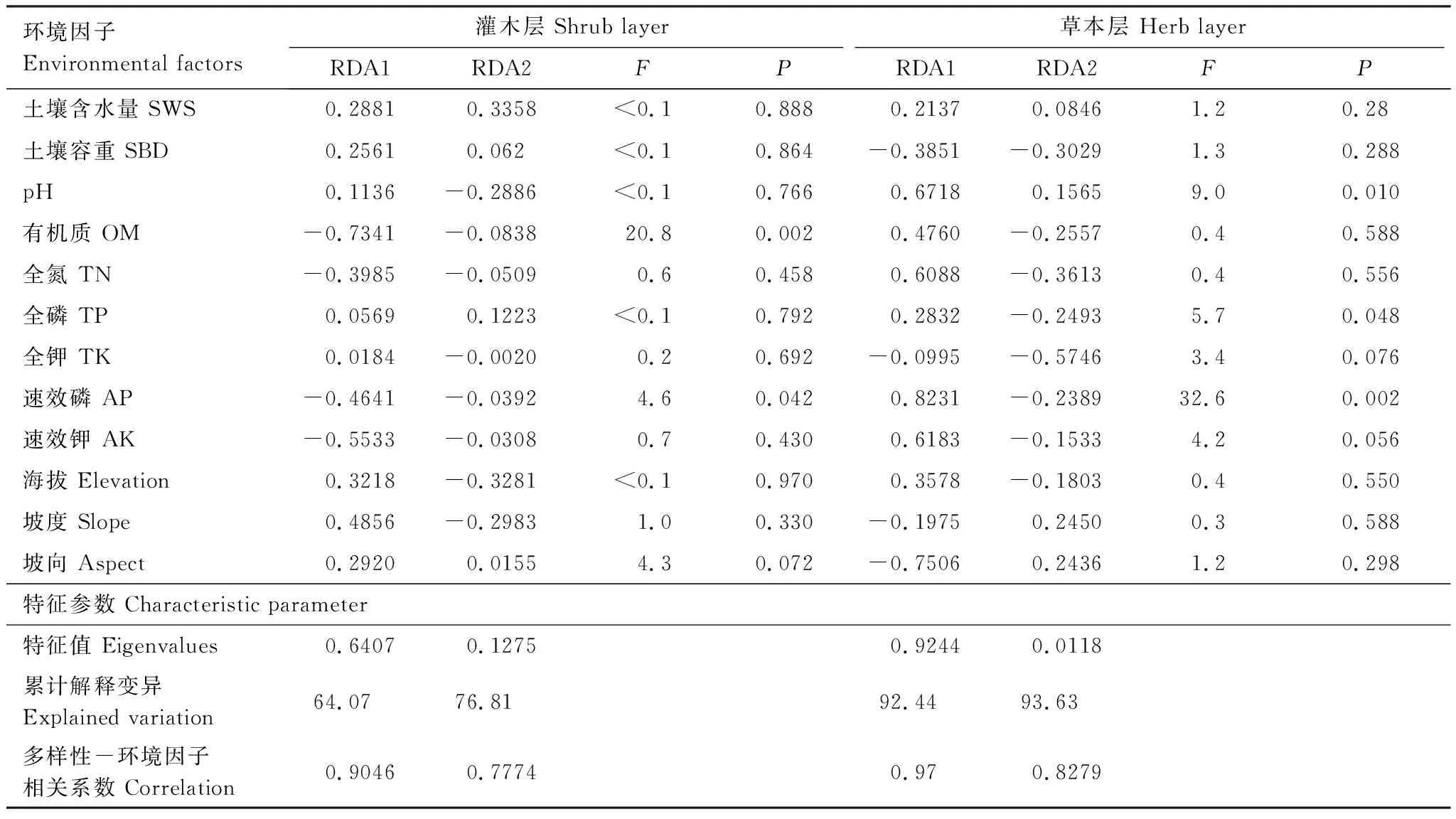
表5 冗余分析(RDA)排序及蒙特卡洛置换检验结果

图3 灌木层冗余分析RDA排序图Fig.3 Redundancy analysis (RDA) biplot of shrub layer S-H:灌木层多样性指数Shannon-Wiener diversity index of shrub layer;S-H′:灌木层优势度指数Simpson dominance index of shrub layer;S-D:灌木层丰富度指数Species richness index of shrub layer;S-JSW:灌木层均匀度指数Pielou evenness index of shrub layer;SWS:土壤含水量Soil water content;SBD:土壤容重Soil bulk density;pH:土壤pHSoil pH;OM:有机质Organic matter;TN:全氮Total nitrogen;TP:全磷Total phosphorus;TK:全钾Total potassium;AP:速效磷Available phosphorus;AK:速效钾Available potassium

图4 草本层冗余分析RDA排序图Fig.4 Redundancy analysis (RDA) biplot of herb layer H-H:草本层多样性指数,Shannon-Wiener diversity index of herb layer;H-H′:草本层优势度指数,Simpson dominance index of herb layer;H-D:草本层丰富度指数,Species richness index of herb layer;H-JSW:草本层均匀度指数,Pielou evenness index of herb layer
3 讨论
3.1 不同人工林林下植物组成及物种多样性
林下植物组成是群落的基本特征[9-10,19]。在本研究中,不同人工林灌草层植物组成及数量分布特征不同,表明研究区林下植物组成易受上层林冠及林内微环境的影响[11],与Rawlik等[5]、赵耀等[15]的研究结果相似。Jang等[6]对森林木本植物生物量采取不同强度收获后,经38年灌木植物生物量均超过处理前水平。张柳桦等[3]认为人工林中灌木层最具更新活力,林下物种多样性水平与更新潜力主要取决于灌木层。本研究中灌木层多样性指数H、优势度指数H′、丰富度指数D均以光皮梾木香樟林最大,草本层植物组成也较为丰富,因此认为其林下整体多样性水平最高,利于林下植物多样性维持及发展。光照影响着林下植物覆盖方式和个体物种丰度,被认为是林下植物生长最重要的驱动力之一[11]。Tinya等[21]发现林下维管和苔藓植物的多样性和物种组成与不均匀光照条件有关。本研究中草本层的H、D、均匀度指数Jsw均以柏木林最大,与王媚臻[9]认为的柏木林多为“低产低效林分”不同,这可能是由于柏木为针叶树种,其林下光照条件较好,为种子库中非耐阴性草本先锋物种种子的入侵和萌发创造了有利条件[15]。银杏楠木林灌木层H、D与草本层H、D、Jsw均最小,其林下整体多样性水平最低,物种组成最单一,不利于林下植被可持续发展。
3.2 不同人工林土壤理化性质
土壤结构与肥力水平是植物多样性与稳定性的基础[1],其决定着生态系统结构、功能及生产水平[10]。在本研究中,5种人工林土壤含水量、土壤容重、全钾均无显著差异,说明林分差异对土壤物理结构未产生明显影响。目前森林生态系统土壤肥力与植物多样性关系的研究结果主要有两种:一是两者呈正相关。如Zheng等[22]认为植物多样性越高的森林群落其生产力也越高,土壤中植物凋落物和根系也越多,因此土壤养分含量也相对更高;Chen等[23]发现森林、草地、农田生态系统的土壤有机碳、微生物量碳含量随群落物种丰富度增加均呈上升趋势。二是两者为负相关。其理论解释主要包括物种数量特征与资源生态位维度两种角度,如 Strubeltet等[24]认为土壤养分含量高使植物种间竞争加剧,优势种利用有利条件迅速生长会导致其他物种生存率减小;Yang等[25]认为植物为同一类土壤养分资源竞争,限制养分资源的增加降低了地下资源权衡维度,使共存物种数量减少。与上述后种结果一致,本研究中有土壤机质、全氮、速效钾、速效磷均以银杏楠木林最高,而光皮梾木香樟林与柏木林最低,与其林下植物多样性水平相反。桤木为非豆科共生结瘤固氮植物[26],枫杨桤木林土壤pH值显著低于其他人工林,这可能与桤木根瘤的酸性有机胺释放使土壤游离氢离子富集有关。土壤磷来源相对固定,且移动性差,因此基本不能被植物直接吸收利用[14]。本研究中全磷呈现柏木枫杨林>枫杨桤木林>银杏楠木林>柏木林>光皮梾木香樟林规律,柏木枫杨林与枫杨桤木林全磷最高,这可能与枫杨对土壤养分的特异性吸收有关[9]。
3.3 林下植物多样性与土壤环境因子的关系
植物多样性与土壤因子之间关系密切[9]。如姚俊宇等[1]发现速效钾、有机质、有效磷对山矾次生林植物多样性影响较大。Fu等[2]表示有机质、全氮对中国东岭山落叶阔叶林植物多样性影响较大。王媚臻等[9]发现全氮、钾、有机质为影响柏木人工林林下植物多样性的主要因子。Xu等[18]发现pH、速效磷是热带季雨林植物多样性的主导因子。在本研究中,有机质、速效磷为影响灌木层多样性的主要因子,pH、全磷、速效磷为影响草本层多样性的主要因子,进一步强调了pH、有机质、土壤磷对森林群落植物多样性的主导驱动作用。本研究未发现土壤含水量、容重对林下灌草层多样性具有显著影响,这可能与土壤物理性质在不同人工林下差异不显著有关。
海拔被认为是影响植物多样性格局的决定性因素[4,11],其包含了温度、湿度、土壤等多种环境因子变化而在空间尺度上形成的生态尺度[4]。坡度主要影响土壤含水量及养分再分配,坡度大会使上坡位土壤养分更易流失[17]。坡向则通过调节太阳辐射及降水再分配,从而影响土壤含水量、土壤温度、土壤风化作用[19]。在本研究中,海拔、坡度、坡向均不是影响林下灌草层多样性的主导因子,这可能是由于海拔变幅较小(767—926 m)、坡度范围较缓(3—16°),因地形因子而导致的林下微环境差异不明显,与尤业明等[11]、赵耀等[15]的研究结果相似。另外,地形因子作用被林冠特征、土壤因子、气候微变化等因素掩盖也可能是其对林下植物多样性无明显影响的原因[11]。
4 结论
综上所述,5种人工林林下植物组成、物种多样性、土壤理化性质具有差异,光皮梾木香樟林林下植物组成最丰富、物种多样性水平最高,经营此种林分有利于林下植物多样性的可持续发展。同时发现,土壤pH、有机质、土壤磷为研究区林下植物多样性的主要土壤驱动因子。本文主要分析了不同人工林类型对林下植物多样性及土壤理化性质的影响,进一步可继续研究树种不同混交比例对林下各层次多样性及土壤性质的影响。
猜你喜欢
现代农业科技(2022年10期)2022-12-17
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
文萃报·周五版(2020年17期)2020-06-01
北方音乐(2019年6期)2019-07-16
家教世界·创新阅读(2019年4期)2019-05-04
家教世界(2019年10期)2019-04-26
作文成功之路·作文交响乐(2019年10期)2019-01-10
安徽文学(2018年11期)2018-11-19
扬子江(2018年1期)2018-01-26
家教世界·创新阅读(2017年11期)2017-11-27
