
不同种源樟树叶片形态特征及生长差异分析
2021-03-30 05:45肖祖飞张北红张琴马一丹金志农张海燕李凤吕雄伟
广西植物 2021年2期
肖祖飞 张北红 张琴 马一丹 金志农 张海燕 李凤 吕雄伟
摘要:為了解不同种源樟树叶片形态特征和生长差异,该文以30个种源樟树为研究对象,对其叶长、叶宽、叶柄长、周长、叶面积、长宽比、形态因子、株高、地径等指标进行测定和差异性分析。结果表明:(1)30个种源间叶片性状的变异系数为3.88%~16.14%,显示不同种源樟树叶片形态特征存在显著差异;叶长、叶宽、叶柄长、周长、面积、叶厚度重复力超过75%,叶长宽比和形态因子重复力超过60%,显示樟树叶片形态差异在一定程度上受遗传控制;种源内个体间叶片形态特征存在差异,叶长、叶宽、叶面积、叶柄长和长宽比变异系数较大,叶周长、叶厚度和形态因子变异系数较小。(2)主成分分析结果显示,选出的3个主成分方差累积贡献率达到86.18%,叶长、周长、面积、叶宽和叶厚度是这3个主成分的主导因子,这些参数对叶片形态划分起主要作用。(3)不同种源间的株高、地径存在极显著差异,来自中国湖南的种源生长表现最好,其次为中国福建、江苏以及日本大阪,中国江西本地种源不存在优势,生长表现较差;种源内个体间株高和地径存在一定差异,中国湖北黄冈种源株高和地径变异系数较小,个体间均匀度较好,而中国湖北宜昌和江西安福种源变异系数较大,个体间差异较大,为优株的选择创造了条件。樟树种源间和种源内存在丰富的遗传变异,为樟树的优质种源以及良种的选育和改良提供了基本材料。
关键词: 樟树, 叶片形态, 种源, 生长差异
中图分类号:Q945
文献标识码:A
文章编号:10003142(2021)02023310
Abstract:In order to understand the leaf morphological characteristics and growth differences of Cinnamomum camphora from different provenances, we measured and analyzed the leaf morphological characteristics and growth indexes of C. camphora from 30 provenances, including leaf length, width, petiole lenghth, perimeter, leaf area, length to width ratio, morphous factor, plant height and ground diameter. The results were as follows: (1) The coefficient of variance of leaf morphological characteristics were 3.88%-16.14% among the 30 provenances, which indicated that the most significant difference among provenances. There existed high heritability in leaf length, leaf width, leaf petiole lenghth, leaf thickness, leaf area, leaf perimeter,reached more than 75%,and the leaf length to width ratio and morphous factor reached more than 60%.There were significant differences in leaf morphological characteristics and growth of C. camphora from different provenances, and the difference of C. camphora leaf traits was controlled by genetic control to a certain degree. The leaf morphological characteristics were different in all of the individuals within provenances. The variation coefficients of leaf length, width, area, petiole lenghth and length to width ratio were larger, while the variation coefficients of perimeter, leaf thickness and morphological factors were smaller. (2) Principal component analysis was performed to select three principal components and their cumulative contributions of variance accouted for 86.18%. Leaf length, area, perimeter, width and thickness were dominating elements, and these parameters played an important role in the division of leaf morphology. (3) There were significant differences in plant height and ground diameter among provenances. The provenances from Hunan of China had the best growth performance, followed by Fujian, Jiangsu, China and Osaka, Japan. The native provenances from Jiangxi, China was no advantage, and the growth performance was poor. The plant height and ground diameter were differences in all of the individuals within provenances. The variation coefficient of plant height and ground diameter from Huanggang of Hubei, China provenances was smaller and the uniformity among individuals was better. The variation coefficient from Yichang of Hubei and Anfu of Jiangxi, China provenances was larger and the difference among individuals was larger, which creates conditions for the selection of superior plants. It is suggested that there are rich genetic variations in C. camphora provenances and within provenances, which provide basic materials for the breeding and improvement of C. camphora with high quality provenances and improved varieties.
Key words: Cinnamomum camphora, leaf morphological, provenances, growth differences
植物表观性状的差异是植物自身遗传因素和环境因子共同作用,经过长期的适应,群体和个体间均产生不同程度和形式的变异,以适应不同的环境(卢欣石和何琪,1999)。植物会通过对自身结构及生理特性的调节来响应环境的变化,进而适应环境的变化(马姜明等,2012) 。植物叶片的形态特征,如叶片形状和大小、叶柄长度等,是植物长期适应环境的结果,与植物的生长发育密切相关(Xu et al., 2009)。植物叶片是植物最重要的同化器官,叶片的形态与植物的营养、生长、生理、生态因子等密切相关。
樟树(Cinnamomum camphora)又名香樟,樟科樟属,常绿乔木,高大挺拔,原产我国东南及西南各地,尤以江西、福建、台湾等省为最多(陈有民,2006)。樟树是我国重要的经济树种,根据叶片精油主成分不同分为芳樟、脑樟、桉樟、异樟、龙脑樟等化学类型,尤其以富含芳樟醇的樟树具有极重要经济价值。对樟树的研究多见于良种的选育(王长宪,2006;胡文杰,2013;王建军,2015)、栽培与组培快繁(郑威等,2013;黎祖尧等2013;叶润燕等,2016;肖祖飞等,2017)、材性(薛晓明等,2016)、抗性(王利宝等,2010;贺珑等,2016)等方面。长期的生殖隔离和各种环境因子的综合作用,使樟树在遗传上存在着较大的地理变异,不同个体和群体间的形态特征和生长差异显著。有研究表明,不同生境和地理种源樟树种子形态和结构存在显著差异(张雷等,2017),不同种源樟树苗高、地径和分枝数存在显著差异(姚小华等,1999;姚小华,2000)。樟树叶片是樟树的重要营养器官,是影响樟树生产力大小的重要器官之一。因此,樟树叶片的形态特征直接影响樟树的生长发育。目前,关于不同种源樟树叶片形态特征差异未见报道。本文以中国江西和福建、日本大阪等地区30份樟树种质资源为材料,对其叶片形态特征和生长指标进行测定,分析不同种源间和种源内樟树叶片形态差异性和生长差异性,旨在为进一步开展樟树种质资源分子水平的遗传多样性和相关性研究提供理论指导,同时为樟树的育种工作提供基础数据。
1研究地概况
试验于2016年11月在南昌工程学院生物技术试验基地樟树种质资源圃进行。该基地位于江西省南昌市东部,115°27′—116°35′E、28°09′—29°11′N,平均海拔为16 m,属典型亚热带季风气候,湿润温和,夏季多偏南风,冬季多偏北风,年平均气温为17~17.7 ℃,降雨量和日照充足,年降雨量1 600~1 700 mm,年日照时间1 723~1 820 h,无霜期长、冰冻期短。土壤为微酸性红壤。
2材料与方法
2.1 材料
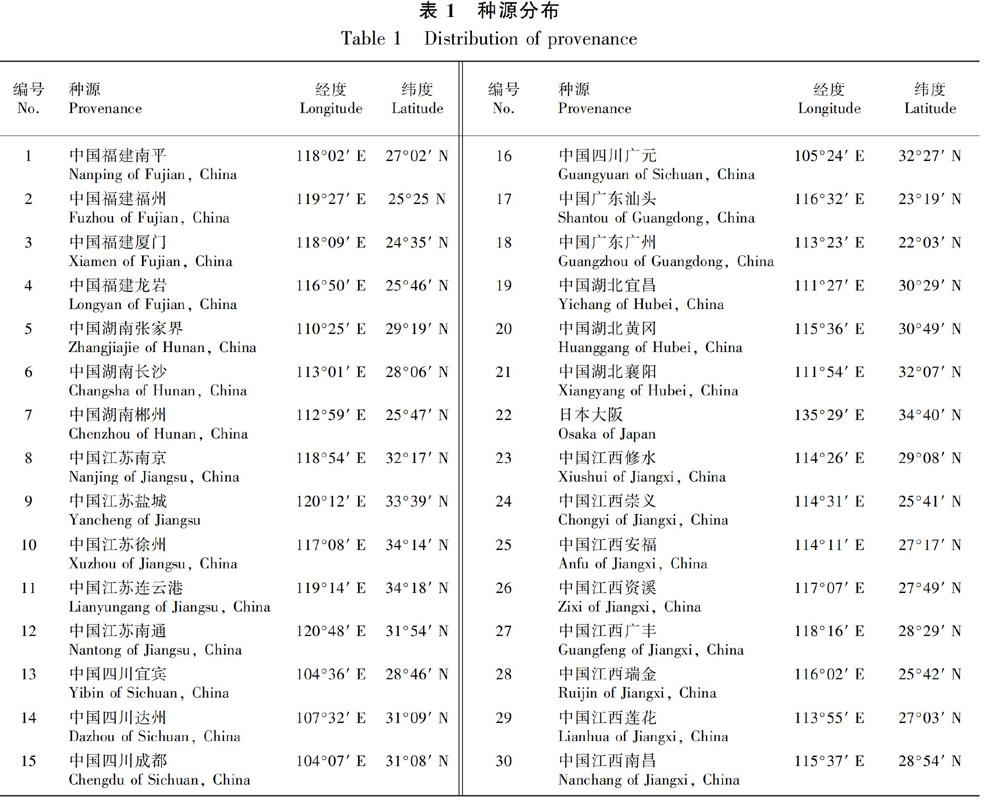
试验材料为3年生樟树实生苗,来自樟树主要分布区,分别为中国江西8个种源地、中国福建4个种源地、中国湖南3个种源地、中国湖北3个种源地、中国广东2个种源地、中国四川4个种源地、中国江苏5个种源地和日本大阪1个种源地,共30个种源地(表1)。种苗立地条件基本一致,苗木管理方式一致。
2.2 方法
2016年11月份,在樟树种质资源圃内每个种源随机选取30个单株,每株四面各选取1个生长健壮的枝条,从枝条顶部数起摘取第5片或第6片叶子,3次重复,采用Yaxin1241叶面积仪测定叶面积、叶长、叶宽、周长,计算叶片长宽比(平均叶片长度/平均叶片宽度)、叶片形态因子(4π×叶面积/周长2),用游标卡尺测量叶柄的长度和叶片厚度。同时,用卷尺测定植株的高度和游标卡尺测定离地5 cm处植株的地径。
2.3 数据处理
采用Excel2007软件进行数据的录入、整理和方差分析,利用SPSS19软件对各形态指标进行统计以及相关性分析、主成分分析。
3结果与分析
3.1 不同种源间和种源内樟树叶片形态特征
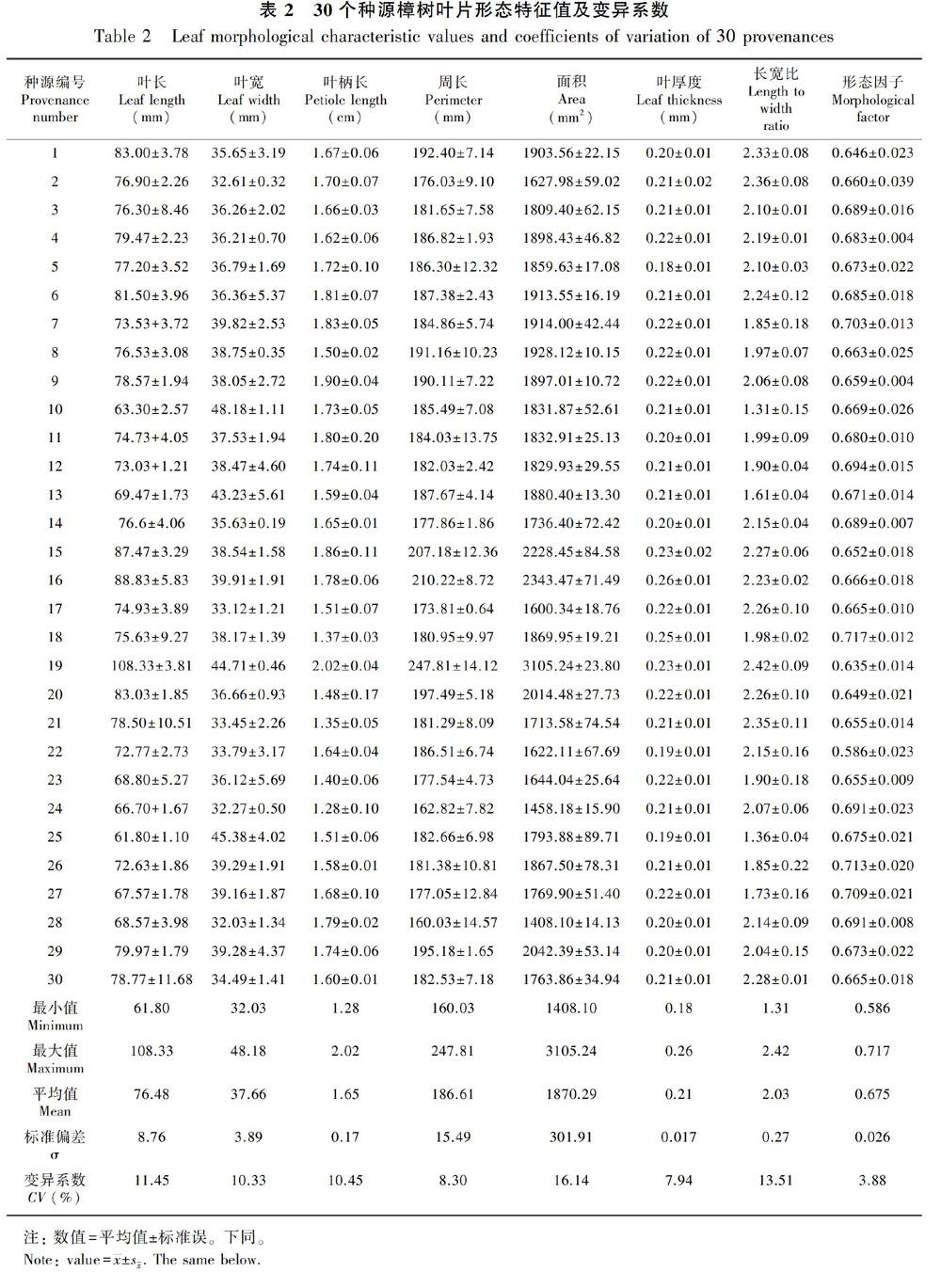
3.1.1 不同种源间樟树叶片形态差异由表2可知,叶长变异幅度为61.80~108.33 mm,叶长最长的是中国湖北宜昌,最短的是中国江西安福;叶宽变异幅度为32.03~48.18 mm,叶宽最宽的是中国江苏徐州,最窄的是中国江西瑞金;叶柄长变异幅度为1.28~2.02 cm,叶柄最长的是中国湖北宜昌,最短的是中国江西崇义;周长变异幅度为160.03~247.81 mm,周长最长的是中国湖北宜昌,最短的是中国江西瑞金;面积变异幅度为1408.10~3105.24 mm2,面积最大的是中国湖北宜昌,最小的是中国江西瑞金;叶厚度变异幅度为0.180~0.262 mm,叶厚度最大的是中国四川广元,最小的是中国湖南张家界;叶长宽比变异幅度为1.48~2.03,长宽比最大的是中国湖北宜昌,最小的是中国江苏徐州。
总体上看,不同种源樟树叶片形态存在较大变异,叶形变异最大的是来源于中国湖北省,而来自中国江西省和日本大阪的叶形变异普遍偏小,低于平均值。变异系数为3.88%~16.14%,形态因子(3.88%)﹤叶厚度(7.94%)﹤周长(8.30%)﹤叶宽(10.33%)﹤叶柄长(10.45%)﹤叶长(11.45%)﹤长宽比(13.51%)﹤面积(16.14%)。形态因子和叶厚度变异系数较小,标准偏差为0.026、0.017,表明樟树叶片的形态因子、叶厚度是比较稳定的性状,而长宽比和叶面积变异系数较大,尤其叶面积变异系数大于15%,表明不同种源间这两项指标差异较大。
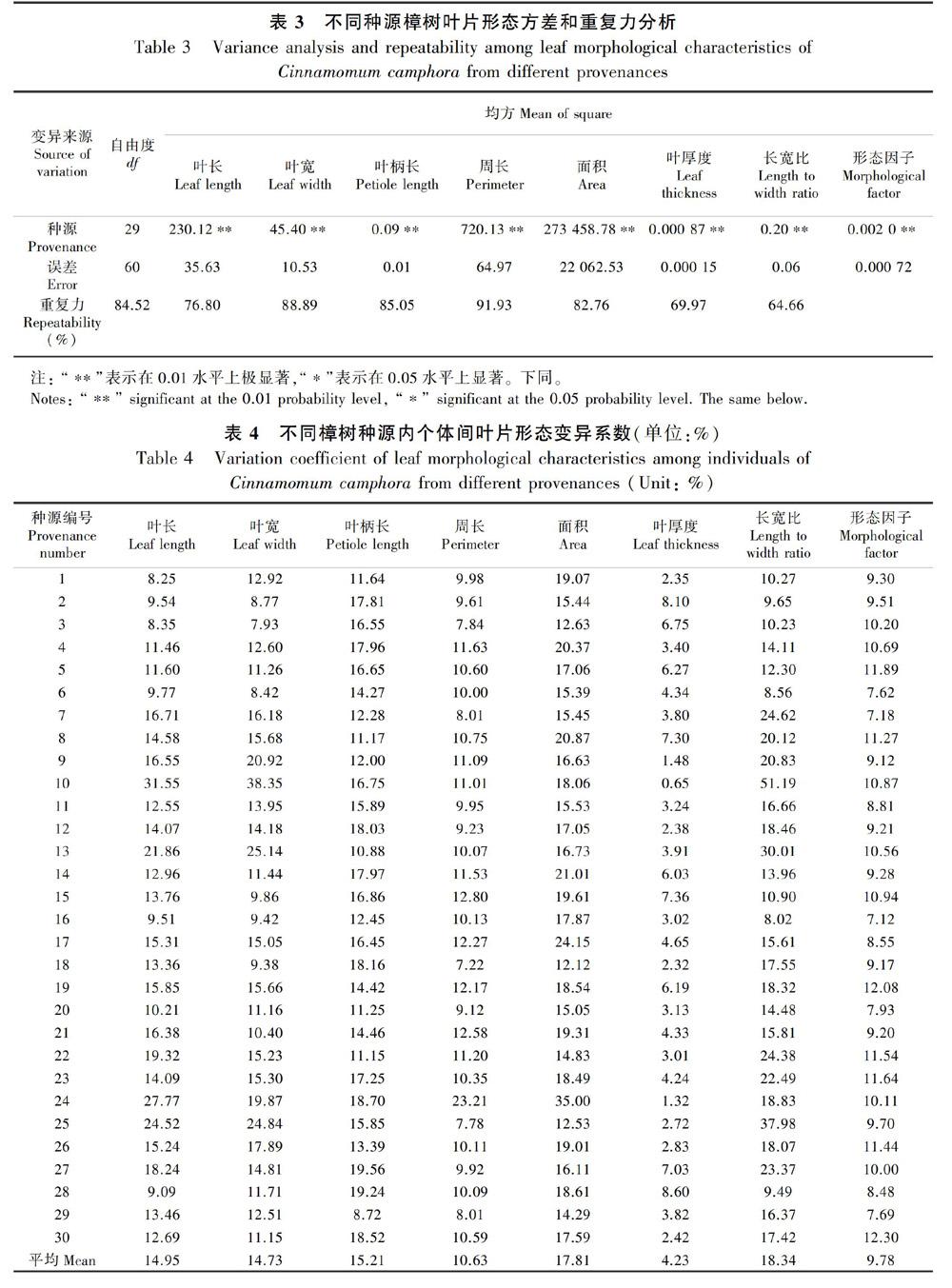
方差分析结果表明(表3),不同种源樟树叶长、叶宽、叶柄长、周长、面积、叶厚度、长宽比及形态因子均存在极显著差异; 叶长、叶宽、叶柄长、周长、面积、叶厚度重复力大于75%,葉长宽比和形态因子重复力也大于60%,表明樟树叶片形态差异在一定程度上是受遗传控制的。
3.1.2 种源内樟树叶片形态差异由表4可知,种源内个体间叶长差异最大的是来自中国江苏徐州种源(31.55%),差异最小的是来自中国福建福州种源(8.25%),二者之间相差23.3%;叶宽差异最大的是来自中国江苏徐州种源(38.35%),差异最小的是来自中国福建厦门种源(7.93%),二者之间相差30.42%;叶柄长差异最大的是来自中国江西广丰种源(19.56%),差异最小的是来自中国江西莲花种源(8.72%),二者之间相差10.84%;叶周长和叶面积差异最大的均是来自中国江西崇义种源(23.21%,35.00%),差异最小的是来自中国广东广州种源(7.22%,12.12%),二者之间相差15.99%、22.88%;叶厚度差异最大的是来自中国江西瑞金种源(8.60%),差异最小的是来自中国江苏徐州种源(0.65%),二者之间相差7.95%;葉长宽比差异最大的是来自中国江西安福种源(37.98%),差异最小的是自四川广元种源(8.02%),二者之间相差29.96%;形态因子差异最大的是来自中国江西南昌种源(12.30%),差异最小的是来自中国四川广元种源(7.12%),二者之间相差5.18%。总体上,种源内个体间叶长、叶宽和长宽比变异系数较大,变异系数较小的是叶厚度和形态因子。
3.2 樟树叶片形态性状与纬度、经度相关性
由表5可知,仅叶面积与纬度呈负相关,形态因子与经度呈负相关,叶长、叶宽、叶柄长、叶周长、叶厚度和长宽比与纬度、经度关联性不大。
3.3 不同种源地樟树叶片形态性状差异主成分分析
30个种源樟树叶片形态性状差异主成分分析结果表明(表6),不同种源樟树叶片形态性状变异的主成分y1、y2和y3的贡献率分别为46.576%、24.628%和14.976%;从y1看,叶长特征值为0.925、周长特征值为0.970、面积特征值为0.955,三者特征值较大,对y1有较强的正向负荷,是第一主成分的主导因子,对叶片形态分类起重要作用;从y2看,叶宽特征值达到0.906,对y2有较强的正向负荷,是第二主成分的主导因子,对叶片的形态分类起主导作用;从y3看,叶厚度特征值较大为0.822,是第三主成分的主导因子。前2个主成分方差累积贡献率为71.204%,前3个主成分方差累积贡献率为86.180%,累计贡献率增长较慢,表明樟树叶片形态特征存在较大的遗传多向性。
3.4 不同种源地樟树株高和地径比较
3.4.1 不同种源地樟树株高和地径差异性分析由表7可知,株高的变异幅度为57.48~200.09 cm。对比平均株高,来自中国四川广元和湖南郴州、湖南长沙的种源生长较快,平均株高分别为178.37、173.92和171.99 cm,远超过总体平均水平(132.45 cm);其次是来自中国江苏、日本大阪和中国福建的种源;来自中国江西、湖北和广东的种源生长较慢,大部分种源株高低于总体平均水平。
地径的变异幅度为0.68~4.99 cm。对比平均地径,来自中国福建福州、福建厦门、福建龙岩和福建南平的种源地径较大,平均地径分别为3.49、3.21、3.16和2.89cm,超过总体平均水平(2.35 cm);其次是来自中国湖南、江苏和日本大阪的种源;来自中国江西、湖北和广东的种源地径较小。
总体上来看,来自中国湖南的种源生长表现最好,其次为中国福建、江苏和日本大阪,中国江西本地种源生长表现较差,没有生长优势。
3.4.2 樟树种源内个体间株高和地径变异系数由表8可知,种源内株高变异系数较大的是来自中国湖北宜昌(15.85%)和江西安福(15.58%)种源,其次是来自中国广东广州(10.05%)和江西崇义(10.09%)种源,最小的是来自中国湖北黄冈(1.49%)种源;地径变异系数最大的是来自中国江西安福(32.02%)种源,其次是中国四川达州(26.73%)和四川宜昌(24.26%)种源,较小的是中国江苏盐城(5.27%)和湖北黄冈(6.11%)种源。在30个樟树种源中,来自中国湖北黄冈种源株高和地径变异系数较小,个体间均匀度较好;而中国湖北宜昌和中国江西安福种源变异系数较大,个体间差异较大,为优株的选择创造了条件,在这两个种源进行个体选择更具有意义。
4讨论与结论
植物个体的差异是由本身遗传因素和环境因子决定的。叶片是植物对环境变化最为敏感的器官之一,受温度、光照、水分等环境条件的影响,植物叶片的长度、宽度、叶柄长度、叶片厚度、叶周长、叶面积等形态特征会产生变化(郭玉华等,2004)。有研究表明,不同种源元宝枫叶片的形态差异较大,与地理、环境因素相关,叶长随纬度的增加显著变短,随年均温和年降水量的升高显著变长;叶片面积随温度和降水量的增加而变大,反之,叶片就小;叶柄长度随纬度和海拔的升高而变短(任红剑等,2017)。通过分析我国7省区天女木兰种群叶片形态特征,发现天女木兰各种群叶片形态存在显著差异, 聚类分析表明地理位置远近与亲缘关系密切相关,分布地域越接近,其亲缘关系越密切(张淑媛等,2014)。苦楝叶片生长性状差异在一定程度上是受遗传控制,叶片性状与纬度、海拔、年均温、年降水量、无霜期、年均日照均相关(陈丽君等,2016)。本研究对30个种源地樟树叶片的8个形态性状参数进行数学分析,其变异系数为3.88%~16.14%,显示不同种源樟树叶片形态特征存在显著差异,种源内个体间也存在一定差异,这主要是由于遗传和环境因子共同作用的结果,30个樟树种源来自樟树主要自然分布区域,分布广泛,地形、气候、土壤等环境条件差异较大,而且长期的地理隔离和自然选择,使得樟树种源间和种源内表观形态特征产生丰富的变异。叶长、叶宽、叶柄长、周长、面积、叶厚度重复力大于75%,叶长宽比和形态因子重复力也大于60%,显示樟树叶片形态差异在一定程度上是受遗传控制的,能够稳定遗传给后代,樟树优良种源的选择潜力较大。主成分分析选出3个主成分,方差累积贡献率达到86.18%,叶长、周长、面积、叶宽和叶厚度是这3个主成分的主导因子,对叶片形态划分起主要作用。叶片各形态性状大多数与纬度、经度关联性不大,究其原因可能与遗传漂变、引种、环境因子等相关,真正的原因有待进一步研究确认。
树高和地径作为幼林生长的重要指标,对优良种源的早期选择具有重要意义。有研究表明,樟树种源和家系间苗高和地径生长量存在极显著差异(黄云鹏,2008;蔡燕灵等,2013;陈辉祥,2015),樟树苗期生物量种源间差异显著,苗高、地径、地上部分等性状之间存在显著的表型和遗传相关,地径与苗期生物量紧密相关(孙祥银等,1999)。本研究结果表明,不同种源间在生长性状上存在差异,株高、地径在种源间存在极显著差异,来自中国湖南的种源生长表现最好,其次为中国福建、江苏和日本大阪,中国江西本地种源生长表现较差,本地种源不存在优势,这种差异很可能是树木本身的基因差异和生长环境互作所致。30个樟树供试种源,来自不同地理区域和生长环境,经过长期的自然选择和地理隔离,不同种源间樟树基因型产生一定差异。因此,不同种源樟树在相同环境条件下生長,株高和地径差异显著,甚至外来种源生长优于本地种源。
表观变异是由遗传因子和环境因素共同作用的结果,表型变异必然蕴藏遗传变异,一般来说表型变异越大,遗传变异也就越大。本研究不同种源樟树叶片形态性状、株高、地径等表观性状具有丰富的自然变异,表明樟树的遗传改良具有广阔前景。
参考文献:
CAI YL, ZENG LH, LIAN HM, et al., 2013. Early selection of local provenance of Cinnamomum camphora in Guangdong Province[J]. Guangdong For Sci Technol, 29(3): 7-12.[蔡燕灵, 曾令海, 连辉明, 等, 2013. 樟树局部种源早期选择研究[J]. 广东林业科技, 29(3): 7-12.]
CHEN HX, 2015. Variation of growth traits of Cinnamomum camphora with different provenances[J]. Prot For Sci Technol, (4): 7-9. [陈辉祥, 2015. 樟树不同地理种源生长性状差异性研究[J]. 防护林科技, (4): 7-9.]
CHEN LJ, LIU MQ, LIAO BY, et al., 2016. Variation of leaf morphological characters of Melia azedarach from different provenances[J]. J SW For Univ, 36(5): 16-20. [陈丽君, 刘明骞, 廖柏勇, 等, 2016. 苦楝不同种源叶片性状变异研究[J]. 西南林业大学学报, 36(5): 16-20.]
CHEN YM, 2006. Landscape dendrology[M]. Beijing: China Forestry Press: 377-379. [陈有民, 2006. 园林树木学[M]. 北京: 中国林业出版社: 377-379.]
GUO YH, CAI ZQ, CAO KF, et al., 2004. Leaf photosynthetic and anatomic acclimation of four tropical rainforest tree species to different growth light conditions[J]. J Wuhan Bot Res, 22(3):240-244. [郭玉华, 蔡志全, 曹昆芳, 等, 2004. 四种热带雨林树种光合和形态解剖特征对不同生长光强的适应[J]. 武汉植物学研究, 22(3):240-244.]
HE L, ZHOU Q, WU XS, et al., 2016. Effect of Zn on the growth of Cinnamomum camphora[J]. For Sci Technol, (11): 7-9. [贺珑, 周琴, 吴雪松, 等, 2016. Zn胁迫对香樟幼苗生长的影响[J]. 林业科技通讯, (11): 7-9.]
HU WJ, 2013. Variations over time and across different parts of tree and selection of optimum individual plant from main components of different chemical types oil in C. camphora[D]. Nanjing: Nanjing Forestry University: 109-116. [胡文杰, 2013. 樟树不同化学型精油主成分时空变异规律及优良单株选择[D]. 南京: 南京林业大学: 109-116.]
HUANG YP, 2008. Studies on Cinnamomum camphora (L.)Presl pedigree in test forest[J]. J Sanming Univ, 25(4): 426-431. [黄云鹏, 2008. 樟树家系子代测定林试验的初步研究[J]. 三明学院学报, 25(4): 426-431.]
LI ZY, JIANG Y, WAN W, et al., 2013. A study on the effects of cutting seasons on Cinnamomum camphoras large scale field cutting seedings[J]. Acta Agric Univ Jiangxi, 35 (3) : 451-455. [黎祖尧, 姜漾, 万文, 等, 2013. 季节对芳樟规模化大田扦插育苗的影响研究[J]. 江西农业大学学报, 35(3) : 451-455.]
LU XS, HE Q, 1999. The genetic variation of population and the analysis for gene diversity[J]. Acta Prat Sin, 8(3): 76-82. [卢欣石, 何琪, 1999. 种群遗传变异及基因多样度分析[J]. 草业学报, 8(3): 76-82.]
MA JM, ZHANG XZ, LIANG SC, et al., 2012. Leaf traits of common plants in Yaoshan Mountain of Guilin, China[J]. J Guangxi Norm Univ, 30(1): 77-82. [马姜明, 张秀珍, 梁士楚, 等, 2012. 桂林尧山常见植物叶片性状研究[J]. 广西师范大学学报, 30(1): 77-82.]
REN HJ, FENG Z, QIAO Q, et al., 2017. Geographic variation trend of leaf morphology in Acer truncatum[J]. J NW For Univ, 32(6): 113-119.[任红剑, 丰震, 乔谦, 等, 2017. 元宝枫叶片形态特征的地理变异[J]. 西北林学院学报, 32(6): 113-119.]
SUN XY, YAO XH, REN HD, et al.,1999. Growth variance and character analysis of Cinnamomum camphora seedling from different provenances[J]. J Zhejiang For Coll, 16(3): 234-237. [孙祥银, 姚小华, 任华东, 等, 1999. 樟树种源苗期差异及性状相关[J]. 浙江林学院学报, 16(3): 234-237.]
WANG CX, 2006. Study on Cinnamomum camphora Presl new germplasm with cold resistance through genetic engineering[D]. Taian: Shandong Agricultural University: 7-28. [王长宪, 2006. 利用基因工程创造香樟抗寒树种新种质技术研究[D]. 泰安: 山东农业大学: 7-28.]
WANG JJ, 2015. A new variety of Cinnamomum camphora ‘Xiaguang[J]. Sci Silv Sin, 51(6): 163. [王建军, 2015. 香樟新品种‘霞光[J]. 林业科学, 51(6): 163.]
WANG LB, ZHU NH, E JH, 2010. Effects of heavy matels lead, zinc and copper on young seedling growth of Cinnamomum camphora and Koelreuteria paniculata[J]. J CS Univ For & Technol, 30(2): 44-47. [王利宝, 朱宁华, 鄂建华, 2010. Pb、Zn等重金属对樟树、栾树幼苗生长的影响[J]. 中南林业科技大学学报, 30(2): 44-47.]
XIAO ZF, TAN CC, LIU XY, et al., 2017. Impact of juvenility, IBA, season on the adventitious rooting of leafy cuttings in Cinnamomum camphora[J]. Acta Agric Univ Jiangxi, 39(5): 907-912.[肖祖飞, 谭城城, 刘香玉, 等, 2017. 童性、IBA和扦插季节对香樟绿枝扦插生根的影响[J]. 江西农业大学学报, 39(5): 907-912.]
XU F, GUO WH, XU WH, et al., 2009. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves[J]. Prog Nat Sci, 19: 1789-1798.
XUE XM, XIE CP, SUN XM, et al., 2016. Comparason on anatomical structure and FTIR spectra between Cinnamomum camphora and Phoebe zhennan woods[J]. J Sichuan Agric Univ, 34(2): 178-184. [薛曉明, 谢春平, 孙小苗, 等, 2016. 樟和楠木的木材解剖结构特征和红外光谱比较研究[J]. 四川农业大学学报, 34(2): 178-184.]
YAO XH, 2002. Study on the genetic variation and selection of Cinnamomum camphora[D]. Changsha: Central South Forestry College: 95-103. [姚小华, 2002. 樟树遗传变异与选择的研究[D]. 长沙: 中南林学院: 95-103.]
YAO XH, REN HD, WU KJ, et al.,1999. The study on genetic variance at seedling stage of Cinnamomum camphora L.[J]. Acta Agric Univ Jiangxi (Nat Sci Ed), 21(3): 320-328. [姚小华, 任华东, 吴柯久, 等, 1999. 樟树苗期遗传变异研究[J]. 江西农业大学学报(自然科学版), 21(3): 320-328.]
YE RY, TONG ZK, ZHANG JH, et al., 2016. Tissue culture and rapid propagation for stems of Cinnamomum camphora[J]. J Zhejiang Agric & For Univ, 33(1): 177-182.[叶润燕, 童再康, 张俊红, 等, 2016. 樟树茎段组培快繁[J]. 浙江农林大学学报, 33(1): 177-182.]
ZHANG L, JIN ZN, LI ZY, et al., 2017. Influence of environments and geographies on properties and structures of Cinnamomum camphora seeds[J]. J CS Univ For & Technol, 37(5): 15-20. [张雷, 金志农, 黎祖尧, 等, 2017. 生境和地理因子对樟树种子性状及结构模型的影响[J]. 中南林业科技大学学报, 37(5): 15-20.]
ZHANG SY, DU FG, WANG RJ, et al., 2014. Numerical analysis on leaf morphology of Magnolia sieboldii[J]. J Beihua Univ (Nat Sci Ed), 15(2): 240-242. [张淑媛, 杜凤国, 王瑞俭, 等, 2014. 天女木兰叶片形态特征的数值分析[J]. 北华大学学报(自然科学版), 15(2): 240-242.]
ZHENG W, YAN WD, LIANG XC, et al., 2013. Effect of nitrogen addition on tree growth in subtropical Cinnamomum camphora forest[J]. J CS Univ For & Technol, 33(4):34-37. [鄭威, 闫文德, 梁小翠, 等, 2013. 氮添加对樟树林生长的影响[J]. 中南林业科技大学学报, 33(4): 34-37.]
(责任编辑何永艳)
猜你喜欢
中学生百科·小文艺(2020年8期)2020-01-13
北方音乐(2019年6期)2019-07-16
湖北农业科学(2018年8期)2018-06-08
安徽农学通报(2017年12期)2017-07-13
作文与考试·小学高年级版(2016年18期)2016-09-10
扬子江(2016年1期)2016-05-19
吉林农业(2016年7期)2016-05-14
江苏农业科学(2014年10期)2014-11-22
