
不同钙离子浓度对喜钙和嫌钙型金花茶光合及生理指标的影响
2021-03-30 05:45柴胜丰付嵘邹蓉唐健民史艳财韦记青
广西植物 2021年2期
柴胜丰 付嵘 邹蓉 唐健民 史艳财 韦记青

摘要:为探讨喜钙型金花茶对高钙环境的生理适应机制以及嫌钙型金花茶的避钙机理,该文采用砂培法,分别以两种喜钙型金花茶 [直脉金花茶(Camellia multipetala)、柠檬金花茶(C. limonia)]和两种嫌钙型金花茶 [金花茶(C. nitidissima)、东兴金花茶(C. tunghinensis)]的幼苗为材料,用不同浓度(设为5、25、50、100 mmol·L1)的钙离子(Ca2+)营养液进行培养,研究其对两种不同类型金花茶的光合及生理指标的影响。结果表明:两种嫌钙型金花茶的净光合速率(Pn)、气孔导度(Gs)、实际光化学效率(ΦPSⅡ)、电子传递效率(ETR)、叶绿素a(Chl a)、叶绿素b(Chl b)、叶绿素总量[(Chl(a+b)]均随Ca2+浓度的升高而降低,表明高钙环境降低了嫌钙型金花茶的光合能力;喜钙型金花茶的这些指标在不同Ca2+浓度下无显著变化,高钙环境未影响其光合作用的正常进行。嫌钙型金花茶叶片丙二醛含量和PSⅡ最大光化学效率(Fv/Fm)在高钙环境下未有显著变化,表明其光合膜系统还未受到伤害。随着Ca2+浓度的升高,嫌钙型金花茶叶片脯氨酸含量显著增加,而可溶性糖含量变化却不大;喜钙型金花茶在不同Ca2+浓度下脯氨酸和可溶性糖含量的变化均不明显,但其可溶性糖含量却明显高于嫌钙型金花茶。喜钙型金花茶对外界钙离子浓度的变化不敏感,其适应高钙的生理机制可能与叶片較高的可溶性糖含量有关;而嫌钙型金花茶对高钙环境的适应性较差,这可能是其不能在钙质土上生长的主要原因。
关键词: 金花茶, 喜钙, 嫌钙, 光合, 荧光, 生理
中图分类号:Q945.79
文献标识码:A
文章编号:10003142(2021)02016710
Abstract:In order to explore the physiological adaptation mechanism of calcicoletype golden Camellia to high calciumion environment and analyze the main reasons that the calcifugetype golden Camellia can not grow on calcareous soil,a pot experiment was conducted with two species of calcicoletype golden Camellia (Camellia multipetala and C. limonia) and two species of calcifugetype golden Camellia (C. nitidissima and C. tunghinensis) seedlings. To exclude the influences from other confounding factors, each pot was filled with sand instead of soil. The seedlings were treated with four Ca levels, 5, 25, 50, 100 mmol·L1. The photosynthetic and physiological indexes of two different types of golden Camellia were measured. The results were as follows: The net photosynthetic rate (Pn), stomatal conductance (Gs), actual photochemical efficiency(ΦPS Ⅱ), electron transfer efficiency (ETR), Chl a, Chl b and total Chl of the calcifugetype golden Camellia decreased with the increase of Ca2+ concentration, which indicates that the photosynthetic capacity of these two species decreased under the high calciumion environment; While these indexes in the calcicoletype golden Camellia did not change significantly under different Ca2+ concentrations, indicating high calciumion environment did not affect its normal photosynthesis. The content of malondialdehyde and the PS Ⅱ maximum photochemical efficiency (Fv/Fm) of the calcifugetype golden Camellia did not change significantly under high calciumion environment, indicating that its photosynthetic membrane system has not been damaged. With the increase of Ca2+ concentration, the proline content of the calcifugetype golden Camellia increased significantly, while the soluble sugar content did not change much; The proline and soluble sugar content in calcicoletype golden Camellia were not obviously changed under different Ca2+ concentration, but the soluble sugar content in calcicoletype golden Camellia was significantly higher than that of the calcifugetype golden Camellia. Calcicoletype golden Camellia were not sensitive to the external calciumion concentration, its physiological mechanism of adaptation to high calciumion may be related to the higher content of soluble sugar in leaves; However, the adaptability of calcifugetype Camellia to high calciumion environment was poor, which may be the main reason for its inability to grow on calcareous soil.
Key words: golden Camellia, calcicole, calcifuge, photosynthesis, fluorescence, physiology
钙(Ca)是植物生长必需的营养元素,是构成细胞壁和细胞膜的重要组分,对维持细胞结构的稳定性,调节无机离子运输等方面起着至关重要的作用(Kinzel,1989);同时,钙也是植物体内重要的信号分子,参与植物生长发育、光合作用电子传递和光合磷酸化、激素调控等,对其生理活动进行广泛调节(Poovaiah & Reddy,1993;Marschner,1995)。因此,植物对钙的吸收、转运和储存对其生理功能有重要影响(White & Broadley,2003)。但是,高浓度的钙离子(Ca2+)是一种细胞毒害剂,如果细胞内Ca2+浓度过高,将会与磷酸反应形成沉淀,从而扰乱与磷代谢有关的生理过程,或妨碍正常的信号转导进而影响植物生长(White & Broadley,2003;Hirschi,2004)。喀斯特喜钙植物通过一系列形态或生理功能的调整适应高钙环境,如将吸收的钙合成草酸钙晶体储藏在组织和细胞间隙内(Webb,1999;Franceschi & Nakata,2005);形成钙化根沉积一部分钙(Musetti & Favalim,2003);降低根系对钙的吸收(Mcconnaughey & Whelan,1997);或通过调节某些生理活性物质的变化来适应高钙环境(张宇斌等,2008;黄芬等,2008)。大多数生长在酸性土上的嫌钙植物对高钙环境相当敏感,有一部分嫌钙植物因土壤中过高的钙含量而影响其生长(Silva et al.,1994;Vicherová et al.,2015);另有相当一部分嫌钙植物之所以不能在钙质石灰土上生长是因为存在磷(P)或铁(Fe)的限制(Tyler,1996;Zohlen & Tyler,2000)。
金花茶组(Camellia, Sect. Chrysantha Chang)植物系山茶科山茶屬常绿灌木或小乔木,因其开黄色的茶花而著称于世,是珍稀的观赏植物和培育杂交茶花新品种的种质资源(韦霄等,2006)。金花茶组植物主要分布在我国广西西南部的防城、龙州、宁明、扶绥、大新及越南谅山等地(张本能和黄广宾,1986),分布区的植被类型主要是北热带石灰岩常绿阔叶林、石灰岩山地季雨林和南亚热带常绿阔叶林(苏宗明和莫新礼,1988),以海拨120~350 m之间的山间沟谷、溪边以及石灰岩坡麓、峰槽谷地较为常见(梁盛业,1989)。根据金花茶组植物生长土壤的不同,可分为石灰土金花茶和酸性土金花茶两大类,前者11种,分布于石灰岩山地(石山区);后者7种,分布于砂、页岩山地(土山区)(张宏达和任善湘,1998)。在自然情况下,尚未发现同一种类的金花茶可以在两类不同性质的土壤上出现(苏宗明和莫新礼,1988)。与砂页岩地区相比,石灰岩地区土壤一般具有富钙偏碱的特性。葛玉珍等(2009)研究表明,石灰土金花茶在酸性土上也能长势良好,而酸性土金花茶在石灰土上生长不良或死亡。根据植物生长对土壤钙质的依赖程度(候学煜,1954),可将分布于石灰土的金花茶归为喜钙型金花茶,而分布于酸性土的金花茶归为嫌钙型金花茶。喜钙型金花茶对高钙环境有很好的适应性,这是否与某些生理活性物质的调节有关?嫌钙型金花茶不能在钙质土上正常生长,高浓度的Ca2+是否是其长势不良的主要原因?为此,本研究以分布于两类不同土壤的喜钙型金花茶和嫌钙型金花茶为材料,通过比较两类金花茶对不同浓度外源Ca2+的光合和生理响应的差异,探讨喜钙型金花茶对高钙环境的适应机制,分析嫌钙型金花茶的避钙机理。研究结果将有助于深入认识金花茶组植物生境多样性的形成及适应机制,为这一珍贵类群的生物多样性保育提供科学依据。
1材料与方法
1.1 材料
在广西桂林市雁山区广西植物研究所温室大棚内进行试验。金花茶为阴生植物,通过黑色尼龙网遮荫, 搭建相对光强为15%的荫棚(中午光合有效辐射为250~300 mol·m2·s1),以便开展相关试验。试验材料为喜钙型金花茶和嫌钙型金花茶各两种(均为2年生扦插苗),喜钙型金花茶为直脉金花茶(Camellia multipetala)和柠檬金花茶(C. limonia),嫌钙型金花茶为金花茶(C. nitidissima)和东兴金花茶(C. tunghinensis),选取长势较好、基径、高度相对一致的苗木进行试验。
1.2 试验处理
采用盆栽控制试验,以石英砂作栽培基质,霍格兰氏(Hoaglands)改良营养液为母液,通过添加乙酸钙[Ca(CH3COO)2]配置不同Ca2+浓度的营养液(分别为5、25、50、100 mmol·L1)。其中,5 mmol·L1的 Ca2+浓度用以模拟酸性土中交换态钙含量,100 mmol·L1的Ca2+浓度用以模拟石灰土中交换态钙含量。栽培基质为60目的石英砂,使用前先将石英砂用0.5%盐酸浸泡1 d,之后用自来水清洗至中性。将洗净的石英砂装入塑料盆内(直径35 cm,高22 cm,底部带排水孔),每盆13 kg,上沿空出约2 cm,以便水和营养液的浇灌。选择阴天进行苗木移栽,移栽前将苗木根系洗净。每盆栽种3株同种苗木,每处理5盆,4个物种共计80盆240株苗木。移栽后缓苗1个月,待植株恢复生长后进行试验,缓苗期间用无钙的1/4 Hoagland和Aron营养液进行浇灌。试验开始后,每隔2 d浇灌 1次营养液,每盆300 mL,每10 d用大量的纯净水淋洗基质,以保持基质中营养浓度处于可控水平。2018年6月下旬开始试验,处理4个月后,测定各项试验指标。
1.3 测定项目和方法
试验结束后,选择植株顶端向下第3片至第5片成熟功能叶,进行气体交换参数、叶绿素荧光、叶绿素、丙二醛、脯氨酸和可溶性糖等各项指标的测定。
1.3.1 氣体交换参数的测定采用LI6400便携式光合测定系统分析仪(USA, LICOR)测定苗木在不同Ca2+浓度下的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等光合气体交换参数。测定时间为9:00—11:00,测定时光合有效辐射设置为300 mol·m2·s1,控制叶室温度为28 ℃,样本室CO2浓度为370 mmol·mol1。每处理测定8株,每株测定1片叶,取其平均值。
1.3.2 叶绿素荧光参数的测定在清晨阳光直射前选取叶片,暗适应20 min后,用MiniImagingPAM调制叶绿素荧光成像系统(德国,WALZ公司)测定叶片的叶绿素荧光参数。先用测量光(强度为0.1 mol·m2·s1)测定初始荧光(Fo),随后用饱和光6 000 mol·m2·s1脉冲(脉冲时间0.8 s)激发产生最大荧光(Fm)。用光化光(强度为55 mol·m2·s1)诱导荧光动力学曲线,测定叶片光适应下的最小荧光(Fo′)、最大荧光(Fm′)和稳定荧光(Fs),并由Wincontrol3软件计算光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)、实际光化学效率(ΦPS Ⅱ)和光合电子传递速率(ETR)。每处理测定6株,每株测定1片叶,取其平均值。
1.3.3 叶绿素含量的测定用95%乙醇提取叶片叶绿素,测定提取液在波长665、649 nm下吸光值,按公式计算出叶绿素a(Chl a)、叶绿素b(Chl b)的含量及Chl a/Chl b的比值(李合生,2000)。
1.3.4 丙二醛、脯氨酸和可溶性糖的测定丙二醛(MDA)含量采用硫代巴比妥酸比色法测定;脯氨酸(Pro)含量采用磺基水杨酸法测定;可溶性糖含量采用蒽酮比色法测定(李合生,2000)。
1.4 数据处理
对上述测定的各指标,利用SPSS 18.0软件进行方差分析及显著性检验(Duncan法,显著性水平P=0.05),用Sigma Plot 12.5绘图。
2结果与分析
2.1 不同Ca2+浓度对四种金花茶气体交换参数的影响
随着Ca2+浓度的升高,嫌钙型金花茶的净光合速率(Pn)呈降低趋势,金花茶和东兴金花茶在50和100 mmol·L1 Ca2+浓度处理下,其Pn显著低于5 mmol·L1 Ca2+浓度处理,与5 mmol·L1 Ca2+浓度处理相比,金花茶在50和100 mmol·L1 Ca2+浓度处理下的Pn分别下降31.3%和63.9%,东兴金花茶则分别下降23.3%和59.4%;两种金花茶在25 mmol·L1 Ca2+浓度处理下的Pn与5 mmol·L1 Ca2+浓度处理无显著差异(图1)。直脉金花茶和柠檬金花茶的Pn在不同Ca2+浓度处理下无显著差异(图1)。气孔导度(Gs)的变化趋势与Pn类似,两种嫌钙型金花茶的Gs随Ca2+浓度的升高而降低,而两种喜钙型金花茶在不同Ca2+浓度处理下无显著变化(图1)。金花茶和东兴金花茶的胞间CO2浓度(Ci)随Ca2+浓度的升高,呈先降低后升高趋势,且均是在50 mmol·L1 Ca2+浓度处理下最小,分别为(243.70±17.30) μmol·mol1和(261.87±15.53) μmol·mol1;直脉金花茶和柠檬金花茶在不同Ca2+浓度处理下,其Ci的变化并不明显(图1)。两种嫌钙型金花茶的蒸腾速率(Tr)随Ca2+浓度的升高,呈先升高后降低趋势,而两种喜钙型金花茶的Tr在不同钙离子浓度下并无显著差异(图1)。
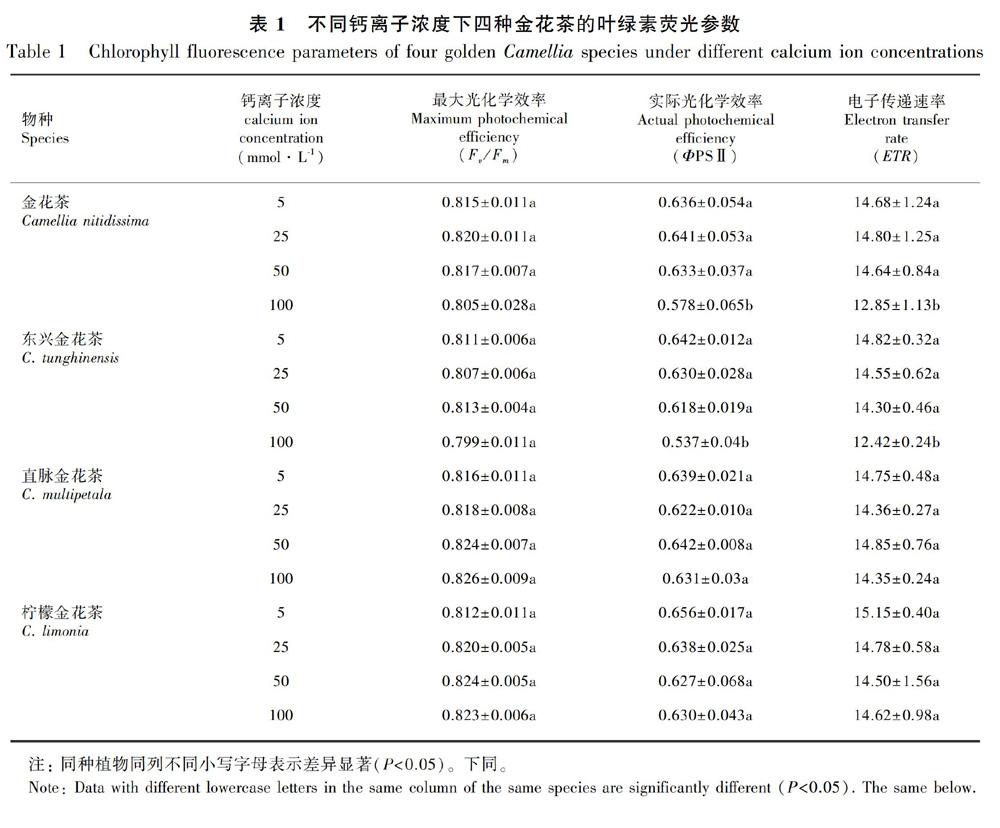
2.2 不同Ca2+浓度对四种金花茶叶绿素荧光参数的影响
随着Ca2+浓度的升高,四种金花茶的PSⅡ最大光化学效率(Fv/Fm)无显著变化,基本都在0.8以上(表1),表明四种金花茶的PSⅡ原初光能转化效率并未明显降低,其PSⅡ反应中心还未受到伤害。两种嫌钙型金花茶的实际光化学效率ΦPS Ⅱ和电子传递效率ETR随Ca2+浓度的升高呈降低趋势,表明高钙处理降低了嫌钙型金花茶的实际光能转换效率,光合电子传递受到影响,进而降低其光合能力;喜钙型金花茶在不同Ca2+浓度处理下的ΦPS Ⅱ和ETR无显著变化,表明高钙处理并未对其光合作用的正常进行产生影响(表1)。
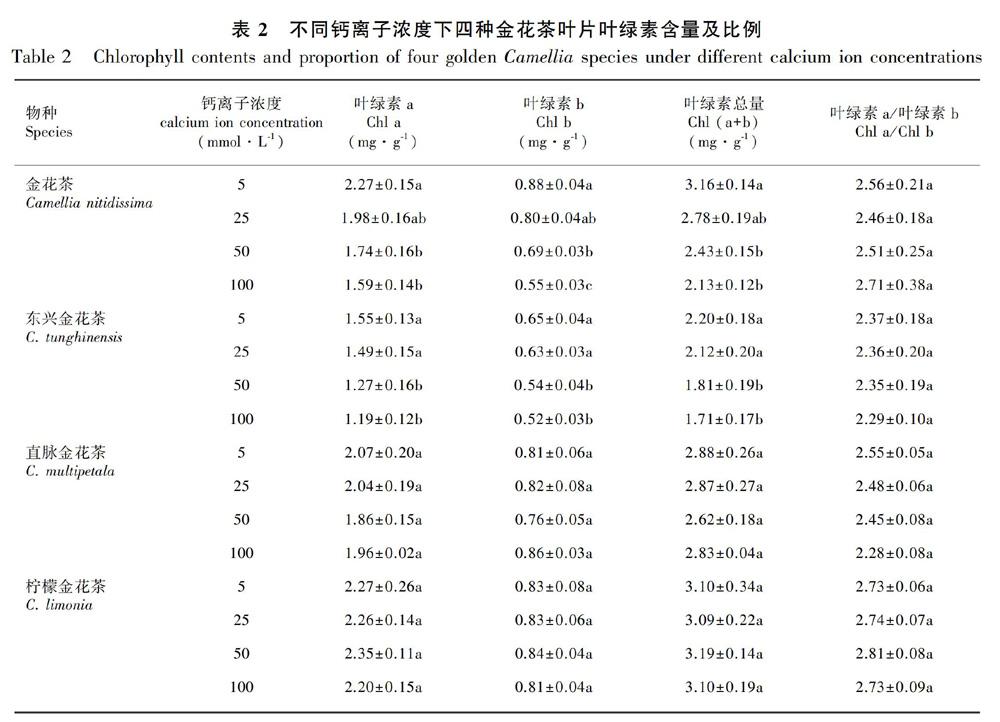
2.3 不同Ca2+浓度对四种金花茶叶片叶绿素含量的影响
叶片中的光合色素参与光合作用过程中光能的吸收、传递和转化,光合色素含量直接影响植物的光合能力。随着Ca2+浓度的升高,两种嫌钙型金花茶叶片的Chl a、Chl b和Chl (a+b)含量均呈降低趋势,在50和100 mmol·L1 Ca2+浓度处理下3个指标均显著低于5 mmol·L1 Ca2+浓度处理;Chl a/Chl b的值在不同Ca2+浓度下并无显著差异(表2)。两种喜钙型金花茶在不同Ca2+浓度处理下的Chl a、Chl b、Chl (a+b)、Chl a/Chl b值的变化并不明显(表2)。
2.4 不同Ca2+浓度对四种金花茶叶片丙二醛、脯氨酸和可溶性糖含量的影响
Data with different lowercase letters between different treatments in the same species represent significant differences at 0.05 level.
之一,其含量的高低在一定程度上反映脂膜过氧化作用水平和膜结构的受害程度。在不同Ca2+浓度处理下,四种金花茶的MDA含量均无显著差异(表3),表明高钙处理还未使四种金花茶的光合机构膜系统受到明显破坏。
脯氨酸(Pro)具有较强的水合力,它的积累可增加植物的抗旱或抗渗透胁迫能力。两种嫌钙型金花茶的Pro含量均随Ca2+浓度的升高而显著增加,与5 mmol·L1 Ca2+浓度处理相比,25、50和100 mmol·L1 Ca2+浓度处理下金花茶叶片的Pro含量分别提高了0.70、1.31和1.94倍,东兴金花茶分别提高了0.89、2.01、3.28倍;而两种喜钙型金花茶的Pro含量在不同Ca2+浓度处理下变化并不大(表3)。
两种嫌钙型金花茶的可溶性糖含量随Ca2+浓度的升高表现为先降低后升高趋势;喜钙型金花茶中,直脉金花茶在不同Ca2+浓度下无明显变化,柠檬金花茶则表现为先升高后降低趋势(表3)。两种喜钙型金花茶的可溶性糖含量明显高于嫌钙型金花茶,这可能与其适应高钙环境有关。
3讨论与结论
不同类型植物对外源Ca2+浓度的需求存在差异,喜钙植物通常在整个生活史或某一生长发育阶段强烈依赖土壤中的高Ca2+浓度环境,表现出嗜钙特性,而嫌钙植物在高Ca2+浓度环境下长势不良(苏迪等,2012)。本研究中,两种喜钙型金花茶的Pn、Gs、Ci、Tr等光合参数几乎不受外界钙离子浓度的影响,在5~100 mmol·L1 Ca2+浓度下,这几个指标几乎保持稳定,表明喜钙型金花茶对外界钙离子浓度的变化并不敏感。这与其他一些喜钙植物在不同钙离子浓度下的表现并不完全一致, 如喜钙树种伞花木(Eurycorymbus cavaleriei)随外源钙离子浓度的升高,其叶长、叶宽、叶形指数、植株高度和茎粗等指标均显著增加(苏迪等,2012);三种喀斯特喜钙苔藓植物必须在一定的Ca2+浓度下才能生长良好,在低钙环境下不能生长或长势不良(陈蓉蓉等,1998);随着Ca2+浓度的升高,石灰土专属种柳叶蕨(Cyrtogonellum Ching)的Pn在30 mmol·L1 Ca2+浓度处理下最高,在100和200 mmol·L1 Ca2+浓度处理下呈下降趋势(罗绪强等,2013)。表明不同喜钙植物对钙的需求并不完全一致。
随Ca2+浓度的升高,两种嫌钙型金花茶的Pn和Gs呈降低趋势,Ci表现为先降低后升高,Tr则表现为先升高后降低。与5 mmol·L1 Ca2+浓度处理相比,25 mmol·L1 Ca2+浓度处理下的Pn、Gs和Tr均无显著变化,表明25 mmol·L1 Ca2+浓度处理并未对金花茶和东兴金花茶的生长产生不良影响;在50和100 mmol·L1 Ca2+浓度处理下,这几个指标均显著降低,表明高于50 mmol·L1 Ca2+浓度处理降低了两种嫌钙型金花茶的光合速率,使其生长受到影响,这与对其他一些嫌钙植物的研究结果相类似。随着钙离子浓度的升高,茶树(Camellia sinensis)的Pn和Gs逐渐降低,其节间距、新梢长度、展叶数、叶面积均明显低于对照(王跃华等,2010);嫌钙植物大白杜鹃(Rhododendron decorum)叶长、叶宽、叶形指数等指标随外源钙离子浓度的升高表现出逐步下降的趋势(苏迪等,2012)。在50 mmol·L1 Ca2+浓度处理下,嫌钙型金花茶Pn降低的同时伴随Ci的降低, 表明此时两
种金花茶净光合速率降低可能主要由气孔限制因素引起,而在100 mmol·L1 Ca2+浓度处理下,Pn降低伴随Ci的升高, 此时净光合速率的降低可能主要由非气孔因素引起(Farquhar & Sharkey,1982),比如与叶片叶肉细胞羧化能力或叶绿素含量降低有关(Candana & Tarhanb,2005;罗绪强等,2013)。
叶绿素荧光可以作为光合作用的探针,叶绿素荧光技术的发展为估算叶片吸收光能的分配与光合电子传递速率提供了可能(Anderson,1999;孙金春等,2011)。在100 mmol·L1 Ca2+浓度处理下,两种嫌钙型金花茶的Fv/Fm与其他3个处理相比稍有降低,但并无显著差异(P>0.05),表明高钙处理下嫌钙型金花茶叶片的光合潜力并未明显降低;这与其他一些嫌钙植物在高钙环境下Fv/Fm明显降低并不一致(王跃华等,2010;王程媛等,2012),这是否与胁迫时间不够,还未对其膜系统产生伤害有关,还有待进一步研究,该结果与高钙胁迫下其叶片MDA含量无显著升高相一致。两种嫌钙型金花茶在100 mmol·L1 Ca2+浓度处理下的ΦPSⅡ和ETR显著低于其他处理,表明高钙环境下其PSⅡ反应中心捕获光能效率和光合电子传递速率均降低,进而影响光合作用的正常进行。在不同Ca2+浓度处理下,两种喜钙型金花茶的Fv/Fm、ΦPSⅡ和ETR均未有显著差异,表明其光系统运行正常,光合作用未受影响,这与光合参数的表现相一致。
叶绿素含量是反映光合器官生理状况的重要值指标,其含量在一定程度反映了植物生长状况和光合作用能力,也可表征逆境中植物组织、器官的衰老状况(Nieva et al.,2005;梁小红等,2015)。研究表明,喜钙植物叶片中叶绿素含量随外界钙离子浓度的升高而增加,光合能力随之增强(张习敏等,2013),而嫌钙植物在过量钙胁迫下,叶片叶绿体膜受到损伤,叶绿素含量降低, 光合作用受抑制(王跃华等,2010;申加枝等,2014)。本研究中,随着Ca2+浓度的升高,嫌钙型金花茶叶片Chl a、Chl b、Chl总量均呈降低趋势,尤其是在50和100 mmol·L1 Ca2+浓度处理下,显著低于5 mmol·L1 Ca2+浓度处理,表明高钙环境下两种嫌钙型金花茶的光合色素代谢紊乱,叶绿素的合成受到抑制,造成叶片捕光能力和光合活性降低,光合能力下降。在不同Ca2+浓度下,两种喜钙型金花茶叶片叶绿素含量并未有显著差异,表明喜钙型金花茶对不同钙离子浓度的适应范围较广,这可能与低钙环境下喜钙植物仍能保持较强的钙吸收能力有关(苏迪等,2012)。
在长期的进化过程中,植物形成了许多机制抵抗外部逆境条件。在一些逆境条件下植物可以通过增加叶片脯氨酸、可溶性糖等物质的含量来增强其渗透调节能力,进而提高植物对逆境的抵抗能力(冯晓英等,2010),但也有研究认为,脯氨酸含量的升高只是植物对逆境条件的一种生理响应(李昆等,1999)。本研究中,虽然嫌钙型金花茶叶片脯氨酸含量随Ca2+浓度的升高显著增加,但可溶性糖含量总体变化并不大,表明嫌钙型金花茶对高钙环境的适应能力有限。两种喜钙型金花茶在不同Ca2+浓度下叶片脯氨酸和可溶性糖含量变化并不明显,表明其并未受到胁迫,对低钙和高钙环境都有很好的适应性,这与喜钙植物伞花木、狗骨木(Swida wilsoniana)等在不同钙环境下的表现相似(张习敏等,2013;张芳等,2017)。在不同钙离子浓度下,喜钙植物云贵鹅耳枥(Carpinus pubescens)叶片POD含量远高于嫌钙植物油茶(Camellia oleiferal),POD的高效表达保证了喜钙植物在受到环境胁迫时能很好地保护其细胞膜结构和其他生理生化过程不受影响(张宇斌等,2008)。本研究中,在不同Ca2+浓度下,喜钙型金花茶叶片可溶性糖含量明显高于嫌钙型金花茶,这可能与POD有相似的效果,以保证喜钙型金花茶能很好地適应高钙环境。
综上所述,喜钙型金花茶对外界钙离子浓度的变化并不敏感,在高钙和低钙环境下均能正常生长,其适应高钙的生理机制可能与叶片较高的可溶性糖含量有关;而嫌钙型金花茶在高钙环境下长势不良,这很可能是其不能在钙质土上生长的主要原因。
参考文献:
ANDERSON JM, 1999. Insights into the consequences of grana stacking of thylakoid membranes in vascular plants: A personal perspective [J]. Aust J Plant Physiol, (26): 625-639.
CANDANA N, TARHANB L, 2005. Effects of calciumion, stress on contents of chlorophyll and carotenoid, LPO levels, and antioxidant enzyme activities in Mentha [J]. J Plant Nutr, 28: 127-139.
CHEN RR, LIU N, YANG S, et al., 1998. The effects of Ca2+ concentration on growth of four bryophytes from karst area on Qianling mountain [J]. J Guizhou Norm Univ (Nat Sci Ed), 16(1): 4-7.[陈蓉蓉, 刘宁, 杨松, 等, 1998. Ca2+浓度对黔灵山喀斯特生境中几种苔藓植物生长的影响. 贵州师范大学学报(自然科学版), 16(1): 4-7.]
FARQUHAR GD, SHARKEY TD, 1982. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol, 33: 317-345.
FENG XY, HU ZP, YI Y, 2010. Variation of proline and soluble protein content in leaves of Eurycorymbus cavalerieian and Pinus armandii under Ca2+ stress [J]. Guizhou Agric Sci, 38(9): 169-170.[冯晓英, 胡章平, 乙引, 2010. Ca2+胁迫下伞花木和华山松脯氨酸及可溶性蛋白质含量的变化 [J]. 贵州农业科学, 38(9): 169-170.]
FRANCESCHI VR, NAKATA PA, 2005. Calciumion oxalate in plants: Formation and function [J]. Ann Rev Plant Biol, 56: 41-71.
GE YZ, ZOU LX, TANG GT, et al., 2009. Phenological characteristics of 7 species of Sect. Chrysantha Chang in Guilin Botany Garden [J]. Acta Agric Jiangxi, 21(4): 59-60.[葛玉珍, 邹丽霞, 唐广田, 等, 2009. 迁地保护下 7种金花茶的物候特征的初步研究 [J]. 江西农业学报, 21(4): 59-60.]
HIRSCHI KD, 2004. The calciumion conundrum, both versatile nutrient and specific signal [J]. Plant Physiol, 136: 2438-2442.
HOU XY, 1954. Indicator plants of acid soil, calcareous soil and saline alkali soil in China [M]. Beingjing: Science Press.[候學煜, 1954. 中国境内酸性土、钙质土和盐碱土的指示植物 [M]. 北京: 科学出版社.]
HUANG F, ZHU MJ, LU Q, et al., 2012. Comparison study on several stress resistance indexes in plant under different calciumion contents environment [J]. J Guangxi Norm Univ(Nat Sci Ed), 30(4): 98-103.[黄芬, 朱敏洁, 卢茜, 等, 2012. 不同钙环境下植物几种抗逆境指标的对比研究. 广西师范大学学报(自然科学版), 30(4): 98-103.]
KINZEL H, 1989. Calciumion in the vacuoles and cell walls of plant tissue [J]. Flora, 182: 99-125.
LI HS, 2000. The experiment principle and technique on plant physiology and biochemistry[M]. Beijing: Higher Education Press.[李合生, 2000. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社.]
LI K, ZENG JM, ZHAO H, 1999. Drought resistance in relation to free proline in the major tree species in dry and hot river valley of Jinsha [J]. For Res, 12(1): 103-107.[李昆, 曾觉民,赵虹, 1999. 金沙江干热河谷造林树种游离脯氨酸含量与抗旱性关系 [J]. 林业科学研究, 12(1): 103-107.]
LIANG SY, 1989. Classification of Camellia section Chrysantha and their ecological geographical distribution characteristics [J]. Guangxi For Sci, (1):1-3.[梁盛业, 1989. 金花茶植物分类研究及其生态地理分布特点 [J]. 广西林业科学, (1):1-3.]
LIANG XH, AN MY, SONG Z, et al, 2015. Effects of exogenousglycine betaine on the physiological characteristics of Zoysia japonica under lowtemperature stress [J]. Acta Pratacul Sin, 24(9): 181-188.[梁小红, 安勐颍, 宋峥, 等, 2015. 外源甜菜碱对低温胁迫下结缕草生理特性的影响 [J]. 草业学报, 24(9): 181-188.]
LUO XQ, WANG SJ, ZHANG GL, et al., 2013. Effects of calciumion concentration on photosynthesis characteristics of two fern plants [J]. Ecol Environ Sci, 22(2): 258-262.[罗绪强, 王世杰, 张桂玲, 等, 2013. 钙离子浓度对两种蕨类植物光合作用的影响 [J]. 生态环境学报, 22(2): 258-262.]
MARSCHNER H, 1995. Mineral nutrition of higher plants.[M]. 2nd ed. London: Academic Press.
MCCONNAUGHEY TA, WHELAN JF, 1997. Calcification generates protons for nutrient and bicarbonate uptake [J]. Earth Sci Rev, 42: 95-117.
MUSETTI R, FAVALIM A, 2003. Cytochemical localization of calciumion and Xray microanalysis of Catharanthus roseus L. infected with phytoplasmas [J]. Micron, 34( 8): 387-393.
NIEVA FJJ, CASYILLO JM, LUQUE CJ, 2005. Ecophysiology of tidal and nontidal population of the invading cordgrass Spartina densiflora: Seasonal and diurnal patterns in a Mediterranean climate [J]. Estuar Coast Shelf Sci, 57: 919-928.
POOVAIAH HW, REDDY ASN, 1993. Calciumion and signal transduction in plants [J].Crit Rev Plant Sci, 12: 185-211.
SHEN JZ, ZHANG XF, WANG Y, et al., 2014. Dynamic effects of excessive calciumion on biochemical components in young shoots and root growth of tea [J]. Shandong Agric Sci, 46(6): 85-88.[申加枝, 張新富, 王玉, 等, 2014. 钙过量对茶树幼苗叶绿素组成及钙、镁吸收的动态影响 [J]. 山东农业科学, 46(6): 85-88.]
SILVA D, RUIZ LP, ATKINSON CJ, et al., 1994. Physiological disturbances caused by high rhizospheric calciumion in the calcifuge Lupinus luteus [J]. J Exp Bot, 45(5): 585-590.
SU D, YI Y, ZHANG XM, et al., 2012. Effect of exogenous Ca2+ on growth of Eurycorymbus cavaleriei and Rhododendron decorum [J]. Guizhou For Sci Technol, 40(3): 23-27. [苏迪, 乙引, 张习敏, 等, 2012. 外源Ca2+对伞花木和大白杜鹃生长及矿质元素含量代谢的影响 [J]. 贵州林业科技, 40(3): 23-27.]
SU ZM, MO XL, 1988. Geographic distribution of Camellia section Chrysantha form China [J]. Guihaia, 8(1):75-81.[苏宗明, 莫新礼, 1988. 我国金花茶组植物的地理分布 [J]. 广西植物, 8(1): 75-81.]
SUN JC, ZHANG YH, WEN Q, et al., 2011. Effects ofdifferent calciumion reagents on photosynthesis characteristics of Catharanthus roseus [J]. J SW Univ (Nat Sci Ed), 33(6): 74-78.[孙金春, 张扬欢, 温泉, 等, 2011. 不同钙效应剂对长春花光合特性的影响 [J]. 西南大学学报(自然科学版), 33(6): 74-78.]
TYLER D, 1996. Mineral nutrient limitations of calcifuge plants in phosphate sufficient limestone soil [J]. Ann Bot, 77: 649-656.
VICHEROV E, HJEK M, HJEK T, 2015. Calciumion intolerance of fen mosses: Physiological evidence, effects of nutrient availability and successional drivers [J]. Perspect Plant Ecol, 17: 347-359.
WANG XY, WANG SY, RONG Y, 2012. The effects of nutrient stress on chlorophyll fluorescence characteristics of Diplazium pinfaense [J]. Earth Environ, 40(1): 23-29.[王程媛, 王世杰, 容丽, 2012. 营养胁迫对薄叶双盖蕨叶绿素荧光特征的影响 [J]. 地球与环境, 40(1): 23-29.]
WANG YH, ZHANG LX, SUN QY, 2010. Effects of excessive calciumion fertilization on photosynthetic characteristics and chloroplast ultrastructure of tea tree [J]. Plant Nut Fert Sci, 16(2): 432-438.[王跃华, 张丽霞, 孙其远, 2010. 钙过量对茶树光合特性及叶绿体超微结构的影响 [J]. 植物营养与肥料学报, 16(2): 432-438.]
WEBB MA, 1999. Cellmediated crystallization of calciumion oxalate in plants [J]. Plant Cell Online, 11(4): 751-760.
WEI X, JIANG SY, JIANG YS, et al., 2006. Researchprogress of Camellia nitidissima, a rare and endangered plant [J]. J Fujian For Sci Technol, 33(3): 169-173.[韦霄, 蒋水元, 蒋运生, 等, 2006. 珍稀濒危植物金花茶研究进展 [J]. 福建林业科技, 33(3): 169-173.]
WHITE PJ, BROADLEY MR, 2003. Calciumion in plants [J]. Ann Bot, 92: 487-511.
ZHANG BN, HUANG GB, 1986. The classification and geographic distribution of golden Camellias [J]. J Wuhan Bot Res, 4(1): 31-42.[张本能, 黄广宾, 1986. 金花茶的分类和地理分布 [J]. 武汉植物学研究, 4(1): 31-42.]
ZHANG F, SONG M, PENG WX, et al., 2017. Effects of different calciumion concentrations on seedling growth and enzyme activities of two karst plant species [J]. Guihaia, 37(6): 707-715.[张芳, 宋敏, 彭晚霞, 等, 2017. 不同钙离子浓度对两种岩溶植物幼苗生长及其酶活性的影响 [J]. 广西植物, 37(6): 707-715.]
ZHANG HD, REN SX, 1998. Flora Reipublicae Popularis Sinicae (the third fascicle) in fortyninth volume[M]. Beijing: Science Press.[张宏达, 任善湘, 1998. 中国植物志 (第49卷第3分册) [M]. 北京: 科学出版社.]
ZHANG XM, SONG QF, LIU LX, et al., 2013. Responses ofgrowth and physiology calcicoles and calcifuges to exogenous Ca2+ [J]. Acta Bot BorealOccident Sin, 33(8): 1645-1650.[張习敏, 宋庆发, 刘伦衔, 等, 2013. 喜钙和嫌钙植物对外源Ca2+的生长生理响应 [J]. 西北植物学报, 33(8): 1645-1650.]
ZHANG YB, ZHANG R, FENG L, et al., 2008. Effect of external calciumion on POD activity of calciphile and calcifuge [J]. J Guizhou Norm Univ (Nat Sci Ed), 26(3): 10-12.[张宇斌, 张荣, 冯丽, 等, 2008. 外源 Ca2+对喜钙和嫌钙植物POD活性的影响 [J]. 贵州师范大学学报(自然科学版), 26(3): 10-12.]
ZOHLEN A, TYLER G, 2000. Immobilization of tissue iron on calcareous soil: Differences betweencalcicole and calcifuge plants [J]. Oikos, 89: 95-106.
(责任编辑蒋巧媛)
猜你喜欢
音乐探索(2022年2期)2022-05-30
中国药学药品知识仓库(2022年9期)2022-05-23
佛山陶瓷(2022年1期)2022-02-19
江苏农业学报(2019年1期)2019-09-10
小资CHIC!ELEGANCE(2018年28期)2018-09-14
Coco薇(2017年5期)2017-06-05
分析化学(2016年12期)2017-02-04
小资CHIC!ELEGANCE(2016年15期)2016-07-26
百科知识(2015年12期)2015-09-10
环球时报(2015-05-13)2015-05-13
