
Na2SO4和Na2CO3胁迫下苦楝幼苗的形态及光合生理特性
2021-03-30 05:45张远兰胡鑫蔡金峰国靖郁万文曹福亮汪贵斌
广西植物 2021年2期
关键词:生长
张远兰 胡鑫 蔡金峰 国靖 郁万文 曹福亮 汪贵斌
摘要:為探索苦楝应对盐胁迫的响应机制,该文以1年生苦楝(Melia azedarach)实生苗为材料,在盆栽条件下设置中性盐
Na2SO4和碱性盐Na2CO3 3个盐浓度(200、400、600 mmol·L1)处理40 d,研究苦楝的抗盐碱水平及在不同程度盐碱胁迫条件下的生长及光合生理变化。结果表明:随着盐浓度的提高,苦楝的苗高、地径和生物量的增长量均呈现下降趋势,且碱性盐胁迫条件下降程度更大,盐胁迫提高苦楝的根冠比。处理10 d时,苦楝幼苗的所有光合指标随中性盐和碱性盐浓度的提高呈相似的下降特征,碱性盐胁迫条件下的降低幅度显著大于中性盐胁迫,且随处理时间的增加,中性盐和碱性盐处理下苦楝幼苗的净光合速率和蒸腾速率显著降低。随着盐浓度的提高,苦楝的叶绿素含量呈现下降趋势,200 mmol·L1盐胁迫对叶绿素含量影响较小,400、600 mmol·L1盐胁迫均对叶绿素含量有显著影响。600 mmol·L1碱性盐胁迫条件下,苦楝叶片相对电导率和饱和水分亏缺最高,显著高于其余处理。同等浓度下,碱性盐胁迫的苦楝叶片相对电导率和饱和水分亏缺显著高于中性盐胁迫处理。综上结果认为,苦楝具有一定的耐盐碱能力,碱性盐比中性盐对苦楝幼苗的影响更大。
关键词: 苦楝, 盐碱胁迫, 生长, 光合, 水分状况
中图分类号:Q945.78
文献标识码:A
文章编号:10003142(2021)02019511
Abstract:To explore the response mechanism of Melia azedarach to salt stress and to provide reference for the application of M. azedarach in saline and alkaline areas. Oneyearold M. azedarach seedlings grown in pots were used as materials, and subjected to sodium sulphate or sodium carbonate of three concentrations (200, 400, 600 mmol·L1) for 40 d to study the growth, physiological and photosynthetic parameters. The results were as follows: The growth of seedling height, ground diameter and biomass showed downward trends with the increase of salt concentration under neutral or alkaline salt stress, and the decline was greater under alkaline salt stress. Salt stress increased the root to shoot ratio. On the 10th day, all photosynthetic parameters showed similar decline characteristics under neutral or alkaline salt stress, and the decrease extent under alkaline salt stress was significantly greater than those under neutral salt stress. With the increase of treatment time, the net photosynthetic rate and transpiration rate decreased significantly under neutral salt or alkaline salt. With the increase of salt concentration, the chlorophyll content showed a downward trend. Salt stress of 200 mmol·L1 had little effect on the chlorophyll content, but 400 and 600 mmol·L1 salt stress had significant effect on the chlorophyll content. Under 600 mmol·L1 alkaline salt stress, the relative conductivity and saturated water deficit of M. azedarach leaves were the highest, significantly higher than those of other treatments. At the same concentration, the relative conductivity and saturated water deficit under alkaline salt stress were significantly higher than those under neutral salt stress. It can be seen that M. azedarach has a certain saltalkali tolerance, the effect of alkaline salt on the M. azedarach seedlings is greater than neutral salt.
Key words: Melia azedarach, saltalkali stress, growth, photosynthesis, water status
土壤盐碱化是指土壤中可溶性盐分(硫酸钠、氯化钠、碳酸钠和碳酸氢钠等)不断向土壤表层积聚形成盐碱土的过程,是农业生态环境退化中的一个主要问题。据统计,全球有超过8亿公顷的土地受到盐(3.97亿公顷)或盐碱(4.34亿公顷)的影响(Munns,2005)。其中由于人类不合理的灌溉,导致地下水中的盐分沿土壤毛管孔隙上升并在地表积累引起的盐渍化称为次生盐渍化,主要发生在干旱和半干旱地区。由于海潮发生带入土壤中大量盐类,在强烈蒸发作用下盐类向地表积累而形成的盐渍化称为滨海盐渍化,主要发生在滨海地区。盐胁迫通常会限制植物的生长,降低土壤肥力,影响作物产量,并给当地人们的生产和生活、经济发展乃至生态环境带来严重威胁(刘正祥等,2014)。治理盐碱化主要是通过引淡淋盐、井排井灌、覆盖、换土和种植耐盐植物等手段,其中种植耐盐植物不仅成本较低且改良效果好,且能够保持水土和提高土壤质量,为盐碱地的有效利用和生态可持续发展提供基础(单奇华等,2011)。
苦楝(Melia azedarach)为楝科楝属落叶乔木,是我国优良的乡土树种,又名苦苓、楝树和森树等(程诗明等,2005)。苦楝是优良的材用树种,其材质坚韧、纹理华美、光洁度高且耐磨性强,亦是药用树种,其花、叶、果实和根皮均可入药(陈丽君等,2014)。此外,苦楝还具有较好的涵养水源、保持水土和抗盐等功能,是东部滨海盐渍区优良的造林树种之一(张子晗等,2014)。研究表明苦楝有较强的抗盐能力,能够在含盐量0.46%以下的盐碱地正常生长(王家源,2013)。硫酸钠和氯化钠是在盐渍土壤中占主导地位的中性盐类,Cl-和SO2-4离子在物理化学行为上截然不同,SO2-4具有较大的水化直径,比土壤溶液中的Cl-更容易形成离子对(Li et al., 2019)。到目前为止,已经有关于氯化钠对苦楝胁迫的研究。苗海霞等(2005)研究表明NaCl胁迫显著抑制苦楝的生长,对地上部分影响更大,且高浓度的盐分胁迫对叶的影响大于对茎的影响;魏海霞等(2005)研究表明苦楝对0.2%浓度的NaCl有一定抗性,脯氨酸是高盐胁迫下苦楝最重要的渗透调节物质;Xu et al. (2018)对6个种源苦楝耐盐性进行研究,表明不同种源苦楝幼苗的脯氨酸、丙二醛和可溶性蛋白含量差异较大,且耐盐性差异较大;Yang et al.(2008)认为碳酸钠和碳酸氢钠是主要的碱性盐成分,其破坏因子与中性盐胁迫类似,又有高pH的附加胁迫。目前关于苦楝对碱性盐耐受能力的研究较少。因此,本研究选择中性盐硫酸钠和碱性盐碳酸钠模拟盐胁迫,以1年生苦楝实生苗为研究对象,测定不同浓度中性盐和碱性盐对苦楝幼苗的生长和光合作用指标的影响,以期了解苦楝的耐硫酸盐和碳酸盐的能力,为其在滨海盐碱地区的开发利用提供基础。
1材料与方法
1.1 材料
试验材料为1年生苦楝实生苗,2018年在南京林业大学下蜀林场育苗大棚中进行试验。2016年年底于南京林业大学校园20年生苦楝母株上采集果实,果实经浸泡、揉搓和去壳后筛选出饱满籽粒,播种育苗后于2017年4月选取长势良好且相对一致的苦楝幼苗进行盆栽。盆栽容器为高28 cm、直径32 cm的聚乙烯塑料盆,每盆栽植苦楝幼苗1棵,每盆装大田土壤4 kg,苗木生长期间正常浇水和管理维护。2018年2月进行平茬,4月份挑选生长相对一致苗木进行处理。
1.2 试验设计
本研究结合江苏地区盐碱地土壤盐分的含量及前人对苦楝耐NaCl盐胁迫的研究设定浓度 (魏海霞等,2005;Xu et al., 2018),并按照Na2SO4与Na2CO3不同浓度处理进行分组,分为CK (0)、低盐浓度(200)、中盐浓度(400)和高盐浓度(600) mmol·L1共7个随机分组,每个处理13棵苗,其中每个处理中5棵苗用于非破坏性测定。为避免盐激反应,同时保证盐分的均匀分布,整个施盐过程分3 d完成,每天用相应浓度的盐溶液浇灌,盐溶液量为100 mL,不施盐的处理施入100 mL蒸馏水,盐分施入完成即该试验处理正式进行。之后进行统一水分管理,如果有溶液渗入托盘,就将托盘中水分回灌至塑料盆中,保持土壤的盐分不流失。
1.3 指标及测定方法
1.3.1 苗高、地径和生物量测定处理第0、第10、第20、第30和40天分别用钢卷尺和游标卡尺测定苗高和地径。处理40 d时,各处理分别取3株苦楝苗,自来水冲洗干净,带回实验室再用去离子水冲洗干净,用纸巾吸干水分,将苗木分为根、茎、叶片三部分,在105 ℃烘箱內杀青 10 min,再于80 ℃烘干直至恒重,分别称重测定生物量。
1.3.2 叶绿素含量测定处理第10、第20、第30和40天采样,称取0.1 g新鲜叶片放入刻度试管,并加入10 mL 80%丙酮溶液,置于黑暗处。待试管中叶片颜色彻底变为白色,使用紫外分光光度计DU800同时对浸提液进行663、645 mm波长的比色测定,从而计算得到总叶绿素含量(mg·g1FW)(王家源,2013)。
1.3.3 光合作用参数测定在处理的第10和第40天,使用Li6400便携式光合测定仪(LiCor 6400,USA)于上午9:00—11:00对苦楝进行光合参数测定。测定时使用开放气路,光强为1 400 μmol·m2·s1,CO2流量为500 μmol·m2·s1。测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。测定时选取完全展开的成熟叶片(从顶部数第4至第5片叶),每处理测定5株,每株3片。
1.3.4 相对电导率测定处理第10、第20、第30和40天采样,使用蒸馏水将取自同一位置的2 g新鲜叶片进行两次冲洗,并将其剪为1 cm左右小段至于烧杯里,用20 mL蒸馏水倒入浸没叶片。将烧杯放入真空干燥器中,进行3次抽气处理,致叶片沉于烧杯底部。取出抽气的烧杯放置3 h,进行均匀搅动,使用电导仪测量其恒温25 ℃时的电导率S1;然后将溶液进行15 min的沸水浴,待恢复常温后再次测量其电导率S2,同样环境下测量蒸馏水的电导率记为S0。计算公式如下:
1.3.5 饱和水分亏缺测定采用饱和称重法,在第10、第20、第30和40天取新鲜叶片称鲜重(Wf)后,浸入水中,待叶片充分吸水后取出,用吸水纸吸干其表面附着的水分,称其饱和鲜重(Wt),然后置于80 ℃烘箱中杀青,再置于55 ℃烘箱中烘干至恒重,称其干重(Wd)。计算公式如下:
1.4 数据处理
利用Excel 2017制作图表,利用SPSS 19.0进行数据统计和方差分析,对不同盐类型、不同盐浓度和不同处理时间的苦楝幼苗苗高、地径、光合指标和叶绿素含量等指标进行三因素方差分析。图表中数据为3次重复的平均值±标准方差(SD)。
2结果与分析
2.1 盐胁迫对苦楝苗高、地径和生物量的影响
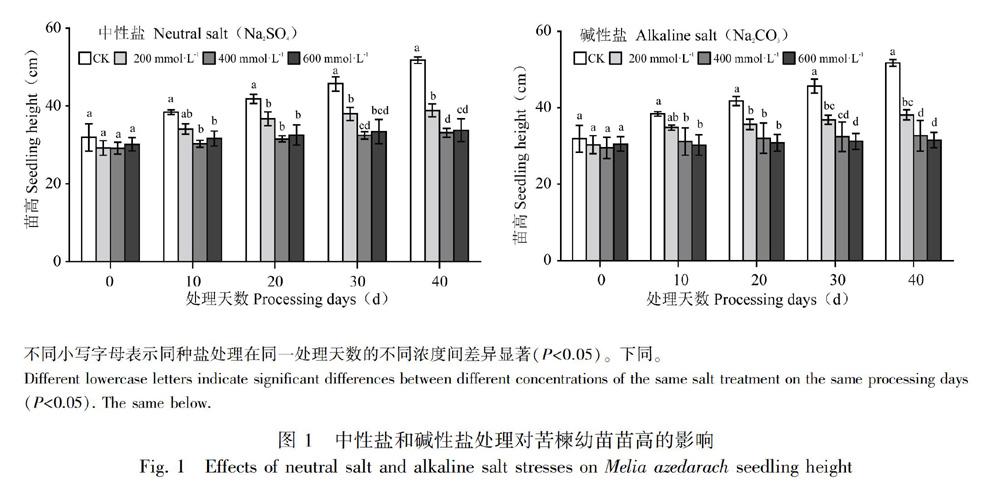
由图1可知,不同浓度中性盐和碱性盐处理均抑制了苦楝苗高的增长,且随着盐浓度的增大和胁迫时间的延长,抑制作用越强。方差分析表明,不同盐分浓度和胁迫时间对苗高影响达显著差异(P<0.05),但不同盐分类型对苦楝苗高影响差异不显著(P>0.05)。
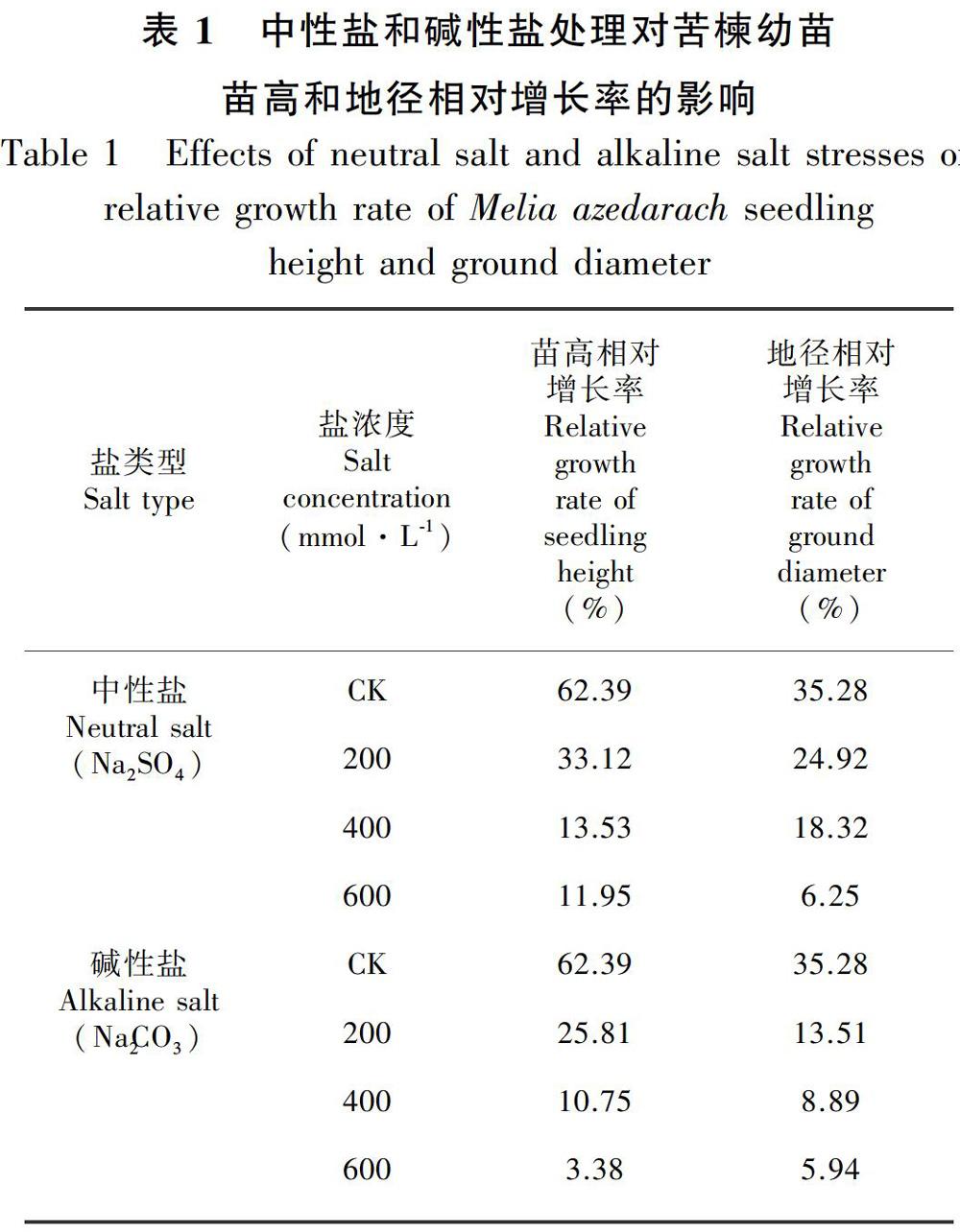
在盐胁迫处理初期(10 d),在200 mmol·L1浓度胁迫下苗高与对照差异不显著,说明低浓度盐胁迫在处理初期并没有对苦楝高生长造成影响,400、600 mmol·L1浓度胁迫下苗高显著低于对照。在盐胁迫20 d后,不同盐胁迫下苦楝苗高均显著低于对照,且各盐处理之间随着胁迫浓度的增加,高生长受到抑制程度加剧,400、600 mmol·L1浓度胁迫下苗高增长很缓慢,几乎处于停滞状态,说明中、高浓度盐碱胁迫对苦楝高生长造成显著影响。由表1可知,盐胁迫下苦楝苗高相对增长率均低于对照,各中性盐和碱性盐处理下分别比对照低46.91%、78.31%、80.85%和58.63%、82.77%、94.58%,特别是在600 mmol·L1碱性盐胁迫下,苗高增长率仅为3.38%,说明碱性盐胁迫对苗高增长率的抑制程度高于中性盐。
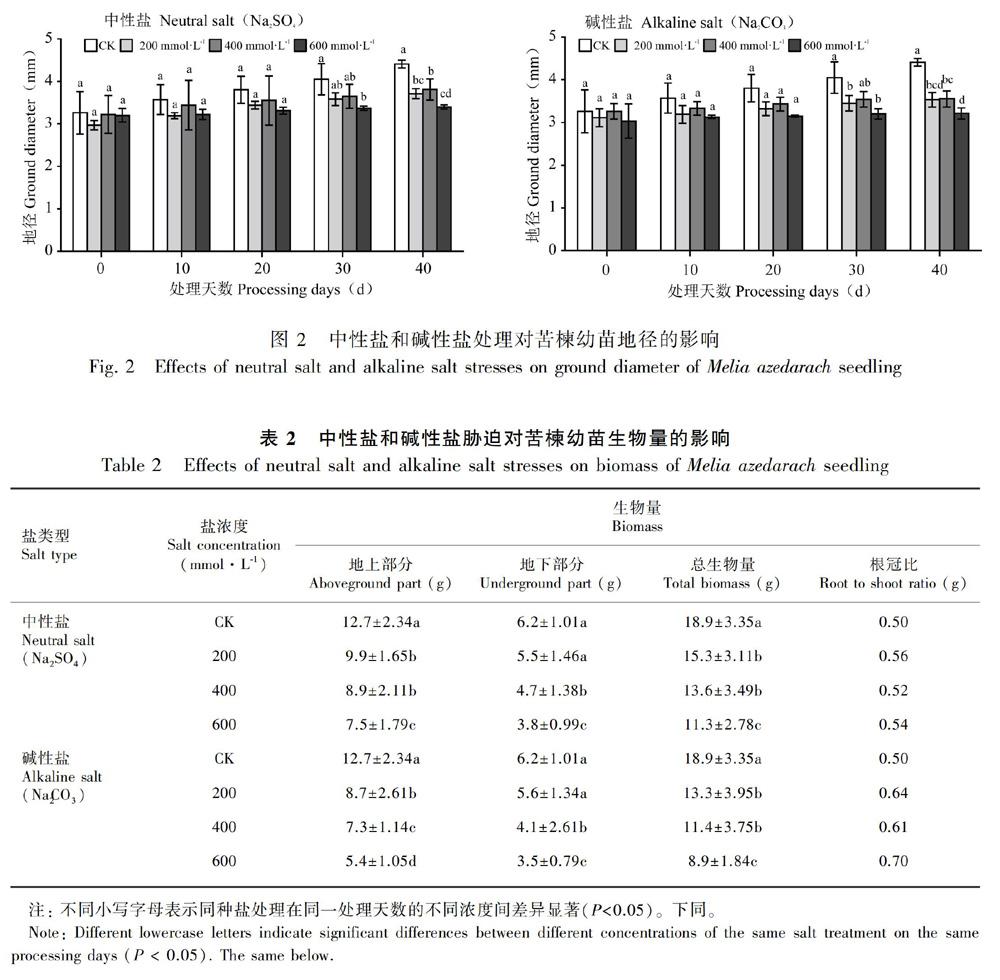
由图2可知,盐处理显著抑制了苦楝地径的增长,且随着盐浓度的提高,抑制效果呈递增的趋势。方差分析表明,不同盐分类型、盐分浓度和胁迫时间对地径影响的差异均达显著水平(P<0.05)。盐胁迫处理前20 d,各盐胁迫对苦楝地径的影响差异不显著,可能是盐胁迫在这个阶段对苦楝地径增长抑制程度不高。处理30 d 以后,各盐处理下苦楝地径显著低于对照,盐胁迫抑制地径增长的程度加剧。由表1可知,随盐胁迫浓度增加,苦楝地径相对增长率呈下降趋势,各中性盐和碱性盐胁迫下分别比对照低29.37%、48.07%、82.28%和61.71%、74.80%、83.16%,碱性盐胁迫对地径相对增长率的抑制程度高于中性盐。
由表2可知,不同盐胁迫条件对苦楝地上、地下及总生物量的积累造成一定影响,随胁迫程度增大,生物量的积累呈下降趋势。方差分析表明,不同盐浓度对苦楝地上、地下及总生物量的影响差异达显著水平(P<0.05),盐分类型对苦楝地上和总生物量的影响差异达显著水平(P<0.05),但对地下部分影响不显著。在200 mmol·L1条件下,中性盐和碱性盐处理的总生物量分别比对照低23.5%和42.1%,且均与对照存在显著差异;其中,地上部分均显著低于对照组,但地下部分与对照组无显著差异。在400 mmol·L1条件下,中性盐和碱性盐处理的总生物量分别比对照低39.0%和65.8%,且均与对照组差异显著;地上部分和地下部分均显著低于对照组。在600 mmol·L1条件下,中性盐和碱性盐处理的总生物量分别比对照低67.3%和112.4%,且均与对照存在显著差异;地上部分和地下部分均显著低于对照。总体上,碱性盐对苦楝幼苗生物量积累的影响更大。
与对照相比,不同盐胁迫下苦楝根冠比均有不同程度提高,说明盐胁迫对地上部分生长的抑制程度要大于地下部分。碱性盐胁迫下更大的根冠比说明碱性盐对苦楝地上部分有更大的抑制作用。
2.2 盐胁迫对苦楝幼苗光合作用参数的影响
由表3可知,随着盐度的提高,抑制作用递增。盐胁迫10 d后,方差分析表明,中性盐和碱性盐各浓度处理净光合速率和气孔导度均低于对照组,且均未达到显著水平(P>0.05)。中性盐和碱性盐各浓度处理蒸腾速率和胞间CO2浓度均低于对照组,且各处理间差异显著(P<0.05)。盐胁迫处理40 d后,中性盐和碱性盐各浓度处理净光合速率和气孔导度均显著低于对照组,200、400 mmol·L1处理间净光合速率和气孔导度无显著差异,均与600 mmol·L1处理存在显著差异。中性盐和碱性盐的高盐浓度处理气孔导度分别比对照低48.3%和62.3%。盐胁迫处理40 d时,苦楝幼苗的蒸腾速率和胞间CO2浓度均显著低于对照组, 随着盐浓度的增大,抑制作用也递增。方差分析表明,不同鹽分类型、浓度和胁迫时间苦楝蒸腾速率和胞间CO2浓度差异达显著水平(P<0.05)。其中200、400 mmol·L1处理间蒸腾速率和胞间CO2浓度无显著差异,但均与600 mmol·L1处理存在显著差异。中性盐和碱性盐的高盐浓度处理蒸腾速率分别比对照低42.9%和137.0%,胞间CO2浓度比对照低56.0%和106.2%。
2.3 盐胁迫对苦楝幼苗叶绿素含量的影响
由图3可知,盐胁迫处理后,随盐浓度的提高,苦楝幼苗叶绿素含量呈先升后降的变化趋势。方差分析表明,不同盐分类型、浓度和胁迫时间对苦楝叶绿素含量的影响差异达显著水平(P<0.05)。盐胁迫10 d后,中性盐和碱性盐处理苦楝叶绿素含量均与对照无显著差异。盐胁迫20 d后,中性盐和碱性盐200 mmol·L1浓度处理的叶绿素含量均显著高于对照组;中性盐和碱性盐600 mmol·L1浓度处理叶绿素含量最低,分别比对照低7.8%和16.6%。盐胁迫30 d后,中性盐各浓度处理的叶绿素显著低于对照组,碱性盐400、600 mmol·L1浓度处理叶绿素含量分别比对照低37.63%、50.11%。盐胁迫40 d后,中性盐和碱性盐400、600 mmol·L1处理叶绿素含量显著低于对照组,其中600 mmol·L1浓度处理叶绿素含量均最低,分别比对照低53.53%、56.64%。总体上,200 mmol·L1浓度的中性盐和碱性盐均对叶绿素含量影响较小,中、高浓度盐胁迫对叶绿素含量有显著影响,中、高浓度碱性盐对苦楝幼苗叶绿素含量的影响更大。
2.4 盐胁迫对苦楝幼苗相对电导率和饱和水分亏缺的影响
由图4可知,盐胁迫后,各处理苦楝的相对电导率均高于对照组,且随着盐浓度的增大,相对电导率呈递增变化趋势;在相同浓度的条件下,碱性盐处理的相对电导率高于中性盐处理的相对电导率。同时,随着盐胁迫处理时间的延长,苦楝的相对电导率呈上升变化趋势,而对照组的相对电导率一直处于相对稳定的状态。方差分析表明,不同盐分类型、浓度和处理时间对苦楝相对电导率影响达显著差异水平(P<0.05)。
3讨论与结论
植物的个体生长发育过程常会受到不同程度干旱、盐碱、冻害和高温等非生物因素的影响,其中盐碱是发生在干旱、半干旱灌溉区及滨海地区最关键的非生物胁迫因子(偶春等,2019)。盐胁迫下,植物的多个生理生化过程都会受到不同程度的抑制,其中植物的生长量可以直观地反映其对盐胁迫的耐受程度(金正律和周秋艳,2014)。本研究中,不同浓度中性盐(Na2SO4)和碱性盐(Na2CO3)处理下的苦楝幼苗苗高、地径和生物量在一定程度上受到抑制,其中低浓度(200 mmol·L1)在盐胁迫20 d后对苗高有显著的抑制作用,而中、高浓度(400、600 mmol·L1)在10 d后就出现显著的抑制作用,且碱性盐处理下抑制作用更大。不同浓度盐胁迫均是在30 d后对地径有显著的抑制作用。不同浓度盐胁迫均和盐分类型对苦楝地上部分和总生物量均有显著的抑制作用,对地下部分影响不显著,碱性盐处理下抑制作用更大。
Bernstein et al.(2004)研究表明植物在遭受胁迫的状态下会将有限的资源分配到特定的器官以维持器官功能的正常发挥来辅助植株降低胁迫对自身的伤害。本研究中盐胁迫导致苦楝根系吸水困难,导致地上生物量受到显著影响,同时苦楝为保证根系正常功能而将有限资源分配到地下部分,保证根系生物量的积累。较高的根系生物量有利于根系对水分和养分的吸收,从而稀释体内的盐分,提高苦楝对胁迫的耐受能力(Silveira et al., 2009;王树凤等,2014)。这与黄稚清等(2019)研究发现一定浓度盐胁迫能够促进红果风铃木根系的生长,但抑制地上部分生长的结果一致。碱性盐胁迫条件下,苦楝需要消耗更多的能量通过脯氨酸来进行渗透调节,又要对升高的pH进行调节,导致对植株生长更大的抑制作用(王保平等,2013)。Lin et al. (2016)研究同样表明碱性盐胁迫下更强的抑制作用可能是由于高pH值的破坏性影响,直接导致离子失衡和代谢紊乱根际。此外,高pH的环境能够通过沉淀Ca2+、Mg2+和HPO-3來降低矿质元素的有效性,从而抑制离子吸收,破坏离子平衡(Yang et al., 2007)。
叶片是植物进行光合作用的主要器官,叶绿素是与植物光合作用有关的最重要的色素。本研究中,在低浓度盐胁迫条件下,苦楝的叶绿素含量与对照差异不显著,一个原因可能是低浓度的盐胁迫刺激苦楝体内的保护机制,促进叶绿素含量的提高(孔强等,2019),另一个原因可能是苦楝吸收较多的钠离子降低叶绿素酶的活性,进而促进叶绿素的积累(武小靖等,2014)。同时也证明苦楝对200 mmol·L1浓度的Na2SO4和Na2C03也有一定抗性。而高浓度盐胁迫下,苦楝的叶绿素含量一直低于对照,主要是因为高浓度盐胁迫会对叶绿体基粒片层和类囊体结构造成不可逆的破坏,造成叶绿素含量的降低(穆永光,2016)。另一种解释是离子毒害降低叶绿素-蛋白质复合物的稳定性,同时增加叶绿素酶活性,促进叶绿素的分解,抑制叶绿素的合成,破坏叶绿体结构(Djanaguiraman & Ramadass,2004;赵秋月和张广臣,2015)。本研究结果发现,盐胁迫显著抑制苦楝幼苗的净光合速率、蒸腾速率、气孔导度和胞间CO2浓度。因为盐胁迫会导致叶绿素生物合成受阻、光合色素降解以及参与光合作用的酶失活,进而造成光合作用紊乱(邹丽娜等,2011;Ashraf & Harris,2013)。盐胁迫刺激植株根系产生的脱落酸以及渗透胁迫导致气孔导度的降低(吴顺等,2014)。气孔导度的降低又阻碍CO2通过气孔进入叶绿体参与光合作用,导致净光合速率的降低,表明气孔限制是苦楝光合速率降低的主要因素(李远航等,2019)。但也有研究表明,高浓度盐胁迫条件下,非气孔限制是光合作用降低的主要因素,但只有当气孔导度下降,同时胞间CO2浓度提高时才能说明是非气孔限制导致光合速率降低(高冠龙等,2018)。本研究结果表明,与中性盐相似,碱性盐胁迫下苦楝的光合指标及叶绿素含量均呈下降趋势,随处理时间增加,下降比例大于中性盐处理。主要是苦楝在碱性盐胁迫下,既受到离子毒害作用,又受到高pH的损害,高pH会破坏植株的核酸、蛋白质等大分子物质,还会对生物膜产生不可逆的损伤,影响其选择透过性,破坏或抑制跨膜电化学电位梯度,从而对植株造成更大的破坏(Yang et al., 2009;郭淑华等,2017)。另外高pH既通过影响叶绿体中类囊体两侧H+浓度降低了叶绿体ATP合成的动力,又通过破坏苦楝叶片细胞内外的酸碱平衡而使植株耗费更多的能量来向细胞外分泌H+或积累具有缓冲作用的代谢产物(郭立泉,2005;刘兆普等,2017)。此外,盐胁迫处理的相对电导率和饱和水分亏缺均显著高于对照,主要是在盐碱条件下,苦楝不能达到最佳含水量,并且钠离子的积累增加,阻碍剩余部分的吸收。且碱性胁迫引起的高pH值强烈抑制根系的吸水,苦楝幼苗合成了大量的渗透调节物质来抵抗胁迫,从而消耗更多的水分(Wang et al., 2019)。盐碱还会对脂质产生氧化损伤,并降低膜的稳定性(Mittal et al., 2012;SienkiewiczCholewa et al., 2018)。总之,盐碱胁迫下苦楝叶绿素含量、蒸腾速率、气孔导度和胞间CO2浓度的降低,导致光合光合速率的降低,光合光合速率的降低和叶面积的减少共同导致植物生长量的减少(Palmerini et al., 2019)。在盐碱地种植苦楝,发展苦楝耐盐品种是提高苦楝耐盐性的有效途径。本研究主要是涉及盐碱胁迫对苦楝幼苗生长和光合生理指标的影响,但盐碱的危害是多方面的,例如盐和碱的积累导致土壤结构的退化,从而影响土壤的水分和空气运动以及根系的伸展(Bano & Fatima, 2009)。本研究表明,苦楝对中性盐和碱性盐胁迫均有一定抗性,碱性盐对苦楝幼苗的胁迫程度更大,揭示盐胁迫下苦楝幼苗的形态方面和光合生理方面的适应机制,能够为抗盐和耐盐苦楝的培育提供理论基础。中性盐和碱性盐胁迫下苦楝的耐盐性机理还有待进一步研究。同时还有必要进行田间试验,以获得更加客观的结果,为苦楝在盐碱地区的推广提供重要依据。
参考文献:
ASHRAF M, HARRIS PJC, 2013. Photosynthesis under stressful environments: An overview [J]. Photosynthetica, 51(2): 163-190.
BANO A, FATIMA M, 2009. Salt tolerance in Zea mays L. following inoculation with Rhizobium and Pseudomonas [J]. Biol Fert Soils, 45(4): 405-413.
BERNSTEIN N, MEIRI A, ZILBERSTAINE M, 2004. Root growth of avocado is more sensitive to salinity than shoot growth [J]. J Amer Soc Hort Sci, 129(2): 188-192.
CHEN LJ, DENG XM, DING MM, et al., 2014. Geographic variation in traits of fruit stones and seeds of Melia azedarach [J]. J Beijing For Univ, 36(1): 15-20.[陈丽君, 邓小梅, 丁美美, 等, 2014. 苦楝种源果核及种子性状地理变异的研究 [J]. 北京林业大学学报, 36(1): 15-20.]
CHENG SM, GU WC, 2005. The phenological division of distribution area in China for Melia azedarach [J]. Sci Silv Sin, 41(3): 186-191.[程诗明, 顾万春, 2005. 苦楝中国分布区的物候区划 [J]. 林业科学, 41(3): 186-191.]
DJANAGUIRAMAN M, RAMADASS R, 2004. Effect of salinity on chlorophyll content of rice genotypes [J]. Agric Sci Dig, 24(3): 178-181.
GAO GL, FENG Q, ZHANG XY, et al., 2018. An overview of stomatal and nonstomatal limitations to photosynthesis of plants [J]. Arid Zone Res, 35(4): 929-937.[高冠龍, 冯起, 张小由, 等, 2018. 植物叶片光合作用的气孔与非气孔限制研究综述 [J]. 干旱区研究, 35(4): 929-937.]
GUO LQ, 2005. Comparison of organic acid accumulation in Puccinellia tenuiflora under salt stress and alkali stress [D]. Changchun: Northeast Normal University.[郭立泉, 2005. 盐、碱胁迫下星星草体内有机酸积累比较 [D]. 长春: 东北师范大学.]
GUO SH, NIU YJ, ZHAI H, et al., 2017. Effect of alkaline salt stress on fluorescence characteristics, xanthophyll cycle and cyanideresistant respiration of grape hybrid rootstocks [J]. Plant Physiol J, 53(11): 88-96.[郭淑华, 牛彦杰, 翟衡, 等, 2017. 碱性盐胁迫对葡萄杂交砧木荧光特性、叶黄素循环与抗氰呼吸的影响 [J]. 植物生理学报, 53(11): 88-96.]
HUANG ZQ, YUAN S, DING SF, et al., 2019. Effects of salt stress on the growth of Handroanthus chrysotrichus seedlings [J]. Mod Hort, 373(1): 25-27.[黄稚清, 袁森, 丁释丰, 等, 2019. 盐分胁迫对红果风铃木幼苗生长的影响 [J]. 现代园艺, 373(1): 25-27.]
JIN ZL, ZHOU QY, 2014. Effects of NaCl stress on seed germination and seedling growth of Zea mays L.[J]. Hubei Agric Sci, 53(13): 2989-2991.[金正律, 周秋艳, 2014. 盐胁迫对不同玉米品种种子发芽及幼苗生长的影响 [J]. 湖北农业科学, 53(13): 2989-2991.]
KONG Q, MA XH, GONG LX, et al., 2019. Growth and physiological responses of Taxodium mucronatum×Cryptomeria fortunei under different salt stress conditions [J]. J SW For Univ, 39(2): 179-183.[孔强, 马晓华, 宫莉霞, 等, 2019. 不同盐胁迫条件下东方杉的生长及生理响应研究 [J]. 西南林业大学学报, 39(2): 179-183.]
LI YZ, YANG JS, YU M, et al., 2019. Different effects of NaCl and Na2SO4 on the carbon mineralization of an estuarine wetland soil [J]. Geoderma, 344: 179-183.
LI YH, HE KN, ZHANG T, et al., 2019. Effects of salt stress on the photosynthetic and physiological indexes of Lycium ruthenicum [J]. Sci Soil Water Conserv, 17(1): 83-88.[李遠航, 贺康宁, 张潭, 等, 2019. 盐胁迫对黑果枸杞光合生理指标的影响 [J]. 中国水土保持科学, 17(1): 83-88.]
LIN JX, WANG YN, SUN SN, et al., 2016. Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under saltalkali stress and nitrogen deposition [J]. Sci Total Environ, 576: 234-241.
LIU ZP, XUE YF, 2008. Effects of NaCl and Na2CO3 stresses on photosynthesis and parameters of chlopophyll fluorescence in Helianthus tuberosus seedlings [J]. J Plant Ecol, 32(1): 161-167.[刘兆普, 薛延丰, 2008. 不同浓度NaCl和Na2CO3处理对菊芋幼苗光合及叶绿素荧光的影响 [J]. 植物生态学报, 32(1): 161-167.]
LIU ZX, ZHANG HX, YANG XY, et al., 2014. Growth, and cationic absorption, transportation and allocation of Elaeagnus angustifolia seedlings under NaCl stress [J]. Acta Ecol Sin, 34(2): 326-336.[刘正祥, 张华新, 杨秀艳, 等, 2014. NaCl 胁迫下沙枣幼苗生长和阳离子吸收, 运输与分配特性 [J]. 生态学报, 34(2): 326-336.]
MIAO HX, SUN MG, XIA Y, et al., 2005. Effects of salt stress on root activity of Melia azedarach L. seedlings [J]. J Shandong Agric Univ (Nat Sci Ed), 36(1): 9-12.[苗海霞, 孙明高, 夏阳, 等, 2005. 盐胁迫对苦楝根系活力的影响 [J]. 山东农业大学学报(自然科学版), 36(1): 9-12.]
MITTAL S, KUMARI N, SHARMA V, 2012. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes [J]. Plant Physiol Biochem, 54: 17-26.
MU YG, 2016. Effect of salinealkali stress on growth and physiology of Amorpha fruticosa [D]. Changchun: Northeast Normal University.[穆永光, 2016. 盐碱胁迫对紫穗槐生长和生理的影响 [D]. 长春: 东北师范大学.]
MUNNS R, 2005. Genes and salt tolerance: bringing them together [J]. NewPhytol, 167(3): 645-663.
OU C, HU X, YAO XM, et al., 2019. Physiological characteristics in Platycladus orientalis under salt and drought stress and regulated by SA [J]. J Arid Land Resour Environ, 33(8): 186-192.[偶春, 胡欣, 姚侠妹, 等, 2019. 盐旱胁迫下侧柏幼苗的生理响应及水杨酸的调控作用 [J]. 干旱区资源与环境, 33(8): 186-192.]
PALMERINI CA, BALDONI L, MARIOTTI R, et al., 2019. Behavior of four olive cultivars during salt stress [J]. Front Plant Sci, 10: 867.
SHAN QH, ZHANG JF, RUAN WJ, et al., 2011. Response of soil quality indicators to comprehensive amelioration measures in coastal saltaffected land [J]. Acta Ecol Sin, 31(20): 6072-6079.[单奇华, 张建锋, 阮伟建, 等, 2011. 滨海盐碱地土壤质量指标对生态改良的响应 [J]. 生态学报, 31(20): 6072-6079.]
SIENKIEWICZCHOLEWA U, SUMISLAWSKA J, SACALA E, et al., 2018. Influence of silicon on spring wheat seedlings under salt stress [J]. Acta Physiol Plant, 40(3): 54.
SILVEIRA JAG, ARAUJO SAM, LIMA JPMS, et al., 2009. Roots and leaves display contrasting osmotic adjustment mechanisms in response to NaClsalinity in Atriplex nummularia [J]. Environ Exp Bot, 66(1): 1-8.
WANG BP, DONG XY, DONG KH,2013. Effect of salinealkali stress on the physiological characteristics of alfafa seedlings [J]. Acta Agr Sin, 21(6): 1124-1129.[王保平, 董曉燕, 董宽虎, 2013. 盐碱胁迫对紫花苜蓿幼苗生理特性的影响 [J]. 草地学报, 21(6): 1124-1129.]
WANG JY, 2013. Studies on salttolerant mechanism of seeds and seedlings in Melia azedarach L.[D]. Nanjing: Nanjing Forestry University.[王家源, 2013. 苦楝种苗耐盐胁迫的生理响应机制研究 [D]. 南京: 南京林业大学.]
WANG SF, HU YX, SUN HJ, et al., 2014. Effects of salt stress on growth and root development of two oak seedlings [J]. Acta Ecol Sin, 34(4): 1021-1029.[王树凤, 胡韵雪, 孙海菁, 等, 2014. 盐胁迫对2种栎树苗期生长和根系生长发育的影响 [J]. 生态学报, 34(4): 1021-1029.]
WANG YN, JIE WG, PENG XY, et al., 2019. Physiological adaptive strategies of oil seed crop Ricinus communis early seedlings (cotyledon vs. true leaf) under salt and alkali stresses: From the growth, photosynthesis and chlorophyll fluorescence [J]. Frontiers Plant Sci, 9: 1939.
WEI HX, SUN MG, XIA Y, et al., 2005. Effects of NaCl stress on the membrane permeability and the content of osmotic adjustable organic substances of Melia azedarach seedlings [J]. J Gansu Agric Univ, 10(5): 599-603.[魏海霞, 孙明高, 夏阳, 等, 2005. NaCl胁迫对苦楝细胞膜透性和有机渗透调节物质含量的影响 [J]. 甘肃农业大学学报, 10(5): 599-603.]
WU S, ZHANG XQ, CAI Y, 2014. Effects of drought stress on chlorophyll contents and photosynthetic characteristics of cucumber seedlings [J]. Chin Agric Sci Bull, 30(1): 133-137.[吴顺, 张雪芹, 蔡燕, 2014. 干旱胁迫对黄瓜幼苗叶绿素含量和光合特性的影响 [J]. 中国农学通报, 30(1): 133-137.]
WU XJ, YANG Q, ZHANG GJ, et al., 2014. Effect of salinity of soil on physiobiochemistry indexes of Lycium barbarum [J]. N Hortic, (12): 137-141.[武小靖, 杨晴, 张国君, 等, 2014. 土壤含盐量对枸杞生理生化指标的影响 [J]. 北方园艺, (12): 137-141.]
XU LP, LIU J, ZHANG ZH, et al., 2018. Effect of salt stress on growth and physiology in Melia azedarach seedlings of six provenances [J]. Int J Agric Biol, 20(2): 471-480.
YANG CW, CHONG JN, LI CY, et al., 2018. Osmotic adjustment and ion balance traits of an alkaline resistant halophyte Kochia sieversiana during adaptation to saline and alkaline conditions [J]. Plant Soil, 294: 263-276.
YANG CW, SHI DC, WANG DL, 2008. Comparative effects of salt and alkali stresses on growth, osmotic adjustment and ionic balance of an alkaliresistant halophyte Suaeda glauca (Bge.) [J]. Plant Growth Regul, 56(2): 179-190.
YANG CW, ZHANG ML, LIU J, et al., 2009. Effects of buffer capacity on growth, photosynthesis, and solute accumulation of a glycophyte (wheat) and a halophyte (Chloris virgata) [J]. Photosynthetica, 47(1): 55-60.
ZHANG ZH, WANG JY, YU FY, 2014. Effects of NaCl stress on germination characteristics of Melia azedarach L. seeds from two provenances [J]. J Nanjing For Univ (Nat Sci Ed), 38(5): 107-112.[張子晗, 王家源, 喻方圆, 2014. 盐胁迫对两种源苦楝种子萌发特性的影响 [J]. 南京林业大学学报(自然科学版), 38(5): 107-112.]
ZHAO QY, ZHANG GC, 2015. Effects of alkaline salt stress on root activity and photosynthetic pigment of three tomato species [J]. Jiangsu Agric Sci, 43(11): 219-223.[赵秋月, 张广臣, 2015. 碱性盐胁迫对3种番茄根系活力和光合色素的影响 [J]. 江苏农业科学, 43(11): 219-223.]
ZOU LN, ZHOU ZY, YAN SY, et al., 2011. Effect of salt stress on physiological and biochemical characteristics of Amorpha fruticosa seedlings [J]. Acta Pratac Sin, 20(3): 84-90.[邹丽娜, 周志宇, 颜淑云, 等, 2011. 盐分胁迫对紫穗槐幼苗生理生化特性的影响[J]. 草业学报, 20(3): 84-90.]
(责任编辑周翠鸣)
猜你喜欢
睿士(2020年11期)2020-11-16
中学生天地(A版)(2020年3期)2020-04-10
故事作文·高年级(2020年3期)2020-03-17
小资CHIC!ELEGANCE(2019年33期)2019-11-22
小资CHIC!ELEGANCE(2019年15期)2019-05-21
财经(2017年6期)2017-03-29
红领巾·萌芽(2016年1期)2016-09-10
课堂内外(高中版)(2016年5期)2016-07-11
课堂内外(小学版)(2016年4期)2016-07-04
英语学习·新锐空间(2013年3期)2013-05-23
