
西门塔尔牛与我国地方黄牛的杂种优势预测分析
2021-03-30 09:20范婷婷张路培徐凌洋高会江李俊雅
畜牧兽医学报 2021年3期
范婷婷,陈 燕,张路培,徐凌洋,高会江,李俊雅,高 雪
(中国农业科学院北京畜牧兽医研究所,北京100193)
我国肉牛品种主要为地方黄牛品种。黄牛品种作为我国的特色资源,具有耐粗饲、抗逆性好、肉质细嫩等优点,但因其役用的历史缘故,同时也存在生长速度缓慢,体型发育不佳、胴体产肉少、优质牛肉切块率低等缺陷[1]。为了增加产肉量,提高牛肉品质,我国引入了大量优良的外国肉牛品种,对本土黄牛品种进行杂交改良,培育出了很多优秀的肉用专门化品种,例如利用夏洛莱培育了辽育白牛和夏南牛。我国自20世纪50年代开始引进西门塔尔牛,利用西门塔尔牛与本地牛进行杂交后选育获得了生产性能良好的中国西门塔尔牛[2]和蜀宣花牛[3]两个乳肉兼用新品种,而且两个培育的新品种在我国的适应性较好。西门塔尔牛的杂交后代占我国肉牛饲养产业的80%以上,是我国肉牛生产及改良所用的主导品种。杂种优势利用成为了提高地方黄牛生产性能的重要途径之一。杂种优势利用的一般方法是在杂交试验的基础上进行不同亲本种群之间的杂交,然后对杂种后代进行评比鉴定,最终筛选出最佳的杂交组合进而应用于生产。这个过程不仅耗费大量的人力、物力和时间[4],而且畜禽的种群又很多,不可能全部两两进行杂交试验,同时大量的杂交试验也带来了育种结果的不确定性。因此,在杂交试验之前对亲本组合潜在的杂种优势进行快速、准确的预测,对最终获得符合生产目标的杂交种,加快育种进程,提高育种效率具有重要的意义。
随着现代分子生物学和分子遗传学的发展,使利用分子水平的遗传标记预测畜禽杂种优势和估计畜禽基因组杂种优势及其组成成分[5-7]成为了可能[8]。目前应用最广泛的是通过微卫星标记预测杂种优势,但是其研究结果却有很大的不一致性[9-11]。而SNP标记以其存在的优势开始被研究者发现,并发掘其在杂种优势预测中的潜在价值。Amuzu-Aweh 等[12]利用全基因组SNPs研究白来航鸡纯系品种的杂交数据,对杂种优势进行预测,发现杂种优势与亲本品系之间等位基因频率之差的平方(SDAF)成正比,预测的杂种优势准确率达到了50%。徐忠等[13]利用全基因组与性状相关的SNPs分子标记估算不同群体间的遗传距离,对金华猪不同杂交模式下的杂种优势进行预测,筛选不同杂交模式下较优的杂交组合。虽然SNP标记得到了广泛的应用,但是关于SNP标记在牛杂种优势预测上的研究报道并不多。
本研究以西门塔尔牛及地方黄牛作为亲本构建杂交群体,并利用770K高密度芯片进行基因分型,分别用全基因组和与性状相关的SNPs数据估算亲本间的遗传距离,预测亲本各杂交组合生长发育及胴体性状的杂种优势, 以筛选出较优的杂交组合,为地方黄牛的杂交改良提供一定的理论参考依据。
1 材料与方法
1.1 试验材料
自内蒙古锡林郭勒盟乌拉盖管理区牧场收集1 222头 西门塔尔牛(Chinese Simmental beef cattle,CSC)群体的数据,此西门塔尔牛群体是将国外引进西门塔尔牛与本地北方黄牛进行杂交后选育获得的,经过十几年的选种选育已经逐渐偏向肉用。8个地方黄牛品种共190头,采集血样,提取基因组DNA,获得基因型数据。地方黄牛品种的具体信息见表1。其中蒙古牛、西藏牛和柴达木牛为北方黄牛品种,其余地方黄牛为南方黄牛品种。选择的目标性状为生长发育、胴体和肉质性状,包括初生重(birth weight,BW)、胴体重(carcass weight,CW)、周岁重(yearling weight,YW)和大理石花纹评分(marbling score,MS)性状。
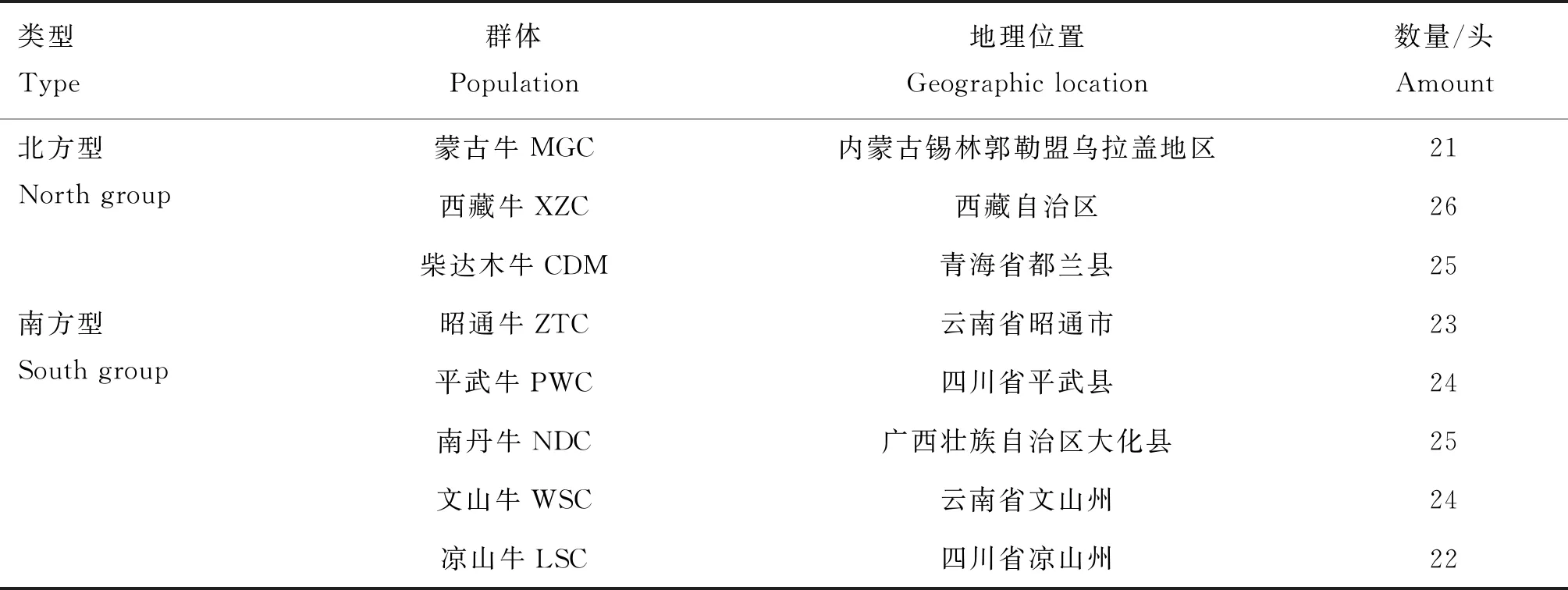
表1 地方黄牛各品种信息
1.2 血液样本基因分型和质量控制
使用llumina BovineHD(770K)芯片对1 222头西门塔尔牛群体和190头地方黄牛个体进行基因分型,在分析之前,利用PLINK v1.90软件[14]对基因型数据进行质量控制,剔除不合格的个体和SNPs,质控标准:1)SNPs检出率>95%;2)SNP的最小等位基因频率>1%;3)哈代-温伯格平衡检验(P>10-6);4)个体SNP缺失率<0.1;5)SNP位于常染色体。质控后,地方黄牛190个个体和622 162个SNPs用于分析,西门塔尔牛1 217个个体和675 029个 SNPs用于分析。接着利用BEAGLE v3.3.2软件对缺失位点的基因型进行填充。
1.3 性状相关SNP位点筛选
将牛QTL数据库(https://www.animalgenome.org/cgi-bin/QTLdb/BT/index)中关于生长、胴体和肉质性状的QTLs筛选出来。通过Perl软件将目的性状QTL与牛参考基因组进行比对,分别获得包含基因的性状QTL,从而筛选出与胴体和肉质、生长性状相关的QTL集合。然后通过R软件将检测到的SNPs位点映射到这些包含基因的性状QTL中,以找到落入QTL覆盖区间内的标记SNPs。
1.4 群体遗传结构分析
为避免互相连锁的SNPs对群体结构分析的影响,选取经PLINK修剪后相互之间连锁不平衡系数 (r2)小于0.2的全基因组SNPs及与性状相关的SNP进行主成分分析(principal component analysis,PCA)(参数:indep-pairwise 50 5 0.2),分析杂交组合两个群体之间的遗传背景联系。通过PLINK软件 (参数:--pca) 进行主成分分析,输出结果用R软件画图展现。
1.5 群体间遗传距离
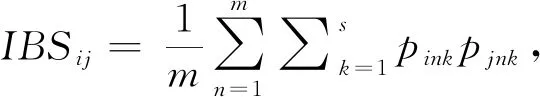
2 结 果
2.1 性状相关的SNP位点分析
通过将西门塔尔牛和地方黄牛的SNPs位点与牛QTL数据库中关于生长、胴体和肉质性状的QTL进行映射,找到与牛大理石花纹评分、胴体重、初生重和周岁重性状潜在相关联的SNPs。西门塔尔牛和地方黄牛与性状相关的SNP数见表2。
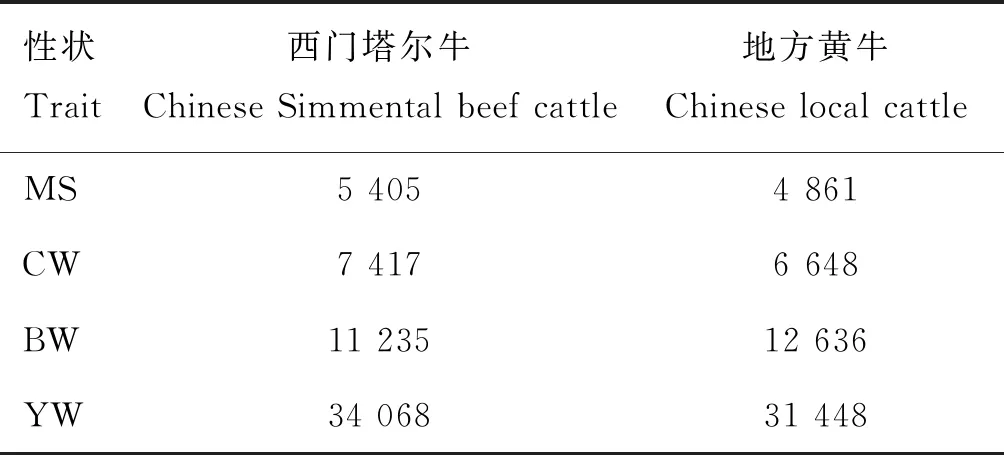
表2 与性状相关的SNP数
2.2 群体的主成分分析
通过全基因组SNPs和与性状相关的SNPs标记对西门塔尔牛和地方黄牛品种的9个杂交组合进行主成分分析,结果如图1、图2所示。从全基因组SNP主成分分析的结果(图1)可以看出,PC1能够解释57.43%的遗传变异,PC2能够解释20.86%的遗传变异。结合PC1和PC2,发现西门塔尔牛群体聚集在一起,南方地方黄牛品种聚集在一起,北方地方黄牛品种聚集在一起。根据PC1可以明显的将南方黄牛品种(南丹牛、文山牛、凉山牛、平武牛、昭通牛)从所有群体中分开,而凉山牛、平武牛、昭通牛相对聚集在一起,分化程度较低,亲缘关系相较于南丹牛和文山牛要近;西门塔尔牛群体与北方黄牛品种(蒙古牛、西藏牛、柴达木牛)相距比较近,表明西门塔尔牛与北方黄牛的亲缘关系相对于南方黄牛较近,遗传背景差异较小。从与性状相关SNPs主成分分析的结果(图2)可以看出,其结果与用全基因SNPs进行主成分分析的结果大致相同。
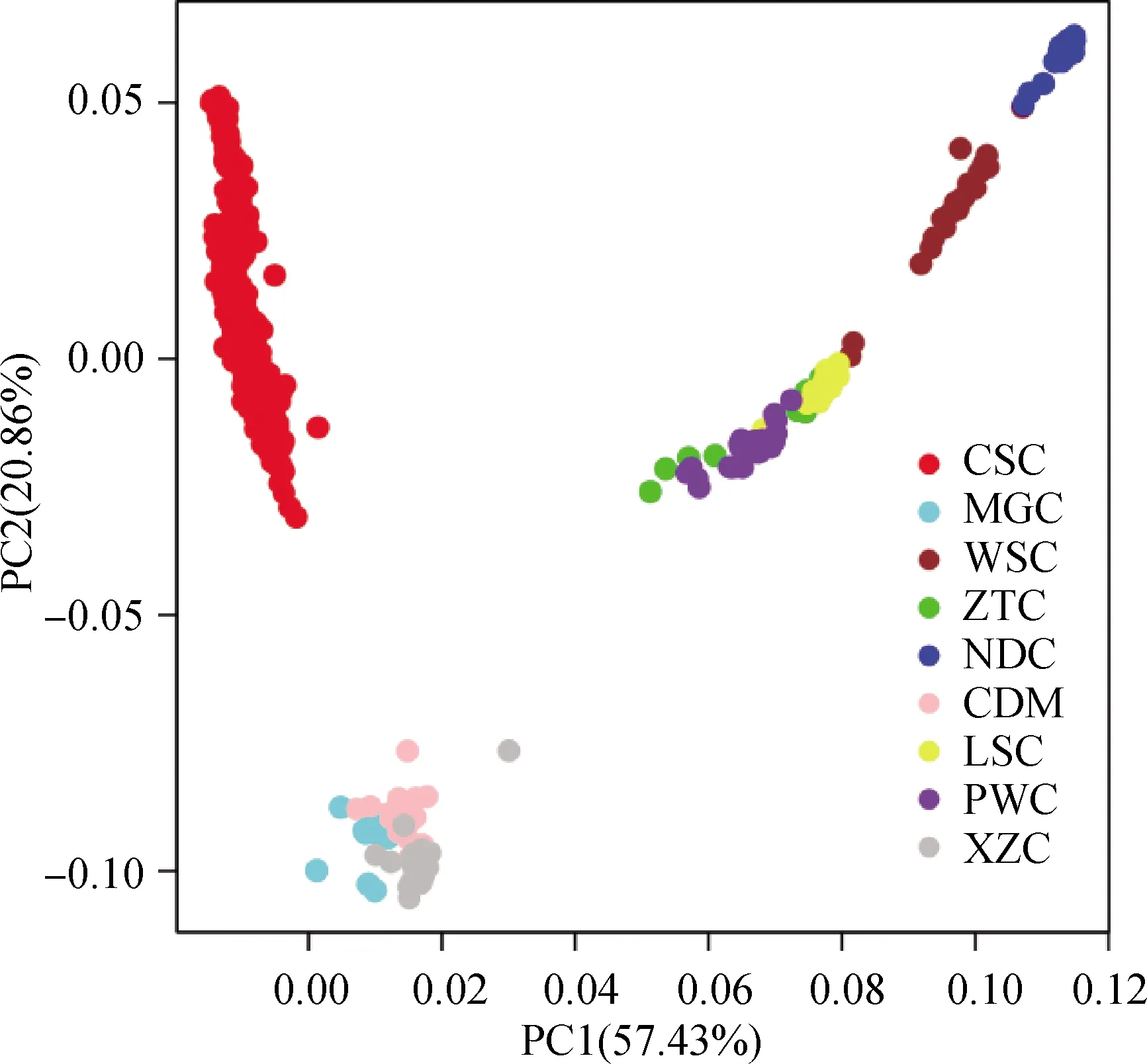
图1 根据全基因组SNPs对西门塔尔牛和地方黄牛群体进行主成分分析Fig.1 Principal component analysis of Chinese Simmental beef cattle and Chinese local cattle populations based on genome-wide SNPs
2.3 各杂交组合间遗传距离比较
由全基因组SNPs及与4个性状具有潜在关联的SNPs计算的亲本间的遗传距离见表3、图3,结果显示,在全基因组水平上,9个亲本间的遗传距离范围为0.306 6~0.323 7,对于初生重性状,9个亲本间的遗传距离变幅为0.301 4~0.323 0,在胴体重性状上,9个 亲本间的遗传距离变幅为0.297 4~0.316 2,对于大理石花纹评分性状,9个亲本间的遗传距离变幅为0.297 2~0.312 5,在周岁重性状上,9个亲本间的遗传距离变幅为0.305 3~0.322 5。在各性状中,遗传距离最大的亲本组合都是CSC与NDC。除了大理石花纹评分性状是CSC与ZTC之间的遗传距离最小,其余性状遗传距离最小的亲本组合都是CSC与PWC。因此,从总体水平来看,在各性状上西门塔尔牛与南丹牛杂交获得的杂种优势可能最大。以上结果与PCA分析的结果相似性很高。
西门塔尔牛与不同地域地方黄牛的之间的遗传距离见图4、图5。西门塔尔牛与北方黄牛组成的杂交组合中(图4),在初生重和胴体重性状上,遗传距离排序为CSC×CDM > CSC×MGC > CSC×XZC;在大理石花纹评分性状上,遗传距离排序为CSC×MGC > CSC×CDM > CSC×XZC;在周岁重性状上,遗传距离排序为CSC×CDM > CSC×XZC > CSC×MGC。西门塔尔牛与南方黄牛组成的杂交组合中(图5),在初生重、胴体重上,遗传距离排序为CSC× NDC > CSC× WSC > CSC× LSC > CSC×ZTC> CSC×PWC;在大理石花纹评分性状上,遗传距离排序为CSC×NDC > CSC×WSC > CSC×PWC > CSC×LSC > CSC×ZTC;在周岁重性状上,遗传距离排序为CSC×NDC > CSC×WSC > CSC×ZTC > CSC×LSC > CSC×PWC。
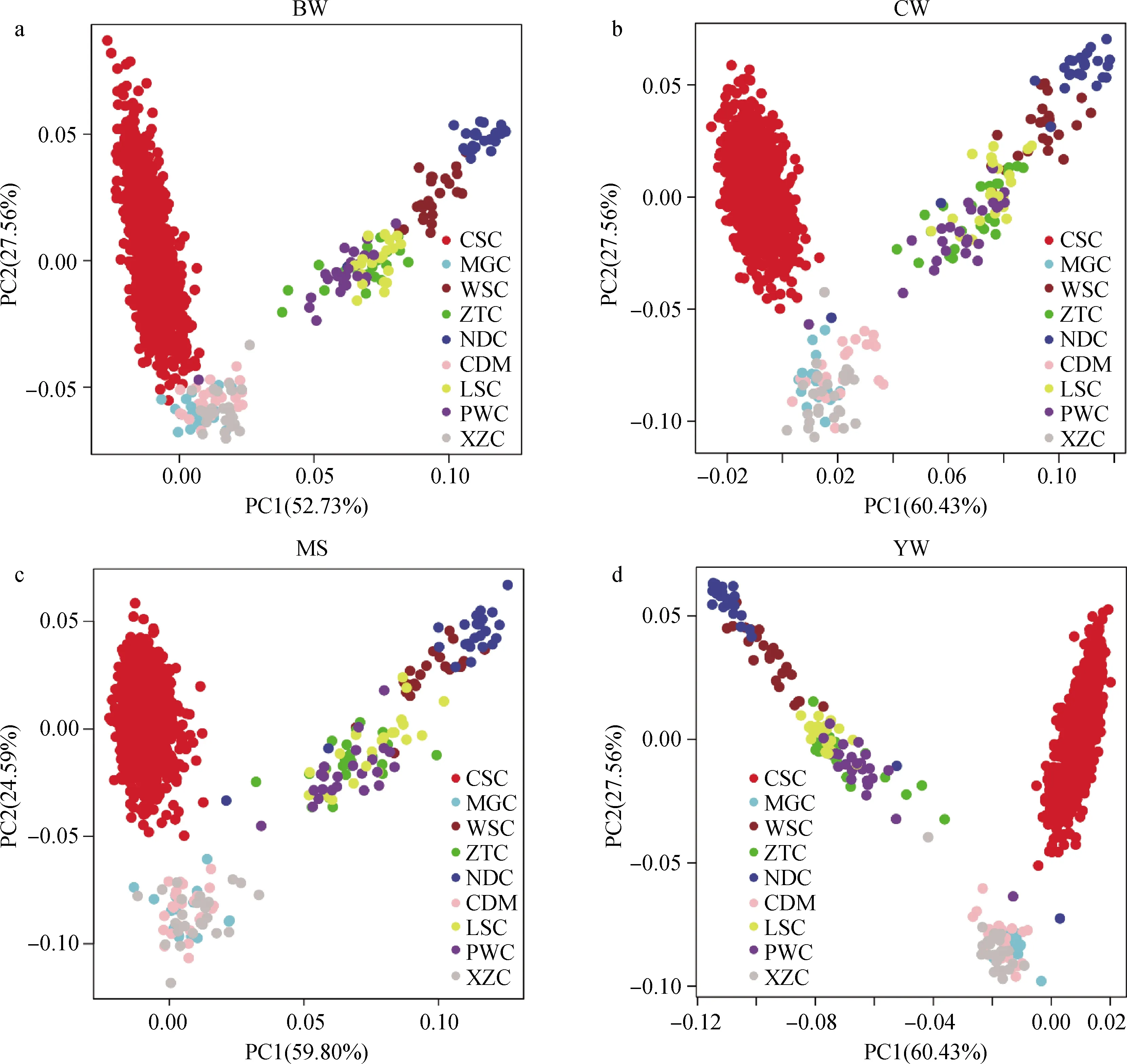
a.初生重的PCA;b.胴体重的PCA;c.大理石花纹评分的PCA;d.周岁重的PCA a. PCA of birth weight; b. PCA of carcass weight; c. PCA of marbling score; d. PCA of yearling weight图2 根据与性状相关的SNPs对西门塔尔牛和地方黄牛群体进行主成分分析Fig.2 Principal component analysis of Chinese Simmental beef cattle and Chinese local cattle populations based on trait-specific SNPs
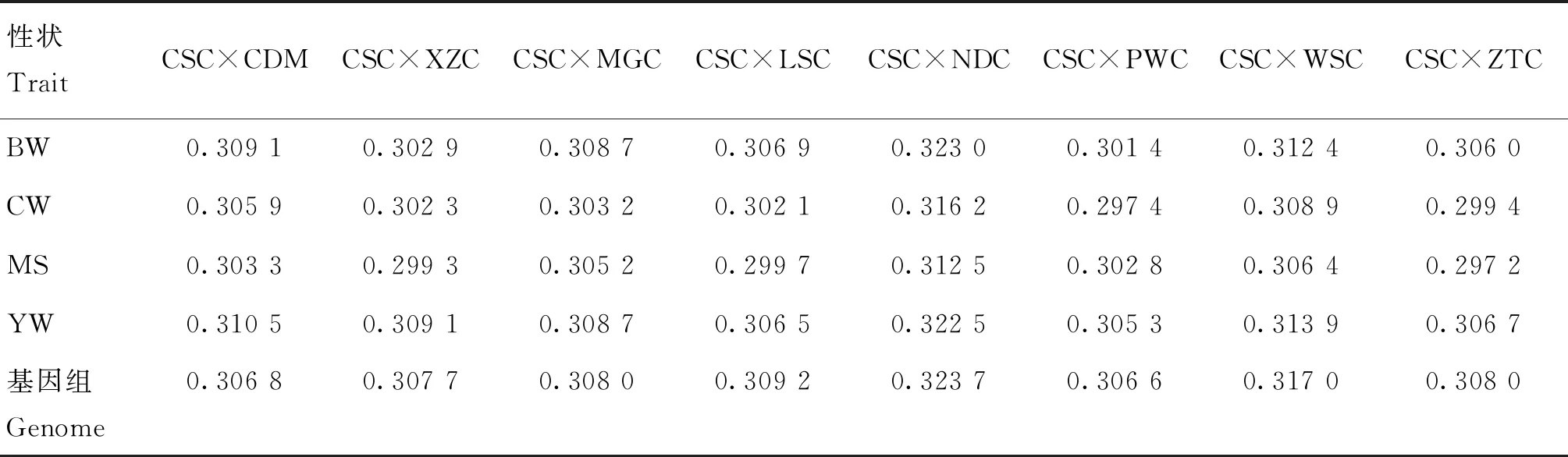
表3 9个亲本间SNPs估算的遗传距离
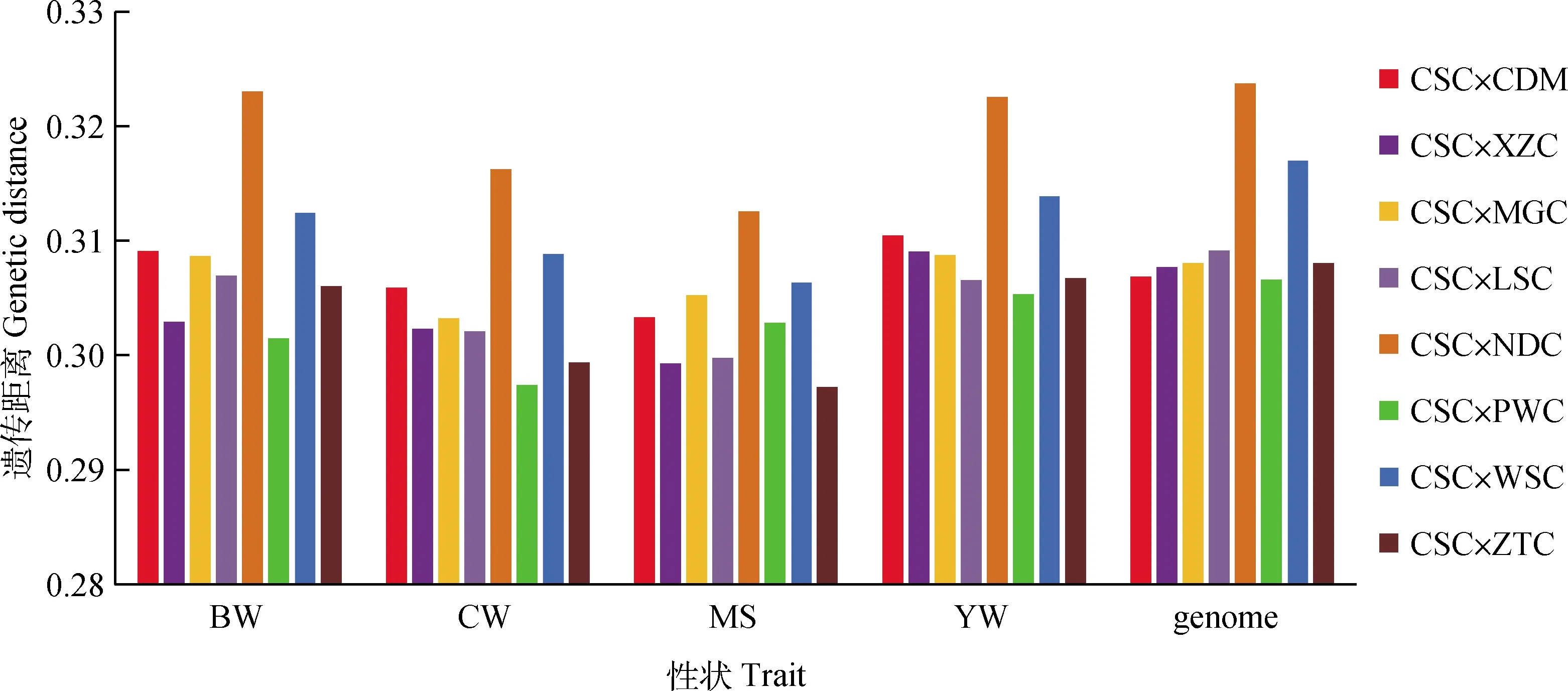
图3 8个杂交组合间的遗传距离Fig.3 Genetic distance among 8 hybrid combinations
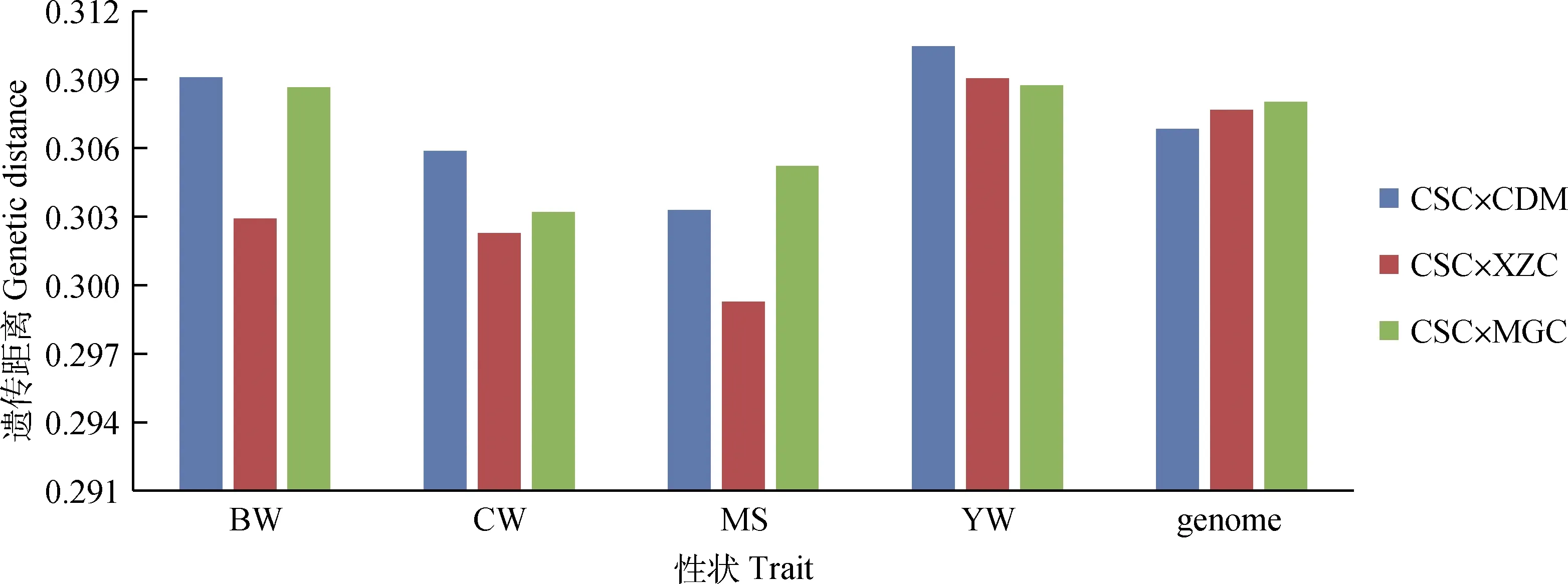
图4 西门塔尔牛与北方黄牛之间的遗传距离Fig.4 Genetic distance between Chinese Simmental beef cattle and Northern breeds of Chinese local cattle
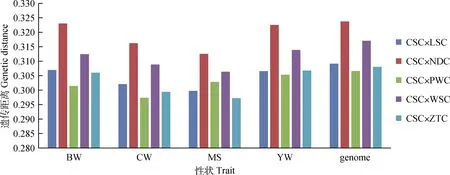
图5 西门塔尔牛与南方黄牛之间的遗传距离Fig.5 Genetic distance between Chinese Simmental beef cattle and Southern breeds of Chinese local cattle
3 讨 论
3.1 分子标记多态性及群体结构分析
随着高通量测序技术的发展,SNP标记作为新一代的分子标记类型,具有密度高、分布范围广、分型简单等优点,在分子遗传研究中逐渐兴起[15]。尤其是快速、高通量的SNP分型技术即SNP芯片的问世,让SNP标记在杂种畜禽的遗传多样性分析[16-17]和关联分析[18-20]等研究中得到广泛应用。本研究采用牛770K SNP芯片对两个亲本群体进行全基因组基因型分型,筛选后得到地方黄牛622 162个有效多态性SNPs位点,西门塔尔牛675 029个有效多态性SNPs位点,可见SNP标记多态性的丰富性和SNP芯片检测方法的高通量特性。
群体结构分析主要是研究遗传变异导致各群体产生分化的一种动态变化规律,能够根据不同品种的分化走向确定品种所属类型。从PCA分析结果可以发现,两个亲本群体中,西门塔尔牛、北方黄牛和南方黄牛基本能保持地域特异性,3个不同地域的牛品种独立形成聚类。西门塔尔牛与北方黄牛品种在PCA图上的距离相对于南方黄牛品种较近,这与李荣岭等[21]采用Nei′s遗传距离(DA)进行NJ聚类分析中,外来品种西门塔尔牛与北方黄牛聚为一类结果相似,说明西门塔尔牛与北方黄牛亲缘关系相对较近,遗传背景差异较小。本研究所用的西门塔尔牛为引进的西门塔尔牛与北方黄牛进行杂交培育而成的,所以与北方黄牛品种的亲缘关系近。西门塔尔牛与北方黄牛品种杂交产生的杂种优势不如与南方黄牛品种杂交形成的杂种优势大。西门塔尔牛与南丹牛遗传关系最远,在所有杂交组合中,CSC与NDC杂交可能会产生最大的杂种优势。
3.2 杂种优势预测分析
在基因序列水平或基因频率水平上计算的不同个体、种群或物种之间的遗传差异的任何数值度量,皆可定义为遗传距离[22]。状态一致(IBS)是指两个或两个以上个体的DNA片段具有相同的核苷酸序列。因此亲本之间的IBS距离应该与后代杂种优势呈负相关。数量遗传学理论认为,杂种优势与遗传距离之间具有一定的线性关系。通常认为,亲本间遗传距离越远、品系纯度越高,后代表现出来的杂种优势就会越明显[23]。吴常信[24]指出,用DNA多态性测定品种或品系间的差异,并据此作出遗传距离要比根据其它材料稳定,因此用来预测杂种优势也更为准确。曹阳等[25]利用微卫星标记分析了4个引进肉牛品种与西门塔尔杂交牛之间的遗传距离,并预测杂种优势,发现西杂牛与4个引进肉牛品种之间的遗传距离为:德国黄牛(1.259 8)、利木赞牛(1.054 6)、红安格斯牛(0.886 7)、夏洛莱牛(0.599 7)。进而推测西杂牛与其他品种牛之间的杂种优势依次为德国黄牛>利木赞牛>红安格斯牛>夏洛莱牛。在其他畜禽,如猪[13,26-27]、鸭[28]、鸡[29]、羊[30-31]等同样依据遗传距离得到了评估杂交组合杂种优势优劣的结果。本研究分别用全基因组SNPs标记及与性状相关的SNPs标记来构建状态同源矩阵,计算各杂交组合亲本间的遗传距离,预测不同杂交组合在生长、胴体和肉质性状上的杂种优势。在全基因组水平推测杂种优势较优的组合可能是CSC与NDC。虽然与性状相关的SNPs标记估算的遗传距离普遍低于全基因组SNPs标记估算的遗传距离,但是在各性状水平杂种优势较优的可能的组合依然是CSC与NDC。与性状相关的分子标记虽然减少了计算中使用标记的数量,但是预测的最优杂交组合和全基因组SNP标记预测结果相同,说明使用与性状相关的分子标记提高了估计的准确性[26]。杂种优势预测可以从不同杂交组合中筛选出最优方案,大大减少了候选的杂交试验,但是此预测方法的可靠性仍然需要进行杂交试验验证。总之, 这一结果可为地方黄牛杂交改良研究提供一定的理论参考依据。
4 结 论
本研究利用覆盖全基因组和与性状相关的SNPs标记估算西门塔尔牛和地方黄牛两个亲本群体之间的遗传距离进而进行杂种优势预测,推测西门塔尔牛与南丹牛可能为最佳的杂交组合。在与北方黄牛的杂种优势预测中,在初生重、周岁重和胴体重性状上,推测西门塔尔牛与柴达木牛可能为最佳的杂交组合;在大理石花纹评分性状上,推测西门塔尔牛与蒙古牛可能为最佳的杂交组合。在与南方黄牛的杂种优势预测中,推测在各个性状中西门塔尔牛与南丹牛可能为最佳的杂交组合。
猜你喜欢
地理教育(2023年1期)2023-02-09
中国动物保健(2022年10期)2022-10-14
蔬菜(2021年7期)2021-11-27
矿产勘查(2020年8期)2020-12-25
长江文艺(2019年5期)2019-08-07
畜牧兽医科学(2018年2期)2018-02-14
中国草食动物科学(2016年2期)2016-11-30
种子科技(2014年8期)2014-05-18
茶叶通讯(2014年2期)2014-02-27
种子科技(2014年9期)2014-01-22
