
3 种雁艾美尔球虫ITS 序列测定与系统发育分析
2021-03-22 07:19李清月龚治忠
阜阳师范大学学报(自然科学版) 2021年1期
李清月,龚治忠,刘 刚
(安徽医科大学 生命科学学院,安徽 合肥230032)
鸟类肠道寄生虫是最常见、对其威胁最为明显的病原体之一。研究表明,寄生虫不仅可以感染鸟类,而且还可以感染哺乳动物甚至人类,具有重要的公共卫生意义[1]。几乎所有的野鸟和家养鸟类都能感染球虫病,球虫病的病原体主要是艾美尔球虫属(Eimeria)[2-3]。艾美尔球虫是一种专性细胞内原生寄生虫,主要寄生在鸟类的小肠壁,生命周期较复杂,可以通过“粪-口”传播方式在动物间传播[4-5]。人类感染艾美尔球虫早期症状不明显,严重时会出现胃肠道不适或者慢性腹泻、厌食、倦怠等症状;有时可引起发热、持续性或者脂肪性腹泻等急性感染症状,甚至会导致死亡[6]。
传统的形态学方法鉴定寄生虫存在一定的局限性,尤其是某些相似或近缘物种较为困难;此外,在镜检时球虫的卵囊必须从粪便中分离出来且进行孢子的孵育,处理过程十分繁琐,不适合规模化检测[7]。目前,关于艾美球虫属物种的分子鉴定及其系统发育关系研究相关工作还比较有限且存在争议,部分学者推测艾美尔球虫属具有单起源特性[8-10],也有学者认为艾美尔球虫可能存在多种起源[11]。如,18S rDNA 基因构建的兔球虫进化树显示,外残体可以反映球虫系统发生关系,其他形态学特征对球虫进化关系的确定影响不明显[12];有研究人员发现地域的差异性对于印度鸡球虫的系统发生关系影响不大[6];国内学者研究发现寄生于肠道的兔艾美尔球虫亲缘关系近,而寄生于肝脏的球虫则形成单独的进化枝[13]。目前,国内外对于迁徙鸟类艾美尔球虫物种鉴定和分类还存在一些争议,系统发生关系尚不明确。
本课题组专注于长江中下游越冬白额雁(Anser albifrons)肠道寄生虫流行病学研究,前期研究发现在白额雁越冬地湿地的周边有大量的家禽养殖场,常出现饲养家禽和白额雁在相同水域混群觅食的现象,这种混群行为增加了寄生虫相互传播的机会(尚未发表),大量研究表明,候鸟和饲养家禽在寄生虫的传播中起着重要的作用,造成寄生虫疾病在家禽、候鸟之间周期性的暴发和循环感染[14-16],因此候鸟携带的肠道寄生虫可能带来严重的公共卫生风险。由于传统分类方法对于寄生虫的物种鉴定还存在一定难度,因此,迁徙鸟类的肠道寄生虫鉴定需要更迅速、准确和效率更高的鉴定方法,大量研究表明ITS(internal transcribed spacer)基因序列具有较高的稳定性和种间特异性[17-19],可以作为研究属、种分类阶元的理想遗传标记[20-21]。设计ITS 基因特异性引物,不仅用来鉴定寄生虫的种类,而且可以研究寄生虫物种间的系统发育关系,ITS 基因序列正在成为寄生虫学研究的主要标记基因[20-21]。本研究拟通过分子生物学方法,鉴定升金湖和菜子湖越冬白额雁肠道内携带的艾美尔球虫种类,并阐明艾美尔球虫属部分物种间的系统发育关系。
1 材料与方法
1.1 样品采集
于2018 年11 月,采取无创采样技术采集来自安徽省升金湖王坝村(30°16'58.89''N,117°0'14.64''E)和菜子湖罗岭镇(30°75'00.11.44''N,117°08'64.25.78''E)越冬白额雁的粪便样本。在采集粪便样品前,用双筒或单筒望远镜观察觅食雁群至少1 h,确定周围没有其他鸟群活动,或与其他集群有明显的障碍物(如田埂、河沟等)间隔后方能采集,根据白额雁的足迹或觅食坑,确定为觅食地点。戴一次性PE 手套仔细辨别并采集新鲜粪便样本,每采集一份更换一次手套,去除接触地面的部分后,将粪便样本放置于50 mL 无菌离心管中,贴上标签,放于冰盒中运回实验室,根据采集地来源及采集时间整理分装,短期内不能检测的样本放置于-20 ℃冰箱保存。
1.2 DNA 提取、PCR 扩增及球虫鉴定
采集的白额雁粪便样品共700 份左右,根据测序结果选出具有代表性的3 种艾美尔球虫序列:升金湖S49 样品,升金湖S5 样品,以及菜子湖C70 样品。为保证环境对寄生虫的影响,样品均为越冬前期所采集的白额雁粪便。采用来自TIANamp Stool DNA 试剂盒(Tiangen Biotech CO,LTD)从粪便样品中提取总DNA,根据文献报道的艾美尔球虫属ITS 基因序列设计引物,上游引物:5’-AAG TTG CGT AAA TAG AGC CCT C-3’,下游引物:5’-CAA GAC ATC CAT TGC TGA AAG T-3’。PCR 扩增体系:10×PCR Buffer 5 μL,MgCl2(25 mmol/L)4 μL,dNTP mixture(2.5 mmol/L)各4 μL,引物各(10 μmol/L)1 μL,rTaq DNA 聚合酶0.5 U,模板DNA 2 μL,灭菌双蒸水32.5 μL,总体积50 μL。反应条件:95 ℃预变性10 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,35 个循环后,72 ℃终延伸10 min。PCR 产物经纯化后送通用生物系统有限公司(安徽)进行双向测序。对获得的ITS 基因序列进行BLAST 搜索,参考置信区间为100%的序列,推断3 种球虫的物种。
1.3 系统发育分析
下载NCBI 上已释放的艾美尔球虫属ITS 基因序列如表1。

表1 22 种艾美尔球虫ITS 序列的GenBank 登录号
以Isospora anthochaerae(GenBank 登录号为KF766051)为外群,用CLUSTALX 2.1 软件对序列进行比对,同时用DNAStar 软件对不同序列进行相似性分析。用Mega 5.0 软件对比对后的序列进行修剪并转换成.nex 格式备用。用Mega 5.0软件对形成的.nex 格式文件进行数据模型选择,最终选取AIC 模式下的模型,将其应用于系统树的构建。最大似然(maximum likelihood,ML)法构建的系统树采用PAUP 4.10 软件,建树前先使用Modeltest v 3.7 选择最优模型,然后将所得最优模型写入PAUP 4.10 软件命令行中,进行最大似然法(ML 树)建树,建树执行快速支持度分析并在一次单独的程序运行中搜索最佳分值的ML 树的方法,自举检验100 次。贝叶斯推论(Bayesian inference,BI)法构建的系统树(用MrBayes 3.1.2 软件建立,建树前使用Modeltest v3.7 选择最优模型,运行4 个马尔可夫链,运行100 万代,直至标准误差小于0.01,每2 500 代抽样一次,舍弃老化样本构建一致树。
2 结果
参考GenBank 上释放的艾美尔球虫属各球虫置信区间最高的ITS 序列,推断出3 种艾美尔球虫分离株的物种:菜子湖C70 株为椭圆艾美尔球虫(E.alabamensis),升金湖S49 株为柔嫩艾美尔球虫(E.tenlla),升金湖S5 株为和缓艾美尔球虫(E. mitis)。三条序列上传至Genbank,登录号为MN598760~MN598762。椭圆艾美尔球虫(E.alabamensis)扩增出ITS 基因片段长度为419 bp,柔嫩艾美尔球虫(E. tenlla)ITS 基因片段长度为271 bp,和缓艾美尔球虫(E.mitis)ITS 基因片段长度为344 bp。另外,E. alabamensis 碱基AT 含量为64.0%,GC 含量为36.0%,E.tenlla 碱基AT 含量为60.2%,碱基GC 含量为39.8%,E.mitis 碱基AT 含量为55.3%,GC 含量为44.7%,本研究中22种艾美尔球虫分离株碱基AT 含量最高为67.8%,碱基GC 含量最高为53.8%,碱基AT 平均含量(58.4%)略高于GC 平均含量(41.5%)如表2。
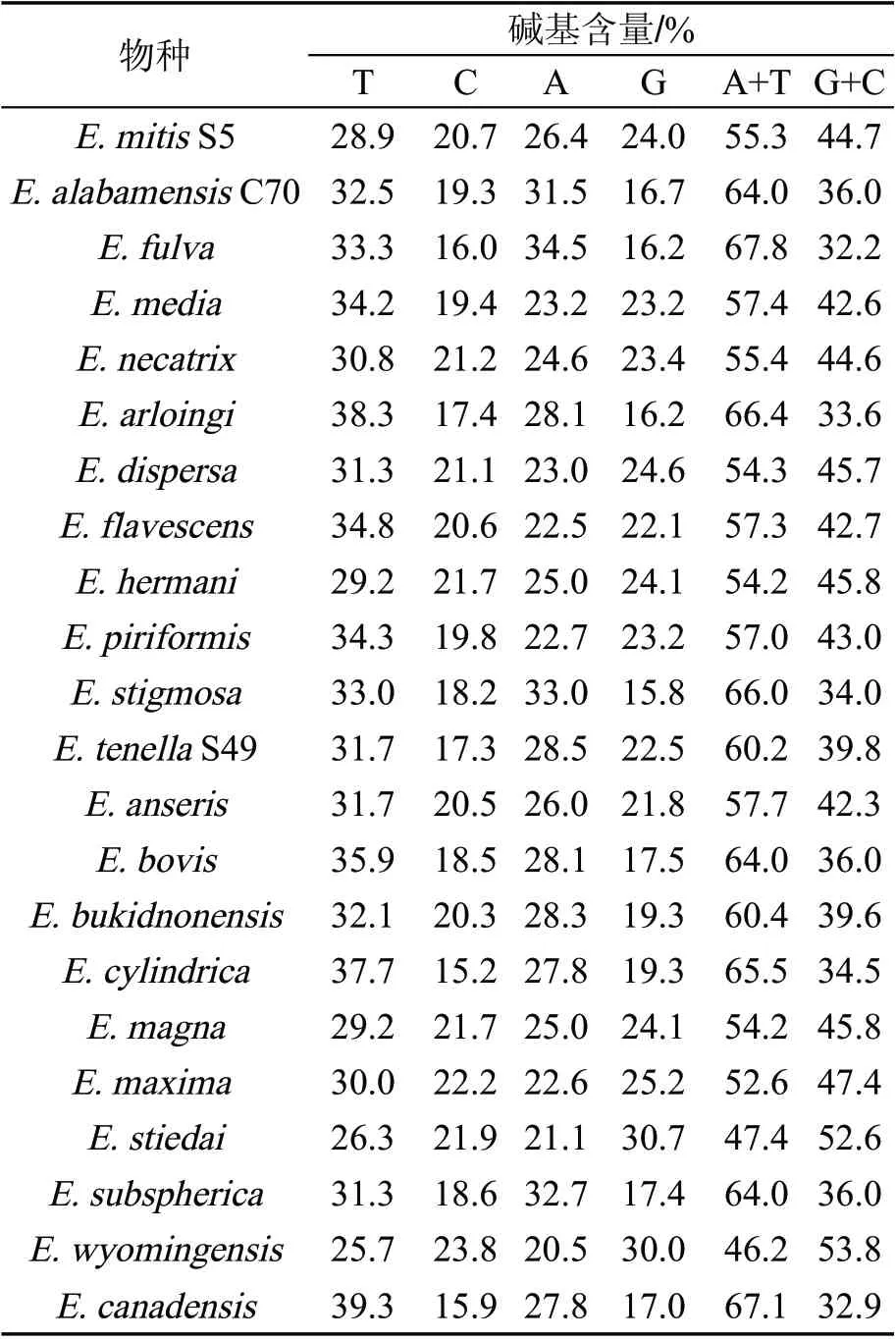
表2 22 种艾美尔球虫分离株ITS 序列的碱基含量
3 种艾美尔球虫ITS 序列中碱基A+T 含量为59.83%,G+C 含量为40.16%,与大多数艾美球虫ITS 序列碱基组成含量相似如表2。
遗传距离分析表明,E. alabamensis 菜子湖C70 株与Genbank 中的家鹅分离出的E.stigmosa和博金斯牛E. subspherica 基因型(登录号AB769806)遗传距离为0.15,遗传差异较小,亲缘关系最近,E. tenlla 升金湖S49 株与Genbank 中的穴兔分离出的E.magna 遗传距离0.85,亲缘关系相对较远,E.mitis 升金湖S5 株与Genbank 中的家鹅分离出的E. hermani 以及穴兔分离出的E.magna 基因型遗传距离为0.21,亲缘关系最近如表3。3 种球虫ITS 序列种间差异性较大,ITS序列可作为这3 种白额雁球虫鉴别的分子标记。

表3 22 种艾美耳球虫ITS 基因序列的遗传距离
构建的ML 和BI 系统进化树具有相似的拓扑结构和较高的支持率,其中ML 树分为3 枝,第1 枝包括分离自羊的E.arloingi,分离自博金斯牛的E.bovis,E.cylindrica 和E.canadensis。第2 枝包含分离自家鹅的E. hermani,分离自白额雁的E.tenlla,E.alabamensis 和E.mitis,分离自火鸡的E. dispersa,分离自家鹅的E. fulva,E. anseris 和E. stigmosa。第3 枝由分离自鸡的E. media,E.maxima,E. necatrix,分离自牛的E. wyomingensis 及 分 离 自 兔 的E.flavescens,E.piriformis 组成,该进化枝的宿主均为哺乳动物,形成了一个高度支持值的分枝。此外,艾美尔球虫菜子湖C70株(E.alabamensis)与Genbank 中的家鹅分离出的E. stigmosa(KP789173)和博金斯牛分离出的E.subspherica(AB769806)亲缘关系较近,在进化树中处于同一进化枝(自举值为99%);艾美尔球虫升金湖S49 株(E.tenlla)形成单独的分枝;艾美尔球虫升金湖S5 株(E.mitis)与Genbank 中的家鹅分离出的E.hermani(KF977223)以及穴兔分离出的E.magna(JX406876)亲缘关系较近,在进化树中处于同一进化枝上(自举值为99%)见图1。BI树的拓扑结构与ML 树相似见图2,3 种来自白额雁的艾美尔球虫均与家鹅中的艾美尔球虫亲缘关系较近,共同汇聚成一大进化枝,并且与寄生于哺乳动物如兔,山羊等艾美尔球虫物种距离较远。ML 树和BI 树的结果表明,艾美尔球虫ITS 基因序列在系统发生关系上也表现出较低的种内变异性和较高的种间变异性。

图1 最大似然法(ML)构建的艾美尔球虫属部分物种系统发育树
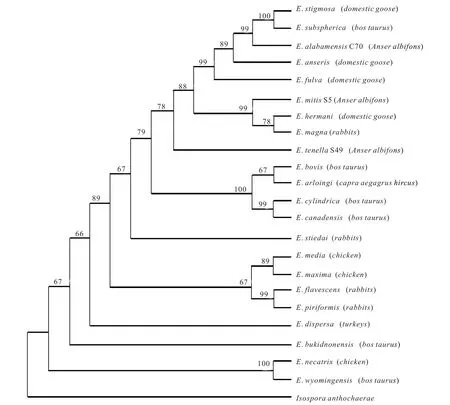
图2 贝叶斯法(BI)法构建的艾美尔球虫属部分物种系统发育树
3 讨论
寄生虫疾病已成为严重威胁野生动物和人类健康的全球性难题,研究表明,寄生虫不仅可以感染野生鸟类和家禽,而且具有感染哺乳动物和人的能力,对人类的威胁也在不断增加,具有重要的公共卫生意义[22]。分子标记法是根据寄生虫基因特征或者核苷酸序列的差异来确定物种和基因型,ITS 基因序列具有种间特异性和种内多样性,能精准反应种间或群体间的进化关系,因此,ITS基因是目前寄生虫鉴定的重要分子标记[23-24]。从基因水平证实艾美尔球虫菜子湖C70 株为椭圆艾美尔球虫(E. alabamensis),艾美尔球虫升金湖S49 株为柔嫩艾美尔球虫(E.tenlla),艾美尔球虫升金湖S5 株为和缓艾美尔球虫(E.mitis),3 株艾美尔球虫的ITS 序列与GenBank 上释放的其它球虫物种的ITS 序列相比变异性较低,与很多学者研究中的ITS 序列具有较高的稳定性和种间特异性的结论相一致[24-25]。利用ITS 基因进行分析,并且与GenBank 上已登录的艾美尔球虫序列相对比,发现与所获得的序列同源性均很高,从生物信息学角度证明ITS 基因序列可以提供有效的遗传数据,能够解决与鸟类相关的艾美尔球虫系统学和群体遗传学中的进一步问题。丰富了迁徙鸟类球虫的核糖体ITS 序列,为鸟类球虫的分类鉴定、分子流行病学调查及相关研究奠定基础。
应用ITS 基因不仅能够鉴定球虫的物种,还可以用来分析球虫不同物种之间的亲缘关系,为球虫的分子遗传学研究提供了理论基础[26]。目前对于迁徙鸟类体内球虫物种的系统发生关系的研究不多,且艾美尔球虫起源的也存在争议,系统发育树表明艾美尔球虫E.mitis,E.tenella 和E.alabamensis 与家鹅体内的艾美尔球虫汇聚为一枝,亲缘关系较密切,来自鸟类的E. mitis,E. tenella 和E.alabamensis 远离宿主为哺乳动物的E.arloingi,E. bovis,E.Cylindrica 以及E.canaclensis,表明树中所有艾美尔球虫均来自同一祖先,因此,本研究支持艾美尔球虫为单起源的观点[8-10]。其中一些物种聚集成与寄生虫宿主相关的进化枝(例如,从鸟粪便中分离的物种),且宿主为鸟类的球虫物种均汇聚为一大进化枝,而寄生于哺乳动物的球虫物种则形成一个单独的进化枝,并且发现寄生虫与宿主的进化程度也很相似,因此艾美尔球虫物种与宿主之间存在协同进化关系。
4 小结
本研究鉴定出越冬白额雁体内3 种艾美尔球虫并对其进行基因序列特征和部分物种的关系进行了分子系统学分析,证实了ITS 基因序列可作为鸟类艾美耳球虫鉴定可靠的遗传标记,也为寄生虫的物种鉴定及流行病学研究提供了重要的理论依据。同时我们利用生物信息学构建的系统发生树表明所有艾美尔球虫均来自同一祖先,支持艾美尔球虫为单起源观点。此外,本研究以粪便为非损伤取样,对粪便学的研究结果起到了补充作用。总之,研究野生越冬鸟类寄生虫具有重要的公共卫生意义,尤其是随着研究的不断深入,一些新的寄生虫将不断的被发现,而寄生虫各个虫种或基因型分子特性以及生物学特性的不断阐明,将有助于人兽共患寄生虫病互传机制的研究,从而为预防人寄生虫疾病乃至治疗人寄生虫疾病奠定必要的基础。
猜你喜欢
广东农业科学(2022年5期)2022-07-21
军事文摘(2021年22期)2022-01-18
中国动物保健(2021年8期)2021-11-24
时代英语·高一(2021年3期)2021-07-29
当代水产(2021年5期)2021-07-21
热带病与寄生虫学(2021年1期)2021-03-23
兽医导刊(2020年23期)2020-12-30
畜牧兽医学报(2015年3期)2015-07-05
中国预防兽医学报(2015年10期)2015-03-09
分忧(2014年9期)2014-09-22
