
抗条锈病小偃麦附加系CH366 的FISH 鉴定
2021-03-18 05:56贾举庆郭慧娟雷梦林张树伟常利芳畅志坚乔麟轶张晓军
山西农业科学 2021年3期
郑 瑗,李 欣,贾举庆,郭慧娟,雷梦林,张树伟,常利芳,畅志坚,乔麟轶,陈 芳,张晓军
(1.山西大学生物工程学院,山西太原 030006;2.山西农业大学农学院,作物遗传与分子改良山西省重点实验室,山西太原 030031;3.山西农业大学农业基因资源研究中心,农业部黄土高原作物基因资源与种质创制重点实验室,山西太原 030031)
小麦条锈病是由条锈病菌引起的靠风力传播的毁灭性病害,具有传播迅速、传播范围广、产量损失大的特点,严重危害了我国小麦的生产[1]。近年来,由于种植结构的改变,大量矮秆的小麦品种投入生产,使得株间环境更适合条锈病原菌传播,条锈病危害日益严重。由于条锈病小种的变异使得已有的抗条锈病基因逐渐丧失抗性,选育新的小麦抗条锈病基因资源并将其应用于育种与生产是解决条锈病危害的有效途径[2]。
中间偃麦草作为小麦的近缘物种,是小麦改良的三级基因源之一,抗寒,分蘖能力强,蕴含丰富的抗病基因。目前普遍认为,中间偃麦草的基因组型为JJJsJsSS,其中,J 基因组、JS基因组与小麦的B 基因组十分接近,因此,容易与小麦杂交,中间偃麦草应用十分广泛[3]。H.B.齐津[4]将小麦与中间偃麦草进行远缘杂交,选育出小冰麦系列新品种;孙善澄[5]利用小麦与中间偃麦草多次杂交后回交并定向选择的方法选育出中1~中5,在抗病方面的应用非常广泛。CHANG 等[6]利用普通小麦与中间偃麦草杂交获得TAI7044、TAI7045、TAI8335 等多个八倍体小麦,并继续与小麦杂交、回交,选育出CHadd7001、CHadd7002[7]、CH13-21[8]等多抗性附加系与易位系。石丁溧等[9]选育出抗白粉病和条锈病的异附加系AF-2,并通过GISH 证明了有2 条染色体来源于中间偃麦草。
CH366 是山西农业大学农学院利用普通小麦晋太170 与小麦-中间偃麦草部分双二倍体TAI7045 杂交,F1代与晋太170 再回交,回交后代经过多次自交得到的体细胞为44 条染色体的小偃麦新品系。本研究通过细胞学方法对CH366 进行了荧光原位杂交(FISH)鉴定,同时进行抗病性分析,以期为小麦遗传改良与抗病育种提供稳定且优良的中间材料。
1 材料和方法
1.1 试验材料
供试材料CH366、亲本晋太170、TAI7045 和对照品种台长29、SY95-71 均由作物遗传与分子改良山西省重点实验室保存并提供。CH366 是由晋太170 与TAI7045 杂交后回交得到;TAI7045 是由畅志坚用普通小麦和中间偃麦草杂交培育的部分双二倍体;晋太170 是由山西省农业科学院作物科学研究所选育的国审优质强筋小麦品种。
1.2 染色体制片
有丝分裂中期染色体制片参考HAN 等[10]的方法,略有改动。取50 粒CH366 种子在4 ℃冰箱放置2 d,室温下晾干。放在5 个培养皿中培养,每个培养皿铺2 层滤纸润湿后放入10 粒种子,在室温黑暗环境下培养2 d。待根尖长到2 cm 左右,放入4 ℃冰箱处理24 h。在每粒种子上剪取1 cm 左右的根尖分别放到2 mL 喷湿离心管中,然后在0.9 MPa N2O 罐处理2 h。用90%醋酸固定10 min,水洗3 次,切取根尖分生区置于2%的果胶酶和纤维素酶中,37 ℃水浴60~65 min。取出后用70%乙醇洗2 次,无水乙醇洗1 次。用解剖针捣碎根尖后,4 000 r/min离心3 min。倒掉乙醇后在室温下晾干,然后加入适量冰醋酸,搅拌均匀后用移液器吸取6.5 μL 混合液滴于载玻片上,15 min 后用显微镜观察染色体中期分裂相,选取染色体完整且分散好的片子保存。
1.3 荧光原位杂交分析
选取多聚寡核苷酸探针Oligo-pTa535-1 和Oligo-pSc119.2-1[11]进行双色荧光原位杂交,杂交程序参照LANG 等[12]的方法,并参照TANG 等[11]的方法对分散的染色体进行命名。用Leica DM2 荧光显微镜进行镜检,DFC500 CCD 拍照。选择分裂相明显且清晰的细胞进行核型分析,以获得可靠的证据。
1.4 抗病性鉴定
2019—2020 年度分别在四川省农业科学院现代农业科技示范园和山西农业大学东阳试验示范基地种植亲本晋太170、TAI7045 和CH366。在四川成都基地人工接种条锈菌最新流行小种条中32、条中33、条中34 混合小种,感病对照选择台长29,诱发品种为川育12,进行条锈病抗性鉴定。在东阳基地人工接种白粉菌株E09,感病对照是SY95-71,诱发品种为SY95-71,进行白粉病抗性鉴定。测试材料对条锈病和白粉病的抗性均按照6 级反应型进行记载[13-14],分别为0(免疫)、0;(近免疫)、1(高抗)、2(中抗)、3(中感)、4(高感)。
2 结果与分析
2.1 CH366 的染色体数目鉴定
取50 粒CH366 的种子发芽并取其根尖进行有丝分裂中期染色体制片,选取细胞完整且染色体分散好的片子进行染色体数目统计。由图1、2 可知,50 粒种子的染色体数目均为44 条,比普通小麦的42 条染色体多2 条,表明CH366 中可能含有2 条不属于普通小麦的染色体。
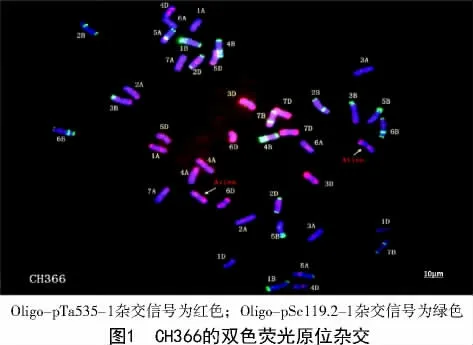

2.2 CH366 的FISH 鉴定
以分别标记了红色和绿色荧光标记的Oligo-pTa535-1 和Oligo-pSc119.2-1 探针对CH366进行荧光原位杂交(FISH),并对其染色体进行分组与核型分析,结果发现,CH366 中有42 条染色体与小麦亲本晋太170[15]染色体结构、荧光信号基本一致,另2 条染色体在长臂末端和短臂上接近着丝粒的位置含有较强的红色杂交信号,与普通小麦的染色体具有明显区别。且这2 条染色体形态与杂交信号完全一致,这表明CH366 中含有1 对来源于TAI7045 或中间偃麦草的染色体,CH366 可能是一个小麦-中间偃麦草异附加系。
2.3 CH366 的抗病性鉴定
在山西农业大学东阳试验示范基地进行CH366 成株期白粉病鉴定。结果显示,TAI7045 对白粉病的反应型为0 级,表现为免疫;CH366 和晋太170 的反应型均为4 级,表现为高感白粉病;说明CH366 中可能不含有抗白粉病基因。在四川省农业科学院现代农业科技示范园进行CH366 成株期条锈病鉴定。结果显示,晋太170 对条锈病的反应型为4 级,表现为高感条锈病。CH366 的反应型为0;级,表现为近免疫;TAI7045 的反应型为0 级,表现为免疫;CH366 与TAI7045 的条锈病抗性基本一致,表明CH366 中的抗条锈病基因可能来源于其所含有的1 对外源染色体,即来源于TAI7045 或中间偃麦草。
3 结论与讨论
小麦的远缘杂交可以将外源染色体导入小麦中,丰富小麦的遗传背景,创造新的种质资源,为发展小麦产业做出贡献。检验是否有外源染色体最直观、快速的方法就是原位杂交技术。建立FISH 核型图可以一目了然地观察到外源染色体形态与数目。最初,TSITSIN[16]通过远缘杂交创造出小麦-中间偃麦草部分双二倍体,之后有大批附加系、代换系如雨后春笋般涌现。韩方普等[17]利用原位杂交对小偃麦异附加系TAI-14 进行鉴定,发现变异的染色体来自于中间偃麦草,且附加系会再次发生变异,产生了双端体异附加系TAI-14t。杜万里[18]对一系列小麦-华山新麦草衍生后代进行原位杂交,筛选出全套二体附加系。本试验对小麦-中间偃麦草后代CH366 进行了FISH 鉴定和核型分析,证实CH366 是一个二体异附加系,包含小麦全套基因组,并添加了2 条外源染色体。添加的2 条外源染色体具体类型需要使用分子标记鉴定和其他技术手段来确定。
国内外学者通过中间偃麦草与小麦杂交选育出多个对小麦抗病、抗寒等性状具有改良作用的基因资源。GAUDERON[19]育成的小麦-中间偃麦草双二倍体TAF46,对小麦冻害与春季低温具有极强的抗性;孙善澄等选育的中4 和中5[5]含有黄矮病的抗性;LIU 等[20]利用普通小麦烟农15 与中间偃麦草育成的部分双二倍体E990256,高抗白粉病。部分双二倍体由于含有较多的外源染色体,农艺性状普遍较差,难以在育种或生产上直接应用。附加系作为外源基因向小麦转移的中间材料,仅含有1 对外源染色体,减少了外源染色体对小麦基因组的影响,更易于与小麦杂交,因此,附加系的选育对创制新的小麦种质资源、扩展小麦遗传基础具有重要意义。本试验选育并鉴定的异附加系CH366 对条锈病表现为近免疫,其小麦亲本晋太170 高感条锈病,因此,推测其条锈病抗性来自其所含有的外源染色体,即其抗条锈病基因来自中间偃麦草。但CH366 作为一个二体异附加系,仍含有较多的外源染色体,易受外源物种不良基因的影响,不利于在小麦育种中应用,因此,在生产实践中,需要利用其与普通小麦再次杂交,利用ph1b 基因诱导或辐射技术,依次创造出代换系和含有不良外源基因较少的小片段易位系。
猜你喜欢
作物学报(2022年2期)2022-11-06
热带作物学报(2022年7期)2022-08-06
舰船科学技术(2022年11期)2022-07-15
今日农业(2021年7期)2021-07-28
农民致富之友(2020年12期)2020-05-11
农民致富之友(2019年20期)2019-07-27
农民致富之友(2019年16期)2019-07-01
西部(2016年6期)2016-05-14
今日农药(2016年5期)2016-05-14
食品工业科技(2014年21期)2014-03-11
