
高粱组Ⅲ WRKY 转录因子对干旱胁迫的表达分析
2021-03-12 09:15
广东农业科学 2021年2期
(中国热带农业科学院湛江实验站/广东省旱作节水农业工程技术研究中心,广东 湛江 524013)
【研究意义】高粱(Sorghum bicolor)是禾本科高粱属1 年生草本植物,不仅是世界上重要的粮食和能源作物,也是优质的饲料作物。高粱具有耐旱、耐涝、耐盐碱等多种特性[1],其基因组较小(约750 Mb),具有丰富的遗传多样性,其优良的基因资源在作物改良中具有广阔的应用前景[2]。转录因子又称反式作用因子,对转录过程起诱导或抑制作用[3]。WRKY 转录因子作为植物中一类重要转录因子,广泛参与植物生长发育、响应激素诱导和生物及非生物胁迫等生物学进程[4],开展高粱WRKY基因耐旱性表达特征分析具有重要意义,可为高粱WRKY基因的功能研究奠定基础。【前人研究进展】WRKY基因家族作为高等植物中备受关注的基因家族之一,自第一个WRKY 转录因子基因从甘薯中克隆以来[5],随着许多物种基因组测序工作完成,越来越多的WRKY基因从基因组水平上鉴定出来,已在青稞[6]、中粒咖啡[7]、大麦[8]和杨树[9]基因组分别鉴定了41、49、98、122 个WRKY基因。WRKY 转录因子包含1~2 个保守WRKY 结构域,结构域由约60 个氨基酸序列组成,其保守的7 个核心氨基酸序列为“WRKYGQK”。根据WRKY保守结构域数目和锌指结构类型,可将WRKY 转录因子分为组Ⅰ、组Ⅱ和组Ⅲ三大类,其中组Ⅱ又分为5 个亚类(Ⅱa~Ⅱe)。组ⅠWRKY 转录因子包含2 个WRKY 结构域,其锌指结构的氨基酸类型为C2H2,组Ⅱ和组Ⅲ转录因子只含有1 个WRKY 结构域,锌指类型分别为C2H2、C2HC 型[10]。据报道,WRKY 转录因子参与了植物生长发育的许多进程,如开花[11]、次级代谢物合成[12]和衰老[13]等。也有较多研究表明,WRKY 转录因子响应各种压力胁迫,如玉米ZmWRKY62-like基因响应盐和干旱胁迫[14];小麦TaWRKY33受盐胁迫诱导表达,TaWRKY33转基因拟南芥比对照具有更好的耐盐性[15];甜高粱SSWRKY28、SSWRKY76基因的表达结果表明,它们可能在应答干旱胁迫时发挥一定作用[16];高粱SbWRKY30基因通过影响拟南芥和水稻的根系结构来提高对干旱胁迫的耐受性,拟南芥和水稻的转基因植株在干旱胁迫后脯氨酸含量、SOD、POD 和CAT 酶活性高于野生型植株,而MDA 含量低于野生型植株[17]。【本研究切入点】前人研究结果表明,组ⅢWRKY 成员广泛参与植物生长发育进程、生物和非生物胁迫应答反应[18]。赵兴奎等[19]已从高粱基因组鉴定了16个组ⅢWRKY成员,但没有研究其响应逆境胁迫的表达谱。【拟解决的关键问题】本研究以高粱品种BTx623 为试材,采用PEG6000 溶液模拟干旱胁迫[20-21],研究高粱组ⅢWRKY成员响应干旱胁迫的表达谱,为挖掘耐旱候选基因提供参考。
1 材料与方法
1.1 试验材料
供试高粱品种为BTx623,由广东省旱作节水农业工程技术研究中心保存,该材料有参考基因组序列,为中等耐旱性材料[22]。高粱组ⅢWRKY基因信息来自参考文献[19],基因序列下载 自Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)。
主要试剂:TransZol Plant 植物总RNA 纯化试剂盒、Trans2K® Plus DNA Marker、TransStart®Top Green qPCR SuperMix,购自北京全式金生物技术有限公司;ThermoScientific 反转录试剂盒K1622,购自Thermo Fermentas 公司。
1.2 试验方法
1.2.1 干旱胁迫处理 供试高粱种子用1%(W/W)NaClO 溶液浸泡10 min,然后用灭菌的蒸馏水冲洗干净,采用 0.5×Hoagland 营养液水培,植株在植物光照培养箱中培养,生长条件为28 ℃、12 h 光照、12 h 黑暗,光照强度为9600 lx。营养液每2 d 更换1 次,待植株生长至3 叶期,用含20%(W/W)PEG6000 的营养液水培,模拟干旱胁迫处理,于处理后0、1、3、6、12 h 收集整株植物,每个样本混合6 株植物,每个时间点取3个生物学重复样本。
1.2.2SbWRKY基因的表达谱分析 从NCBI 数据库(https://www.ncbi.nlm.nih.gov)下载有关干旱胁迫的高粱28K 芯片表达数据(GSE48205),利用GeneSpring GX 11.5软件(安捷伦科技有限公司)进行分析,获得探针的相对表达倍数,计算log2(fold change)值,然后根据探针对应的基因编号,整理基因的相对表达值。
1.2.3 总RNA 提取、cDNA 合成及荧光定量表达分析 采用植物总RNA 纯化试剂盒提取植株总RNA,对总RNA 用1.5%琼脂糖凝胶电泳检测RNA 完整性;以总RNA 为模板,使用反转录试剂盒获得cDNA,所用Marker 为Trans2K® Plus DNA Marker。以cDNA 为模板使用qRT-PCR Mix按照操作说明配制反应体系,在LightCycler480 Ⅱ实时荧光定量PCR 仪上进行反应,内参基因选用高粱GAPDH基因。PCR 反应体系(10 μL):cDNA 1 μL,2×Mix 5 μL,上游引物、下游引物(表1)各0.25 μL,补ddH2O 至10 μL;PCR扩增程序:95 ℃ 3 min;95 ℃ 10 s,60 ℃ 35 s,35 个循环。
1.2.4 qRT-PCR 结果统计分析 按照2-ΔΔCT法计算基因相对表达值[23]。使用EXCEL 的student’s T检验进行差异显著性分析。
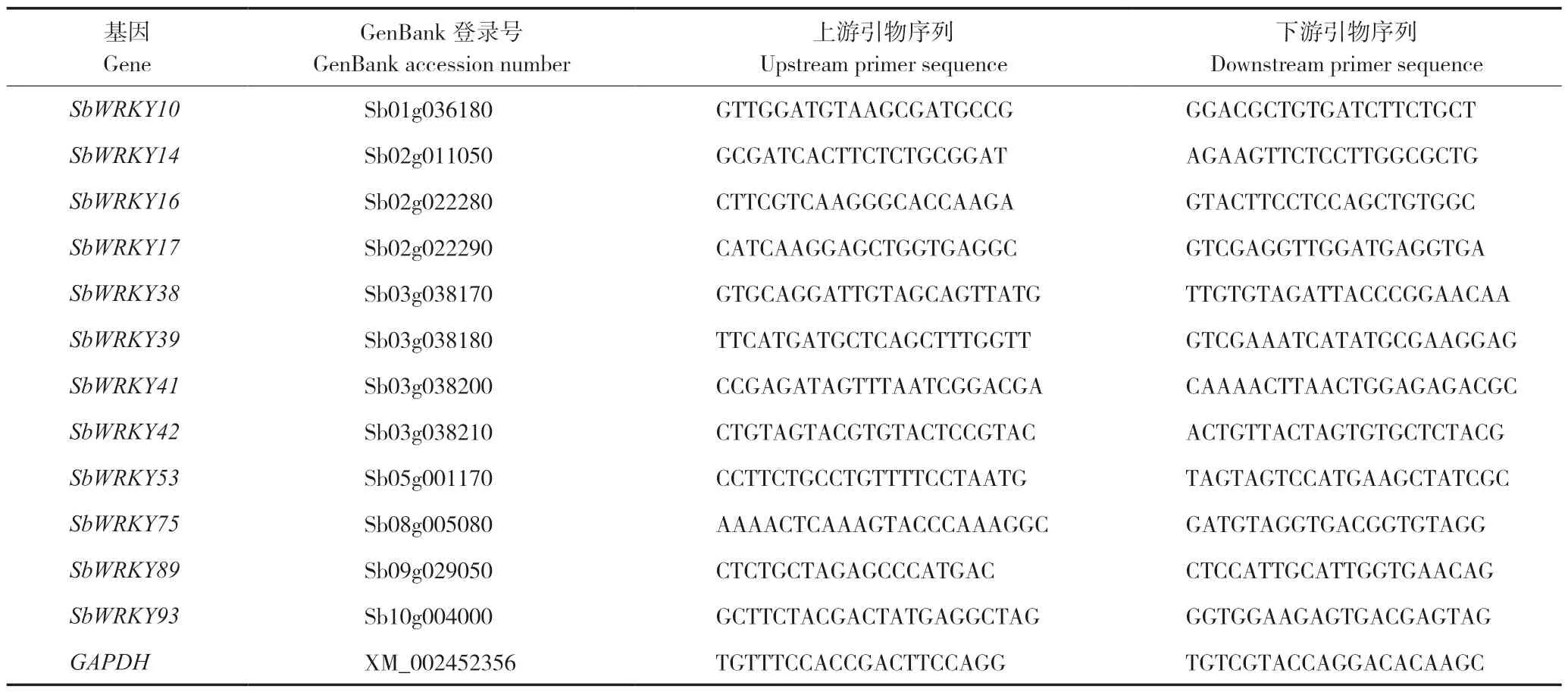
表1 高粱组Ⅲ WRKY 基因的qRT-PCR 扩增所用引物序列Table 1 Primer sequences for qRT-PCR amplification of Group III WRKY genes of sorghum
2 结果与分析
2.1 总RNA 完整性检测
为保证反转录和qRT-PCR 试验顺利开展,本研究对提取的15 个总RNA 样本进行琼脂糖凝胶电泳检测,其完整性较好(图1),可用于后续试验。
2.2 SbWRKY 的基因芯片分析结果
为研究高粱响应干旱胁迫的表达特征,从NCBI 数据库下载的高粱28K 芯片(GSE48205)包含对照、干旱胁迫、热胁迫和复合胁迫(干旱胁迫和热胁迫共同处理)共4 组表达值,将表达值导入GeneSpring GX 11.5 软件,经数据过滤、标准化处理、重要性分析和差异分析等流程,最后筛选得到4 个组ⅢSbWRKY基因的差异表达倍数值(表2)。由结果可知,SbWRKY14、SbWRKY32下调表达;SbWRKY39在干旱、热胁迫时下调表达,但在复合胁迫时上调表达;SbWRKY41在3 种胁迫处理时均上调表达。

图1 15 个RNA 样本的琼脂糖凝胶电泳检测Fig.1 Detection of 15 RNA samples by agarose gel electrophoresis
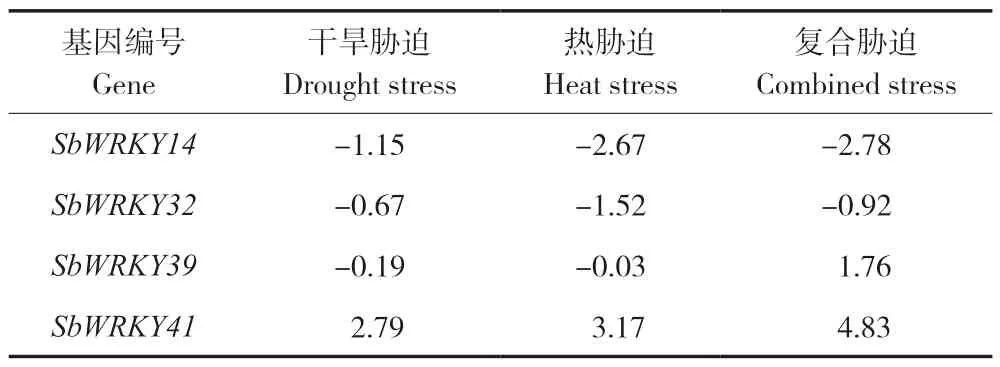
表2 不同非生物胁迫下高粱SbWRKY 基因表达值Table 2 Expression values of SbWRKYs in sorghum under different abiotic stresses
2.3 SbWRKY 基因的qRT-PCR 分析结果
为研究组ⅢSbWRKY基因响应干旱胁迫的表达谱,采用PEG6000模拟干旱胁迫于不同时间点取样,通过qRT-PCR分析基因处理与对照之间的相对表达值,在16个组Ⅲ成员中成功获得了12个基因的表达谱(图2);各基因在干旱胁迫处理后不同时间点与对照(0 h)相比,SbWRKY10在处理后3、6 h下调表达,而在处理后12 h上调表达;SbWRKY14在处理后1、6、12 h下调表达;SbWRKY16、SbWRKY38、SbWRKY41和SbWRKY42在处理后1、3、6、12 h均下调表达;SbWRKY17在处理后1、6、12 h下调表达;SbWRKY39和SbWRKY89在处理后1、3、12 h下调表达;SbWRKY53在处理后1、12 h下调表达,而在6 h上调表达;SbWRKY75在处理后1 h下调表达,而在3、6、12 h上调表达;SbWRKY93在处理后6 h上调表达。

图2 干旱胁迫下12 个SbWRKY 基因的表达谱Fig.2 Expression profiles of 12 SbWRKY genes under drought stress
3 讨论
WRKY 转录因子在响应各种非生物胁迫方面已有相关报道,WRKY 转录因子在植物对干旱胁迫的响应中起重要作用[24]。在拟南芥中过表达TaWRKY1和TaWRKY33激活了几个与逆境相关的下游基因,提高了拟南芥在各种胁迫下的萌发率,促进了根系的生长[25]。在烟草和梨中过表达PbrWRKY53增强了对干旱胁迫的耐受性。与野生型相比,转基因植株的活性氧生成相对更少、抗氧化酶活性和代谢产物更高。此外,在转基因烟草中过量表达PbrWRKY53导致PbrNCED1的表达水平提高;敲除PbrWRKY53可下调PbrNCED1的表达丰度,同时降低其耐旱性[26]。水稻组ⅢWRKY 转录因子OsWRKY11 在干旱和高温胁迫时诱导表达,以热激蛋白HSP101基因作启动子驱动OsWRKY11表达时,转基因植株耐热性和耐旱性增强,叶片萎蔫变慢,离体叶片失水较慢;结果表明OsWRKY11基因在应对高温和干旱胁迫反应中起重要作用,可能有助于提高植物的抗逆性[27]。
为了解SbWRKY基因在干旱胁迫调控中所起作用,本研究利用公共数据库基因芯片表达谱数据,分析高粱组ⅢWRKY基因在干旱胁迫、热胁迫和复合胁迫条件下的表达模式,2 个基因(SbWRKY14和SbWRKY32)在3 种处理条件下均下调表达,1 个基因(SbWRKY41)在3 种处理条件下上调表达。基因的表达趋势通常反映其相应的功能,在雷蒙德棉中,干旱胁迫处理后,有16 个基因(其中包含3 个组Ⅲ成员)诱导表达,15 个基因表达量减少[28];本研究通过qRTPCR 技术检测出12 个基因的表达谱,其中4 个基 因(SbWRKY10、SbWRKY53、SbWRKY75和SbWRKY93)在1 个或多个时间点诱导表达,而4个基因(SbWRKY16、SbWRKY38、SbWRKY41和SbWRKY42)在所有时间点均抑制表达。这些结果表明,不同的SbWRKY基因在应对干旱胁迫时可能起不同的调控作用。SbWRKY41的qRT-PCR检测结果与基因芯片检测的表达谱呈相反表达趋势,可能是由于两个研究采用的材料、干旱处理方法不同所致:本研究所用试材为BTx623,以20% PEG6000 模拟干旱胁迫;而芯片试验所用试材为R16,采用自然干旱胁迫处理[29]。
此外,WRKY 转录因子广泛参与了植物生长发育、各种生物及非生物胁迫进程,如在水稻中已验证了16 个OsWRKY 转录因子的功能,结果表现为Ⅰ类参与抗病进程,Ⅱb 类具有抗病和抗逆功能,Ⅱd 类调控植物的营养生长,而Ⅲ类具有调控营养生长和抗逆等功能[30]。本研究只开展了Ⅲ类基因在干旱胁迫条件下的表达谱分析,这些成员还可能响应低温、高温和高盐等胁迫反应并发挥一定作用。为深入了解这些基因的功能,后续还需克隆这些基因,通过转基因实验验证其生物学功能。
4 结论
高粱WRKY 转录因子基因在应对逆境胁迫中起重要调控作用,本研究在前人研究基础上挑选16 个高粱组ⅢWRKY 转录因子基因开展进一步研究。通过基因芯片分析结果表明,SbWRKY14和SbWRKY32在3 种处理条件下全部下调表达,表明这2 个WRKY基因在干旱胁迫、热胁迫和复合胁迫下具有相似功能;SbWRKY41在3 种胁迫条件下诱导表达。为进一步分析该组成员的表达谱,以20% PEG6000 模拟干旱胁迫,在0、1、3、6、12 h 取样,采用qRT-PCR 检测出该组12 个基因的表达谱;SbWRKY10、SbWRKY53、SbWRKY75和SbWRKY93在1 个或多个时间点诱导表达,而SbWRKY16、SbWRKY38、SbWRKY41和SbWRKY42在所有时间点均抑制表达。通过这两种分析方法,筛选到诱导和抑制表达基因,这为我们今后深入了解SbWRKY基因在干旱胁迫中的调控功能提供参考,也为我们克隆耐旱相关基因提供候选基因。
猜你喜欢
军事文摘(2022年16期)2022-08-24
湖北农业科学(2022年11期)2022-07-18
今日农业(2022年4期)2022-06-01
青年文学家(2022年1期)2022-03-11
今日农业(2021年20期)2021-11-26
今日农业(2021年14期)2021-10-14
今日农业(2021年11期)2021-08-13
疯狂英语·新读写(2021年2期)2021-02-25
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
