
牙髓炎的发病机制和分子靶标研究
2021-03-08 06:31努尔比亚木麦麦提依明
中国医药导报 2021年1期
努尔比亚木·麦麦提依明 吴 龙 赵 今
新疆医科大学第一附属医院牙体牙髓科,新疆乌鲁木齐 830000
牙髓是一种特殊的结缔组织[1]。牙髓也会产生炎症反应[2]。牙髓炎是影响牙髓组织的最常见的病理性疾病[3-4]。目前针对牙髓炎的诊断和治疗具有一定的阻碍[5]。细菌感染是引起牙髓炎的主要原因[6]。牙髓防御系统中可以决定牙髓炎症的程度[7]。细胞因子和趋化因子积聚可能促进牙髓炎的进展[8-9]。目前对牙髓炎的分子失调机制仍然没有达成共识。本文从GEO 数据库的两个基因表达谱中整合了牙髓炎与正常牙髓组织的差异表达基因。通过生物信息学的分析方法探究牙髓炎的关键失调基因和分子机制。
1 资料与方法
1.1 数据来源及分析
从GEO 数据库(http://www.NCBI.nlm.nih.gov/GEO/)收集牙髓炎数据集GSE77459(2016 年2 月)和GSE9 2681(2017 年12 月)。使用“limma”R 包(version 3.40.6)进行差异分析。设定阈值P <0.05 和log2FC>1 获得差异表达基因(DEGs)。从蛋白质相互作用数据库(http://string-db.org)(version 11.0)筛选只包含差异基因的数据,设置得分为>500。利用Cytoscape 3.5.1 软件进行网络分析,得到基因的连接度(degree)。
1.2 富集分析和基因集富集分析(GSEA)
使用“cluster Profiler”R 包(version 3.12.0)进行基因本体论(GO)分析。使用Cytoscape 3.5.1 分析京都基因与基因组百科全书(KEGG)信号通路。通过Broad Institute(http://software.broadinstitute.org/gsea/)进行GSEA,筛选KEGG 信号通路。
1.3 样本收集
各收集3 例牙髓炎及健康的牙髓组织。所有的样本均获得了患者的知情同意并获得了新疆医科大学第一附属医院医学伦理委员会批准。
1.4 实时定量PCR(qRT-PCR)
利用Trizol(Invitrogen)提取牙髓组织总RNA,并用PrimescriptTMRT Reagent Kit(TaKaRa)逆转录cDNA。通过SYBR Premix Ex TaqTM(TaKaRa)进行qRT-PCR反应。引物见表1。使用2-ΔΔCT的方法分析数据。

表1 引物序列
1.5 Western blot(WB)
通过RIPA 裂解液提取牙髓组织总蛋白。采用凝胶电泳分离等量蛋白质并转移到聚偏二氟乙烯膜上。5%脱脂牛奶封闭1 h 后在4℃下孵育一抗过夜。冲洗后与二抗孵育2 h。通过化学发光试剂(Millipore)观察膜上的免疫反应蛋白。β-actin 为内参蛋白。抗体均购自Abcam 公司。所有实验至少重复3 次。主要抗体如下:PTPRC(Abcam)、CD86(Abcam)、CD44(Abcam)、ERK1/2(Abcam)、p-ERK1/2(Thr202/Tyr204;Abcam)、β-actin(Abcam)。
1.6 统计学方法
使用R 3.6.1 进行数据分析,使用SPSS 20.0 软件进行统计分析。采用student t 检验比较两组之间的差异。以P <0.05 为差异有统计学意义。
2 结果
2.1 牙髓炎组织中的差异表达基因
GSE77459 中牙髓炎和健康对照之间有1280 个DEGs(图1A)。差异变化倍数最大的前10 个基因见表2。GSEA 结果显示,牙髓炎组织中Toll 样受体、MAPK、JAK-STAT 信号通路明显升高(图1B)。通过GSE92681 的DEGs 验证到81 个基因(图1C)。它们可能是牙髓炎潜在的失调因子。

表2 GSE77459 中差异变化倍数较大的前10 个基因
2.2 生物功能和通路
GO 功能富集分析共识别了201 个生物学进程(BP)(图2A),42 个细胞成分(CC)(图2B)和6 个分子功能(MF)(图2C)。它们主要集中于T 细胞活化和ERK1/2 通路的正向调节、白细胞介素-2(IL-2)等细胞因子产生的调控等生物过程。另外,获得的11 个KEGG 信号通路则反映了牙髓炎的差异基因主要与免疫和炎症反应相关(图2D)。
2.3 PPI 网络构建与枢纽基因鉴定
潜在失调基因的PPI 网络中包括55 个基因(图3A)。通过网络的degree 鉴定了前3 个基因作为关键失调基因,包括PTPRC、CD86 和CD44。此外,它们在两套数据中的表达方向一致(图3B)。
2.4 关键基因及信号通路验证
通过qRT-PCR 和WB 验证了CD44 在牙髓炎组织中显著表达上调(图4A)。前文发现CD44 正向调控ERK1/2 通路。在牙髓炎中ERK1/2 通路被激活(图4B)。因此,CD44 可能通过ERK1/2 途径促进牙髓炎的疾病进程。
3 讨论
牙髓对外部因素很敏感,如龋齿引起的微生物感染等刺激[10]。这些因素可能引发炎症级联,进而通过神经源性炎症向疼痛方向发展[11]。因此,深入了解牙髓炎症过程对发展牙科诊断治疗是至关重要的。本研究对牙髓炎和对照的测序数据进行了系统分析,鉴定了在牙髓炎中具有重要作用的关键分子和信号通路。
差异变化倍数最大的两个基因中,IGKV1-5 在牙髓炎组织中表达显著上调。有研究显示其在牙周炎组织中表达上调,是与炎症和疼痛有关的分子[12-14]。显著下调基因NR1D1 可以调控NLRP 3 炎症小体的活性[15],还与增强巨噬细胞的抗细菌特性有关[16]。有81 个基因在两套数据中同时差异表达。本研究认为这些基因更有可能是牙髓炎过程中的失调基因。另一方面,从GSEA 和富集分析结果中发现,牙髓炎相关的失调基因主要参与免疫炎症相关的生物功能和信号通路。在牙髓感染的情况下,趋化白细胞使溶酶体酶增加,从而造成组织损伤[17]。Toll 样受体4(TLR 4)通路参与大鼠急性牙髓炎所致伤害性感觉[18]。此外,TLR 4 的刺激还会引发一系列信号级联,诱导肿瘤坏死因子-α和IL-1 的释放[19-21]。
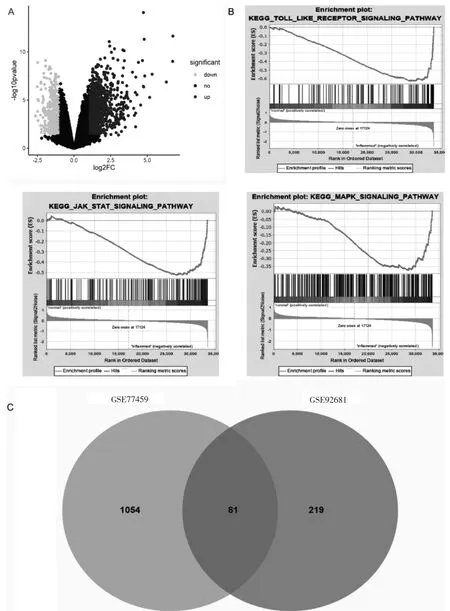
图1 牙髓炎组织的差异表达基因
牙髓炎相关失调基因的PPI 网络分析发现,大部分基因具有相互作用关系,可能在牙髓炎过程中具有协同作用。在PPI 网络中的核心因子中,qRT-PCR 结果验证了CD44 的表达差异。CD44 是一种广泛表达的黏附分子,已有研究报道其在牙髓炎组织中的表达显著上调[22]。CD44 正向调控ERK1 和ERK2 级联反应,对炎症具有促进作用[23]。本研究识别了CD44 通过ERK级联反应促进牙髓炎的进程。这一过程应进行深入研究,以了解其作为标志物和治疗靶点的潜力和影响。
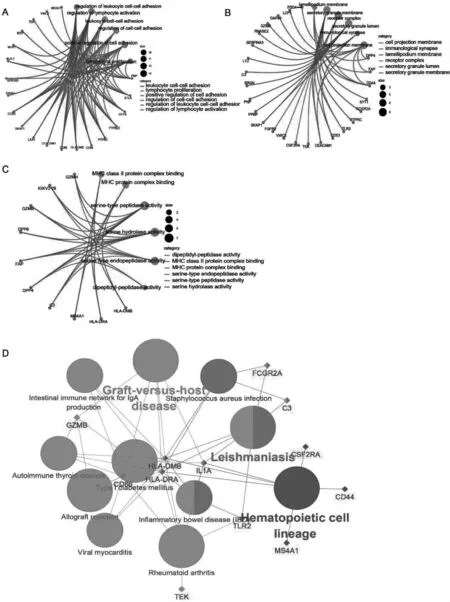
图2 失调基因的富集结果

图4 验证关键失调基因

猜你喜欢
中华养生保健(2020年10期)2021-01-18
天津医科大学学报(2019年3期)2019-08-13
中华老年口腔医学杂志(2016年4期)2017-01-15
电子制作(2016年15期)2017-01-15
海南医学(2016年8期)2016-06-08
系统工程与电子技术(2016年2期)2016-04-16
中国卫生标准管理(2015年14期)2016-01-15
中国医疗美容(2015年2期)2015-07-19
中国医疗美容(2015年2期)2015-07-19
电测与仪表(2014年1期)2014-04-04
