
空肠弯曲杆菌感染小鼠模型的研究进展
2021-03-06 09:17:30程雪婷陈妍灵黄晓东
中国实验动物学报 2021年1期
程雪婷陈妍灵黄晓东∗
(1. 武汉大学中南医院,武汉 430071; 2. 武汉大学同仁医院,武汉 430060)
空肠弯曲杆菌是一种人兽共患的食源性病原菌,主要与家禽和家畜有关[1-2]。根据感染的分离株类型、细菌数量和宿主免疫状态不同,导致患者有不同程度的胃肠炎、伴或不伴血性腹泻、发烧、腹痛[3]。空肠弯曲杆菌感染通常是一种自限性疾病[4],少数情况下可以导致格林—巴利综合征(Guillain-Barré syndrome,GBS)、反应性关节炎和肠易激综合征或乳糜泻在内的长期后遗症[5]。由于世界范围内感染数量的增加,抗生素抗性分离株[6]和感染后后遗症的出现,这种细菌已成为威胁人类健康的重要病原体。
尽管小鼠是常用的微生物感染模型,但是常规的空肠弯曲杆菌小鼠感染模型存在定植困难或不出现疾病表现等缺陷[6],导致我们缺乏有效的小鼠感染模型,阻碍了弯曲菌病的研究。成功构建小鼠感染模型需要我们对感染模型建立的关键因素进行了解,并对常规小鼠模型进行改良。宿主与微生物相互作用是常见的影响细菌定植和感染后临床表现的原因[7],本文从菌株和小鼠两个角度出发,综述了影响空肠弯曲杆菌感染模型建立的关键因素,以期为成功建立合适的小鼠感染模型提供参考。
1 菌株对构建感染模型的影响
1.1 与定植相关的毒力因子
空肠弯曲杆菌的一些毒力因子可以影响细菌的粘附、侵袭、定植等多种功能,对于该菌的致病性至关重要[8]。粘附被认为是空肠弯曲菌在鸡[9]、人[10]及鼠[11]的和定植中起重要作用的因素。空肠弯曲菌合成了几种粘附素:纤维连接蛋白结合外膜蛋白F(fibronectin-binding outer membrane protein,CadF)以及纤维蛋白A(fibrillarin A,FlpA)被认为是重要的毒力因子,能促进空肠弯曲杆菌对宿主细胞的粘附,并使其能够在高水平下定植[12-13]。侵袭对于空肠弯曲菌的毒力有重要作用,丝氨酸蛋白酶(high-temperature requirement factor A,HtrA)通过破坏肠道紧密连接,造成肠道屏障缺陷,有利于空肠弯曲杆菌侵袭[14]。Heimesaat 等[15]发现用HtrA 突变型菌株感染幼鼠,幼鼠的肝、肾、肺部出现的炎症变化均低于野生株。鞭毛蛋白A(flagellin A,FlaA)、鞭毛蛋白B(flagellin A,FlaB)和鞭毛生物合成蛋白(flagellar biosynthesis protein,FlhF)参与组成鞭毛被认为对于宿主粘附和侵入起作用。Schmidt等[16]的研究发现FlaA 和FlaB 缺陷型小鼠并不影响细菌定植,但是对于引起弯曲菌病的临床表现是必不可少的。Ren 等[17]的研究发现FlhF 失活导致空肠弯曲杆菌在鸡的定植减少且完全丧失在小鼠中的连续定植能力。此外,空肠弯曲菌的特定代谢能力对于其在各种宿主的肠道定植中起重要作用,一些代谢相关基因的缺失会导致细菌生长或定植受限[18-19]。常见的代谢相关基因包括:过氧化氢酶(catalase, katA)、 超 氧 化 物 歧 化 酶(superoxide dismutase,sodB)、天门冬氨酸/谷氨酸结合蛋白(aspartate/glutamate-binding protein,peb1A)、天冬氨酸酶(aspartoacylase, aspA)、 天冬氨酸转氨酶(aspartate transaminase,aspB)等(表1)。对菌株的基因分析可帮助鉴定那些在小鼠模型中易感的空肠弯曲杆菌的分离株。
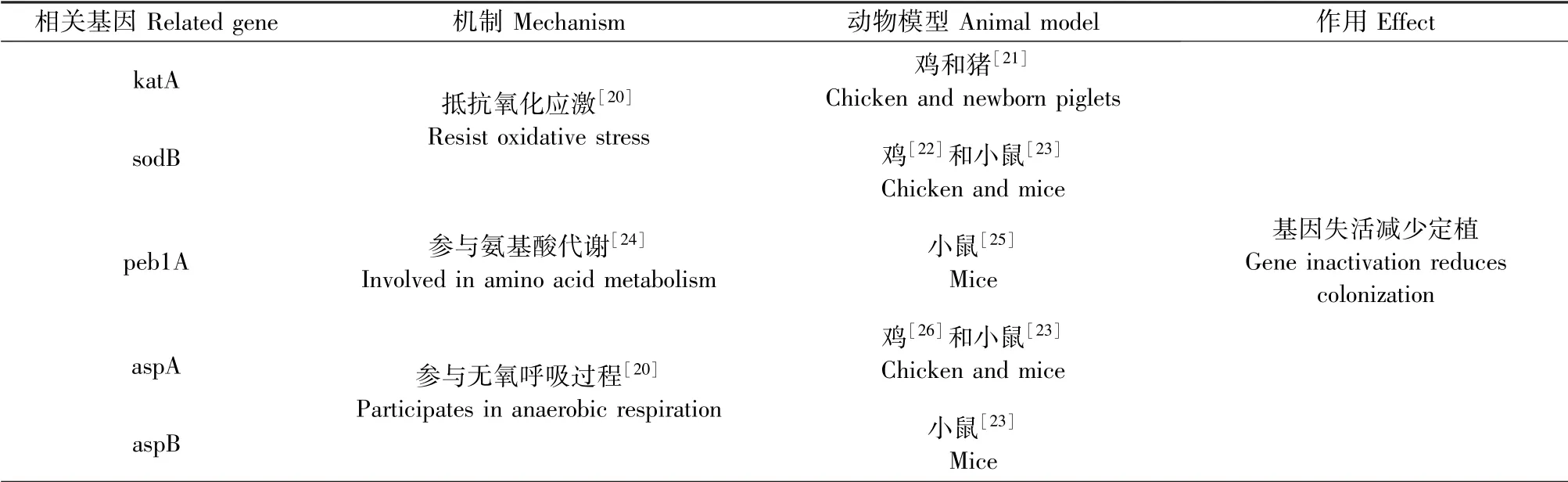
表1 影响空肠弯曲杆菌在宿主定植和生长的代谢相关基因Figure 1 Metabolic related genes that influence the colonization and growth of Campylobacter jejuni in the host
1.2 与疾病表现相关的毒力因子
空肠弯曲杆菌感染后的不同临床表现与单个分离株的不同毒力潜力相关。一部分空肠弯曲杆菌分离株通过产生一种紧密连接毒素(zonula occludens toxin,ZOT)能靶向破坏细胞间紧密连接,被认为可能导致腹泻[27-28]。小鼠实验中证实,一些临床分离株可以在小鼠肝中定植和诱发肝炎,这可能与菌株产生的肝毒性因子有关[29-30]。目前对于肝毒性因子研究较少,具体成分未知。以往研究表明, 空 肠 弯 曲 杆 菌 表 达 的 脂 寡 糖(lipooligosaccharide,LOS)诱导与神经组织中神经节苷脂的分子模拟被认为诱导交叉反应抗体,导致易感宿主触发急性多发性神经病-格林-巴利综合征(GBS)[31],Godschalk 等[32]发现GBS 相关菌株中特定的LOS 基因座的表达过高,并伴随神经节苷脂模拟结构的表达增加,提示LOS 基因座与GBS 和神经节苷脂模拟物有关。有研究发现空肠弯曲杆菌可通过细胞致死肿胀毒素促进结直肠癌的发生[33],这表明一些空肠弯曲杆菌毒素有潜在致癌性。不同毒力因子可能导致不同疾病表现,这在我们构建不同疾病模型时需要考虑。
2 宿主因素对构建感染模型的影响
2.1 小鼠品系
小鼠的品系可以对空肠弯曲杆菌感染的敏感性产生影响。Vuckovic 等[34]比较了三种不同品系小鼠(BALB/c、C57BL/6 和DBA/2)感染空肠弯曲杆菌后的临床表现。结果显示,在感染小鼠的肝和脾可以看到细菌和炎症反应,BALB/c 小鼠最明显,此外,BALB/c 小鼠原发性肝感染的持续时间最长。Baqar 等[35]通过小鼠鼻内感染比较了空肠弯曲杆菌菌株81-176 对BALB/c、C3H/Hej、CBA/CAJ 或C58 J三种小鼠品系的感染毒性,BALB/c 小鼠组的死亡率最高,C58 J 组死亡率最低,BALB/c 小鼠表现出比其它小鼠更强的毒性反应。以上结果表明不同小鼠品系对空肠弯曲杆菌的敏感性不同,虽然BALB/c 小鼠感染后的炎症反应相对更明显,但是其对细菌毒性的不耐受可能限制了其作为动物模型的利用。
2.2 免疫缺陷
常规小鼠作为空肠弯曲杆菌感染模型时不存在显著炎性/免疫应答反应[36],这可能是小鼠先天免疫系统对空肠弯曲杆菌的耐受性更高[37]。然而,免疫缺陷鼠对空肠弯曲杆菌等人类病原体敏感更高,利用基因敲除技术构建免疫缺陷小鼠也常应用于构建空肠弯曲杆菌感染模型[38]。目前常用的是IL-10 缺陷的小鼠模型[39-40],空肠弯曲杆菌可以在IL-10-/-小鼠模型稳定地定居,具有高致病性负荷,并患有非自限性急性小肠结肠炎,出现血性炎性腹泻,因为它模仿了人弯曲菌病的关键特征,常被用于空肠弯曲杆菌的研究。然而,IL-10-/-小鼠模型必须在无菌环境下饲养,因为它倾向于受自身肠道微生物影响而发展成自发性结肠炎,使这个模型存在缺陷[41]。SIGIRR-/-小鼠模型的建立可以避免上述缺陷,空肠弯曲杆菌在这些小鼠的定植率很高,肠道表现出明显的炎症组织学迹象,病理学上类似于人类空肠弯曲杆菌的急性感染,也是研究空肠弯曲杆菌导致胃肠道表现的有利模型[41-42]。
2.3 肠道菌群
空肠弯曲杆菌定植除了与宿主细胞有关,也与宿主的肠道菌群有关。常规的鼠感染模型中存在定植困难或不出现疾病表现,这可能是由于是鼠微生物群提供的定植抗性所致[43-44]。人们普遍认为“定植抗性”是由于原有的肠道菌群在和新来的病原体竞争肠道资源时处于优势,一旦原有微生物群落破裂,就会有足够的营养和场地以供空肠弯曲杆菌栖息,通过去除竞争性共生细菌可以促进定居[45]。许多研究结果支持这一假设,无菌或微生物区系有限的小鼠很容易被空肠弯曲杆菌定植[46]。在感染前利用单一抗生素[47-49]或抗生素合剂[46]也可以使小鼠更容易感染空肠弯曲杆菌。在使用抗生素后对小鼠的肠道微生物组进行分析后发现,微生物多样性水平下降,尤其是粪肠球菌[8]。进一步研究发现特定的厌氧菌菌群可以预防空肠弯曲杆菌引起的结肠炎,给小鼠服用对厌氧菌敏感的克林霉素可以诱导空肠弯曲杆菌致结肠炎[48]。Mohan等[50]发现益生菌可以减少小鼠的感染率和降低肠道炎症程度,这可能得益于益生菌增加了肠道微生物丰富度,提升了定植抗性。此外,婴儿小鼠容易感染空肠弯曲杆菌和肠道炎症[15],这表明早期的微生物群可能无法提供与“成人”菌群相同的保护作用。Bereswill 等[46]发现用“人源化”菌群代替常规饲养的小鼠的菌群时,它有助于空肠弯曲菌的定植。这种机制尚不清楚,推测可能是“人源化”微生物群可能提供了促进空肠弯曲杆菌定植的营养优势。综上,宿主的肠道菌群稳态和丰富的菌群多样性不利于空肠弯曲杆菌感染,肠道无菌状态、肠道微生物多样化水平下降、人源化肠道菌群都有利于空肠弯曲杆菌感染。
2.4 饮食组成
饮食组成对肠道菌群的影响也会对空肠弯曲菌定植产生影响[39]。Brown 等[51]发现用低脂/低蛋白饮食建立的小鼠营养不良模型进行的实验证明营养不良会导致肠道屏障功能下降,肠道通透性增加,并引起小肠微生物群和代谢改变,从而增加了肠道感染的易感性。Bereswill 等[52]发现喂食“西式饮食”的小鼠以及具有常规微生物群的肥胖小鼠会使小鼠更容易感染空肠弯曲杆菌。最近报道了锌缺乏对抗生素预处理小鼠中空肠弯曲杆菌感染的症状形成有直接影响[53],此外,与正常患者相比,克罗恩病患者血清锌浓度更低并对空肠弯曲杆菌更加易感[54],提示缺锌可能会增加宿主对空肠弯曲杆菌易感性。
2.5 宿主适应
空肠弯曲杆菌可以通过宿主适应增强对宿主感染,这可能是因为宿主的免疫应答对细菌造成选择性压力,间接的筛选出了那些更能适应宿主的细菌[37]。Bell 等[41]发现通过分离小鼠粪便的空肠弯曲杆菌进行培养后再感染小鼠,这种连续传代培养重复四次,小鼠发病率和疾病严重程度增加。进一步研究表明空肠弯曲杆菌在经过宿主适应后,一些关键的突变基因会促进菌株在宿主的定植[55-56]。这提示我们经过宿主适应的分离株可能更有利于细菌在小鼠的成功定植。
3 结语
空肠弯曲杆菌的不同毒力因子通过影响细菌的粘附性、侵袭性以及生长代谢等功能,对细菌的定植以及产生不同疾病表现产生作用。以往研究已经证实一些基因在空肠弯曲杆菌的宿主定植和产生不同疾病表现中发挥关键作用。通过分析空肠弯曲杆菌的基因信息可以帮助我们在小鼠模型中鉴定易感的细菌分离株及构建特定疾病表现的模型。常规小鼠模型对于空肠弯曲杆菌存在定植抗性,通过小鼠的基因改造、使用幼鼠、改变小鼠饮食、利用无菌鼠或感染前使用抗生素、菌株的连续传代和采用人源化菌群等方式有利于构建小鼠感染模型,目前应用最广泛的方式是基因改造、应用GF 级小鼠和感染前使用抗生素,鉴于基因改造和GF 级小鼠的成本较高,最廉价的方式是使用抗生素。菌株的连续传代和采用人源化菌群虽然可增强对宿主感染,但其操作复杂、周期长,因而目前并不常用。此外,虽然饮食改造的方式目前并不常用,但其成本低廉且操作简单,相信在未来可以有更广泛的应用。通过对有利于构建小鼠感染模型的因素进行分析,可以帮助我们寻找更加方便、廉价和更能反映疾病特征的小鼠模型,为弯曲菌病的研究提供科学基础。
猜你喜欢
世界科学技术-中医药现代化(2021年9期)2021-12-31 03:31:02
云南化工(2021年6期)2021-12-21 07:31:04
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
农药科学与管理(2019年6期)2019-11-23 08:17:12
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
华南农业大学学报(2015年5期)2015-12-04 03:04:38
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国当代医药(2015年33期)2015-03-01 02:09:29
河南医学研究(2014年5期)2014-02-27 14:52:44
