
长期菌渣化肥配施对稻田土壤酶活性的影响及交互效应
2021-03-04 02:01陈文博王旭东石思博季诗域叶正钱任泽涛
浙江农林大学学报 2021年1期
陈文博,王旭东,石思博,季诗域,叶正钱,任泽涛,刘 璋
(1. 浙江农林大学 环境与资源学院 浙江省污染土壤修复重点实验室,浙江 杭州 311300;2. 南京农业大学 资源与环境科学学院,江苏 南京 210095)
土壤酶在土壤生态系统中起着关键作用,是源自于微生物、动植物活体或残体的一类生物活性物质,能够催化土壤中的生物化学反应[1]。土壤酶活性作为反映土壤中各种生物化学过程的方向和强度的指标[2],参与了有机质分解、养分循环等重要机制。其中土壤过氧化氢酶、蔗糖酶、脲酶活性对评价土壤肥力水平有着重要的意义。蔗糖酶又名转化酶,参与土壤中碳水化合物的转化,是土壤碳循环重要的酶类之一;脲酶是唯一对尿素的水解转化具有催化作用的酶,是参与土壤氮循环的重要酶类之一;过氧化氢酶作为一种重要的氧化还原酶,参与土壤中物质循环和能量转换的过程,也对土壤碳循环有一定的促进作用。土壤酶活性受施肥、土壤类型[3]、栽培制度[4−5]和管理措施[6−8]等因素的密切影响,外源有机物料的添加及化肥的施用对土壤酶活性的影响较大。菌渣是一种特殊的有机物料,是在农业生产中最丰富但也容易被忽视的生物质资源。菌渣还田,不仅能保持土壤肥力,同时能达到土地改良的效果。菌渣施入土壤,能提供大量的菌丝残体、纤维素、半纤维素、木质素和矿物质等,同时也是丰富的氨基酸、多糖等营养物质的来源[9]。但因人们对其营养价值认识不足,还田技术仍比较滞后而导致其常被随地丢弃或燃烧,造成了资源的极大浪费。中国目前菌渣利用率仅为33%[10],菌渣不恰当处置导致散发出温室气体和霉菌孢子蔓延等问题,造成生态环境污染[11]。因而研究菌渣化肥配施对稻田土壤酶活性的影响,促进农业废弃物的高效利用,加快农业可持续发展,具有重要的理论和现实意义。已有研究表明:菌渣化肥配施能有效提高土壤pH,增加土壤养分含量[12],达到改土增肥的目的。温广婵等[13]研究表明:配施菌渣有利于土壤有机质和其他速效养分的积累。马嘉伟等[14]研究发现:菌渣化肥配施能有效提高土壤养分,促进生育时期内水稻Oryza sativa生长。龚臣等[15]研究发现:菌渣化肥配施不仅对稻田土壤有机碳组分有显著影响,而且还影响了土壤肥力。石思博等[11]研究发现:菌渣化肥配施对土壤微生物量碳氮、可溶性碳氮和矿化碳含量均有显著影响。综上所述,以往研究多数注重菌渣还田对土壤理化性质和水稻生长的影响,而关于菌渣如何对酶活性产生影响,特别是酶活性如何随着水稻生育时期发生变化,从而影响水稻生长或养分吸收的机制目前还不清楚。在此之前,对菌渣化肥配施对稻田土壤微生物量碳氮、可溶性碳氮和矿化碳含量[16]的影响已有研究,发现菌渣还田对土壤微生物量碳氮、可溶性碳氮和矿化碳含量都有显著的影响,但是要准确揭示土壤有机碳氮的周转和养分循环,还需要对长期菌渣化肥配施后土壤酶活性的变化进行系统研究。
1 材料与方法
1.1 材料与设计
试验地点位于浙江省嘉兴市秀洲区王店镇五浪园水稻田 (30°37′~30°40′N,120°39′~120°44′E,平均海拔4 m)。该地四季分明,年平均日照2 017.0 h,气候温和,属于东亚季风区,冬夏季风交替显著,常年平均气温为15.9 ℃,年平均降水量1 168.6 mm。长期定位试验始于2010年,供试土壤在定位试验前(2010 年)的基本理化性质为:pH 6.80,有机质 25.16 g·kg−1,碱解氮 94.29 mg·kg−1,有效磷 37.01 mg·kg−1,速效钾127.22 mg·kg−1。土壤耕作方式是休耕轮作,土壤质地为粉砂质黏土,其中砂粒、粉粒、黏粒的相对含量分别为29.38%、47.37%和23.25%。供试菌渣选用种植黑木耳Auricularia auricula后的桑Morus alba枝屑物料经发酵后的产物。主要成分为:有机碳451.8 g·kg−1,全氮11.4 g·kg−1,全磷1.0 g·kg−1,全钾6.0 g·kg−1,碳氮比为39.6。菌渣的原料由5个典型成分组成:纤维素、半纤维素、木质素、灰分和蛋白质,质量分数分别为28.21%、20.16%、15.11%、14.83%和14.3%。供试水稻品种为‘甬优 1540’‘Yongyou 1540’。
本试验采用双因素随机区组设计,分别设3个化肥施用水平和菌渣还田水平,共9个处理,每个处理重复3次,共27个独立试验小区,单个小区面积为20 m2。施用菌渣量为0、10、20 t·hm−2,分别记为 0%(F0对照)、50%(F50)、100%(F100);化肥用量分别为当地常规施肥量的 0%(C0)、50%(C50)、100%(C100)。各处理具体见表1。结合当地单季稻-休闲的水稻轮作制度,移栽水稻前将土壤进行翻耕,同时施入不同比例混合的菌渣和化肥(基肥)。常规化肥100%施用量具体如下:基肥施碳酸氢铵(含氮质量分数17%)300 kg·hm−2;分蘖初期(7月23日)追施尿素(含氮质量分数46%)150 kg·hm−2;分蘖盛期(9 月 4 日)追施控释复合肥{[m(氮)∶m(磷)∶m(钾)]=22∶11∶13}225 kg·hm−2。50% 化肥处理则减半进行。

表1 试验处理Table 1 Experimental treatments
1.2 样品采集及分析方法
分别于水稻移栽前2周(2017年6月)、分蘖盛期(2017年9月)、灌浆后期(2017年11月)、收获期(2017年12月)在各个小区0~20 cm土层多点采集土壤。土样在阴凉处风干、磨碎、装袋,以备用于常规土壤理化性质分析;同时取新鲜原状土分析土壤容重,进行土壤酶活性的分析。
土壤pH按土水质量比为1.0∶2.5酸度计测定;有机质采用重铬酸钾外加热法测定;全氮采用半微量开氏法测定;碱解氮采取碱解扩散法测定;有效磷采用氟化铵-盐酸浸提-钼锑抗比色法测定;速效钾采用醋酸铵浸提-火焰光度法测定[17];土壤蔗糖酶、过氧化氢酶、脲酶活性分别采用3,5二硝基水杨酸比色法、高锰酸钾滴定法、靛酚蓝比色法测定。过氧化氢酶活性以每克土消耗0.1 mol·L高锰酸钾毫升数表示,蔗糖酶活性以24 h后每克土葡萄糖的毫克数表示,脲酶活性以24 h后每克土氨氮的毫克数表示[18]。
1.3 统计方法
数据处理采用Origin 8.5、SPSS 18.0和R 3.3.3进行统计分析和图像绘制,采用双因素方差分析(two-way ANOVA)和Duncan新复极差法进行差异显著性检验,通径分析采用Excel 2016。
2 结果与分析
2.1 菌渣化肥配施对土壤过氧化氢酶活性的影响
由表2可知:水稻不同生育时期各处理中土壤过氧化氢酶活性为3.01~10.20 mL·g−1,从高到低依次表现为收获期、分蘖盛期、灌浆后期、移栽前期。不同时期同一处理土壤过氧化氢酶活性呈现出在移栽前上升,至分蘖盛期达到高峰,灌浆后开始下降,收获期则再次升高的变化趋势。在水稻移栽后,大部分处理的土壤过氧化氢酶活性均高于对照(C0F0),其中C100F50处理极显著高于对照(P<0.01)。在分蘖盛期,各处理土壤的过氧化氢酶活性均呈上升趋势,其中C50F100、C100F0、C100F50处理涨幅最明显。到灌浆后期,除C100F100处理的土壤过氧化氢酶活性提高之外,其余处理均呈下降趋势,其中C100F0、C100F50处理降幅比较明显。在化肥施用 C0水平下,菌渣施入量越多,水稻发育前期土壤过氧化氢酶活性也随之越高,但后期差异不显著;在化肥施用 C50水平下,菌渣施入量越多,土壤过氧化氢酶活性随之增加。在菌渣施用F0和 F50水平下,随着化肥施用量的增加,过氧化氢酶活性表现为先升高后降低的趋势。在化肥施用C100和菌渣施用F100的水平下,过氧化氢酶活性随着菌渣化肥施用量的增加,同样表现为先增后减的趋势。单施菌渣和单施化肥以及其两者的交互作用对土壤过氧化氢酶活性在水稻的4个生育时期均有显著(P<0.05)或极显著影响(P<0.01)。在水稻发育的4个时期,单施化肥对土壤过氧化氢酶活性的影响不显著,但是单施菌渣对土壤过氧化氢酶活性均具有显著(P<0.05)或极显著影响(P<0.01)。菌渣与化肥互作效应对土壤过氧化氢酶活性在水稻灌浆后期和收获期有极显著影响(P<0.01)。

表2 菌渣化肥配施对土壤过氧化氢酶活性的影响和互作效应Table 2 Effect and interaction of the application of fungal residue and chemical fertilizer on catalase activity in soil
2.2 菌渣化肥配施对土壤蔗糖酶活性的影响
从表3可以看出:水稻不同生育时期各处理中土壤蔗糖酶活性为0.20~2.04 mg·g−1,从高到低依次表现为灌浆后期、分蘖盛期、收获期、移栽前期。在水稻整个生育期,各处理的土壤蔗糖酶活性呈现出先升高后降低的趋势。在水稻移栽后,大部分处理的土壤蔗糖酶活性均高于对照处理(C0F0),其中C50F100处理显著高于对照处理(P<0.05)。在灌浆后期,各处理土壤的蔗糖酶活性均呈上升趋势,其中C50F100、C100F50、C100F100处理涨幅最明显。到收获期,各处理的土壤蔗糖酶活性显著降低,其中处理C50F50、C50F100降幅最大,并且C50F100处理的土壤蔗糖酶活性最大。在化肥施用 C0水平下,随菌渣施入量增加,土壤蔗糖酶活性前期随之降低,但到后期又逐渐升高,但差异并不显著;在化肥施用 C50水平下,除移栽前期之外,其他3个时期土壤蔗糖酶活性都随菌渣施入量的增加而增加。在化肥施用C100和菌渣施用F100的水平下,水稻生育前期,随着菌渣化肥施用量的增加,土壤中蔗糖酶活性随之增加,但水稻发育后期却逐渐下降。单施菌渣和单施化肥以及其两者的交互作用对土壤蔗糖酶活性在水稻的4个生育时期均有显著(P<0.05)或极显著(P<0.01)影响。在水稻发育的4个时期,单施化肥对土壤蔗糖酶活性均具有显著影响(P<0.05),但单施菌渣对土壤蔗糖酶活性影响不显著。菌渣与化肥互作效应对土壤蔗糖酶活性在水稻移栽前期和收获期有极显著影响(P<0.01)。

表3 菌渣化肥配施对土壤蔗糖酶活性的影响和互作效应Table 3 Effect and interaction of the application of fungal residue and chemical fertilizer on soil invertase activity

表4 菌渣化肥配施对土壤脲酶活性的影响和互作效应Table 4 Effect and interaction of the application of fungal residue and chemical fertilizer on soil urease activity
2.3 菌渣化肥配施对土壤脲酶活性的影响
由表4可知:水稻不同生育时期各处理中土壤脲酶活性为 0.54~4.80 mg·g−1。与移栽前期相比,水稻土在分蘖盛期、灌浆后期、收获期各处理的脲酶活性规律比较相似,差异不显著。除C0F0、C50F50、C0F100处理外,其他处理的土壤脲酶活性从移栽前至分蘖盛期呈持续下降趋势,到了灌浆后期开始上升,上升趋势持续至收获期。在水稻移栽后,大部分处理的土壤脲酶活性均高于对照(C0F0),并在移栽前期,各处理的土壤脲酶活性极显著高于对照(P<0.01),其中以C100F50处理最大。各处理的土壤脲酶活性在移栽前期之后均有所下降,在分蘖盛期最小,其中C50F100、C100F50、C100F100处理降幅最明显。在化肥施用C0水平下,菌渣施入量多,土壤中脲酶活性差异不显著。在化肥施用C50水平下,随着菌渣施入量增多,土壤脲酶活性从移栽前开始下降,到收获期又开始回升。在化肥施用C100和菌渣施用F100的水平下,土壤中脲酶活性随着菌渣和化肥施用量的增加,在不同时期均表现出先增加后减少的趋势。单施菌渣和单施化肥以及两者的交互作用对土壤脲酶活性在水稻的4个生育时期均有显著(P<0.05)或极显著(P<0.01)影响。在水稻的4个生育时期,单施化肥对土壤脲酶活性均具有显著(P<0.05)或极显著(P<0.01)影响,并且单施菌渣对土壤脲酶活性也均具有极显著(P<0.01)影响。除了灌浆后期之外,菌渣化肥配施对土壤脲酶活性在其他3个时期都有显著(P<0.05)或极显著(P<0.01)影响。在水稻发育前中期,菌渣化肥配施对土壤脲酶活性的互作效应高于蔗糖酶和过氧化氢酶。
2.4 土壤酶与土壤养分的通径分析
从图1A可以看出:菌渣化肥配施对各理化因子的间接通径系数,除土壤有机碳、全氮、有效磷为负值外,其他均为正值。碱解氮对配施效应最大(1.00),其次是pH(0.17)。速效钾较低(0.07)。各因素对配施效应的间接效益(绝对值)大小顺序依次为碱解氮、全氮、有机碳、pH、有效磷、速效钾。蔗糖酶对各理化因子的直接通径系数,除有机碳、全氮、有效磷为负值,其他均为正值,碱解氮对蔗糖酶活性效应最大(1.80),其次是速效钾(0.52),pH较低(0.49)。各因素对蔗糖酶活性的直接效益(绝对值)大小顺序依次为碱解氮、全氮、有机碳、速效钾、pH、有效磷。
图1B表明:菌渣化肥配施对各理化因子的间接通径系数,除速效钾为负值外,其余均为正值。有效磷对配施效应最大(0.30),其次是全氮(0.24),有机碳最低(0.03)。各因素对配施效应的间接效益(绝对值)大小顺序依次为有效磷、全氮、碱解氮、pH、有机碳、速效钾。过氧化氢酶对各理化因子的直接通径系数,除pH为负值外,其余均为正值,有效磷对过氧化氢酶活性效应最大(0.69),其次是全氮(0.17)和碱解氮(0.17),速效钾较低(0.05)。各因素对过氧化氢酶活性的直接效益(绝对值)大小顺序依次为有效磷、pH、全氮和碱解氮、有机碳、速效钾。
由图1C可知:菌渣化肥配施对各理化因子的间接通径系数,除pH为负值外,其余均为正值,有效磷对配施效应最大(0.38),其次是全氮(0.09),速效钾较低(0.01)。各因素对配施效应的间接效益(绝对值)大小顺序依次为有效磷、pH、碱解氮、全氮、有机碳、速效钾。脲酶对各理化因子的直接通径系数,除速效钾为负值外,其他均为正值,全氮对脲酶活性效应最大(0.56),其次是有效磷(0.54),有机碳较低。各因素对脲酶活性的直接效益(绝对值)大小顺序依次为全氮、有效磷、速效钾、pH和碱解氮、有机碳。
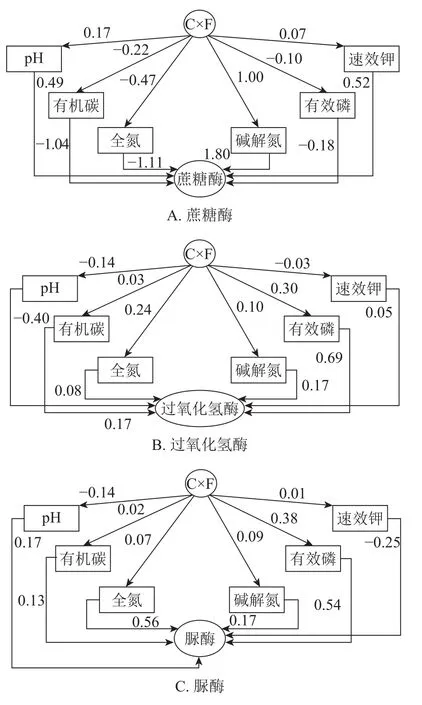
图1 不同土壤理化因子与土壤酶的通径分析Figure 1 Path analysis of different soil physical and chemical factors and soil enzyme
3 讨论
3.1 菌渣化肥配施对土壤过氧化氢酶活性的影响
过氧化氢酶参与土壤中物质循环和能量转化等过程,在一定程度上可以作为表征土壤生物氧化过程强弱的指标[19]。陈闯等[20]研究认为:在作物发育中后期,有机肥混施能提高土壤过氧化氢酶活性。有研究表明:有机物料还田能有效提高土壤过氧化氢酶活性[21−22]。本研究发现:在化肥施用C0、C50和菌渣施用 F0、F50水平下,过氧化氢酶活性在移栽前期升高,到分蘖盛期、灌浆后期开始下降,最后到收获期上升到最大。这说明与单施化肥或菌渣相比,中等量菌渣化肥处理的过氧化氢酶活性较好,因为菌渣与化肥配施有利于菌渣在土壤中转化和积累,土壤氧化还原能力较强[23]。在化肥施用C100和菌渣施用F100的水平下,随着菌渣和化肥施用量增加,土壤过氧化氢酶活性上升到一定状态时也会下降,说明就提高过氧化氢酶活性而言,中等量菌渣化肥配施优于高量菌渣化肥配施。这可能是由于化学肥料的过量使用影响了土壤酶活性[24−25]。单施有机肥和单施化肥均能提高土壤过氧化氢酶活性。本研究发现:在水稻发育中后期,菌渣与化肥互作效应对土壤过氧化氢酶活性有极显著影响,一方面因为在水稻发育中后期,水稻根系生长加快,土壤生物呼吸作用增强;另一方面由于菌渣和化肥配施后,微生物与酶的代谢活动加强,导致过氧化氢酶活性也随之增加[26−28]。
3.2 菌渣化肥配施对土壤蔗糖酶活性的影响
蔗糖酶是参与土壤碳循环的重要酶类之一[21]。程曼等[26]研究认为:长期不同秸秆还田能提高土壤蔗糖酶活性。也有研究表明:长期施肥能够提高土壤蔗糖酶活性[29]。本研究发现:在化肥施用C0、C50和菌渣施用 F0、F50水平下,随着菌渣和化肥施用量的增加,除了分蘖盛期之外,其他时期土壤蔗糖酶活性增加。这说明与单施化肥或菌渣相比,中等量菌渣化肥配施处理的蔗糖酶活性效果好。主要是因为菌渣作为土壤中碳库,为蔗糖酶提供了更多的酶促基质,菌渣还田使稻田土壤有机质和养分含量显著增加,并且菌渣中含有大量有机碳,刺激了微生物活性。在化肥施用C100和菌渣施用F100的水平下,随着菌渣化肥施用量的增加,土壤中蔗糖酶活性也在增加,并在不同时期均表现出先增加后减少的趋势。说明菌渣化肥配施显著提高了土壤蔗糖酶活性,但高量菌渣化肥配施下土壤蔗糖酶活性的贡献反而降低。一方面是因为菌渣本身富含纤维素类等有机成分,致使土壤中有机质增多,从而更加有利于蔗糖酶活性的增加[30];另一方面可能是由于高量菌渣下含有较高的养分,碳氮比较高,不适于微生物分解,限制了有机物质形成腐殖质并释放养分[30]。单施有机肥和单施化肥均能提高土壤蔗糖酶活性。本研究发现:在水稻发育中后期,菌渣与化肥互作效应对土壤蔗糖酶活性有极显著影响,这可能与水稻的生长需求和菌渣还田的养分释放规律有关。
3.3 菌渣化肥配施对土壤脲酶活性的影响
脲酶是一种酰胺酶,是参与土壤氮循环的重要酶类之一[31]。李东坡等[32]研究认为:脲酶活性在作物生长的前、中期能够明显表明培肥方式对土壤生物活性产生的效果。本研究发现:在化肥施用C0、C50和菌渣施用 F0、F50水平下,土壤脲酶活性随着菌渣和化肥施用量的增加而增加,但在化肥施用C100和菌渣施用F100的水平下表现出先增加后减少的趋势。这说明菌渣化肥配施能够显著提高土壤脲酶活性,但高量菌渣化肥配施下土壤脲酶活性的贡献反而降低。一方面是因为菌渣腐解可以为土壤提供丰富的有机碳,并且可以为微生物活动和酶活性的提高创造有利的条件[33];另一方面可能是因为菌渣本身的碳氮比较高,微生物分解需要更多的化肥氮或来自土壤的矿质氮,从而在分解过程会与水稻生长过程之间产生争氮现象。有研究表明:单施有机肥和单施化肥均能提高土壤脲酶活性,但是与单施化肥相比,有机肥与化肥相结合对土壤脲酶活性有显著提高的作用[34−35]。本研究发现:在水稻分蘖盛期,菌渣化肥配施对土壤脲酶活性的互作效应高于蔗糖酶和过氧化氢酶。说明在3种酶中,菌渣化肥配施更有利于脲酶活性的增强,暗示菌渣化肥配施处理下,在水稻生育前期,土壤微生物对氮循环的直接影响较大。
3.4 土壤酶与土壤养分的通径分析
通径分析是回归分析的拓展,是具有方向性的相关系数,能表示原因与结果(自变量与因变量)之间的关系,可以用它来估计自变量对应变量直接影响效应的大小,比较其相对重要性[36]。为了进一步研究土壤基本理化性质对土壤酶影响的数量关系,其相关系数可以分解为直接作用系数和间接作用系数,从而揭示各变量对因变量的相对作用。本研究中,菌渣化肥配施对各理化因子的间接通径系数发现:配施对碱解氮(1.00)这条路径影响最大。说明菌渣化肥配施主要通过提高速效氮质量分数来促进酶活性。可能因为菌渣促进了氮矿化相关酶的活性,从而加快了有机质的分解和有效氮的释放。过氧化氢酶对各理化因子的直接通径系数,有效磷对过氧化氢酶这条路径影响最大(0.69)。说明有效磷通过直接效应调控土壤过氧化氢酶活性。菌渣的施入有利于耕层土壤有机质的累积,对土壤磷素有活化作用;另外,菌渣还田还增加了磷素向土壤返回的通量,且有机肥中磷的有效性高于土壤磷[37],从而改善土壤理化性质及酶活性。蔗糖酶对各理化因子的直接通径系数,碱解氮(1.80)对蔗糖酶这条路径影响最大,说明碱解氮通过直接效应调控土壤蔗糖酶活性。增施菌渣特别是在水稻生长中后期能使土壤中有效氮质量分数显著增加,随着菌渣施入量增加,土壤供氮能力也随之增加,但并不是越多越好,而是有机无机配施效果更好,更加有利于水稻吸收利用。脲酶对各理化因子的直接通径系数,全氮对脲酶这条路径影响最大(0.56)。说明全氮通过直接效应调控土壤脲酶活性。因为脲酶是参与土壤氮循环重要的酶类之一,它可将土壤有机物中碳—氮键水解生成氨、二氧化碳和水,其活性的提高能促进土壤中的有机氮向有效氮的转化[38]。
4 结论
菌渣化肥配施对土壤过氧化氢酶、蔗糖酶、脲酶在水稻不同生育时期有着不同的影响,提高脲酶活性的作用主要体现在水稻生长前中期,而蔗糖酶、过氧化氢酶主要体现在后期,所以在农业生态系统碳、氮循环中,菌渣化肥配施对水稻发育前中期有助于氮循环,后期有助于碳循环。
菌渣化肥配施显著提高了土壤过氧化氢酶、蔗糖酶、脲酶的活性,但高量菌渣化肥配施对土壤过氧化氢酶、蔗糖酶、脲酶的贡献反而降低。
土壤有效磷、碱解氮和全氮通过直接效应调控土壤过氧化氢酶、蔗糖酶和脲酶活性,菌渣化肥配施主要通过提高碱解氮质量分数促进土壤酶活性。菌渣化肥配施对脲酶的交互效应高于过氧化氢酶和蔗糖酶,所以在3种酶中,菌渣化肥配施更有利于脲酶活性的增强。
综合来看,C100F50处理是提高土壤酶活性和碳氮循环的最佳选择。
猜你喜欢
山西农业大学学报(自然科学版)(2021年6期)2021-12-28
中国农业科技导报(2021年10期)2021-12-07
腐植酸(2021年2期)2021-12-04
中国稻米(2021年5期)2021-10-09
数理化解题研究·高中版(2021年5期)2021-09-10
云南农业(2021年5期)2021-06-19
新疆农业科学(2020年6期)2020-07-02
食药用菌(2020年1期)2020-02-22
数学学习与研究(2019年20期)2019-11-30
安全与环境工程(2018年4期)2018-08-08
