
盐碱胁迫下施加生物炭和减少氮肥用量对甜菜光合特性的影响
2021-03-04 03:38:24刘新宇武沛然李彩凤王玉波张树友陈婧婷
西北农林科技大学学报(自然科学版) 2021年1期
刘新宇,武沛然,李彩凤,王玉波,张树友,陈婧婷
(东北农业大学 农学院,黑龙江 哈尔滨 150000)
随着地球环境的持续恶化,各种问题应运而生,土壤盐碱化现已成为全球性的环境问题,日益侵占并威胁着人类有限的生活空间。如何改良和利用盐碱地已经成为当前农业面临的主要问题。我国东北地区的盐碱地面积为7.66×106hm2,约占全国盐碱地面积的7.9%[1]。光合速率下降是导致盐碱胁迫植物生长缓慢的主要原因之一。盐碱胁迫能破坏植物的叶绿体结构[2],叶片中盐分的积累导致植物叶绿素含量下降,影响植物的光系统,降低植物接收光能的能力[3],使植物生长发育缓慢甚至死亡。
甜菜(BetavulgarisL.)是世界两大糖料作物之一,也是我国北方地区重要的经济作物,本身具有一定的抗盐碱能力。目前有关盐碱胁迫对甜菜生长影响的研究较多,有研究表明,低盐环境可以促进甜菜生长,高盐环境下甜菜生长则会受到抑制[4]。盐碱胁迫会显著抑制甜菜的光合作用[5],施加有机肥能有效缓解盐碱胁迫对甜菜光合作用的抑制,并提高甜菜的产量[6]。
生物炭是在缺氧或绝氧条件下,将生物有机材料在高温下热裂解产生的固体[7]。在农业生产中,可以将生物炭按一定比例与市场上的氮磷钾肥混合,形成新型的炭基有机、无机复合肥。生物炭与化肥混合成粒后,两者紧密结合,可以使肥效缓慢释放并减少化肥的流失,从而提高化肥利用率,减少环境污染[8]。由于生物炭能提高土壤的腐殖质和有机质含量,具备“锁”碳能力,从而使土壤肥力和二氧化碳含量提高[9],间接提高植物的光合能力。如果在逆境 (盐碱或渍害) 条件下施加生物炭,则能降低土壤体积质量,增加土壤营养成分含量,同时提高作物产量[10-11]。另外,在施加生物炭的条件下同时减少施肥量,也能显著提高玉米产量[12]。
中国黑龙江松嫩平原盐碱土面积大,对其进行修复及开发利用具有十分重要的实际意义,但是目前在甜菜栽培上尚缺乏进一步有效提高其耐盐碱能力的技术措施,从而限制了甜菜在该地区的种植、开发和利用。为此,本试验在盐碱胁迫条件下,通过生物炭与氮肥配施,研究甜菜叶绿素含量、RuBP羧化酶活性、净光合速率及叶绿素荧光参数的变化规律,探索盐碱胁迫下施加生物炭和减少氮肥用量对甜菜光合特性的影响,以期为甜菜抗盐碱栽培及其生理研究提供理论依据。
1 材料与方法
1.1 试验材料
供试甜菜品种为KWS0143,由德国KWS公司生产。生物炭由玉米秸秆在500 ℃高温低氧下制成,pH值为9.29,碳含量为67.08%,由东北农业大学资源与环境学院提供。
1.2 试验设计
试验于2018年在黑龙江省哈尔滨市东北农业大学试验站(126°63′W,45°10′N)进行。供试土壤为本地自然黑钙土,土壤基础肥力(0~20 cm)为:有机质含量35.66 g/kg,碱解氮含量176.27 mg/kg,速效磷含量83.30 mg/kg,速效钾含量153.20 mg/kg,土壤土水比1∶5浸提液的电导率(EC1∶5)为12.2 mS/m,pH值为7.65。
将供试土壤与中性盐(Na2SO4∶NaCl=1∶2(物质的量比,下同)) 和碱性盐(Na2CO3∶NaHCO3=1∶2) 混合,此时土壤中Na+含量为3 g/kg,EC1∶5为33.2 mS/m,pH值为9.15。
试验共设7个处理,其中3个正常施肥处理,分别为黑钙土(CK)、盐碱胁迫处理(ALK)及盐碱胁迫下施加3%生物炭(N180);4个盐碱胁迫下施加3%生物炭并同时减少氮肥用量处理,其施肥量分别为162 kg/hm2(N162)、144 kg/hm2(N144)、126 kg/hm2(N126)和108 kg/hm2(N108),其中生物炭施用量为风干土壤质量(每桶10 kg) 的3%,正常施肥的施用量比例为N∶P2O5∶K2O=180∶90∶90,减少氮肥施用处理的磷、钾肥施用量与正常施肥处理相同。肥料分别使用尿素、磷酸二胺和硫酸钾。将各处理的盐碱、化肥和生物炭与过筛后的土壤混匀后装入桶中,桶的直径为30.0 cm,高为26.5 cm。播种前每桶浇水2 L (浇透),4月31日进行人工播种,每桶播5穴,每穴播10粒种子,每个处理40桶。待第2对真叶展开时间苗,每桶留1株。生育期内浇水、除草同大田管理。
在甜菜整个生育期内,分别在06-22,07-11,08-05,08-25和09-15,随机选择各处理长势相同的3株甜菜进行取样。
1.3 测定指标及方法
1.3.1 叶绿素含量的测定 取甜菜的第3对真叶,采用乙醇提取法[13]测定甜菜叶片中的叶绿素含量。
1.3.2 RuBP羧化酶活性的测定 RuBP羧化酶活性测定采用酶联免疫分析法,并使用上海酶联生物科技有限公司提供的试剂盒进行测定。
1.3.3 净光合速率和气孔导度的测定 在取样前后的晴朗天气,使用便携式光合作用-荧光测量系统 (GFS-3000,上海泽泉) ,于09:00-11:00测定甜菜的净光合速率和气孔导度。控制光合有效辐射为1 500 μmol/(m2·s),叶室CO2浓度为390 μmol/mol。
1.3.4 叶绿素荧光参数的测定 在取样前后的晴朗天气,于09:00-11:00使用便携式叶绿素荧光仪(PAM-2500,上海泽泉) 测定叶绿素荧光参数,包括光系统Ⅱ最大光能转换效率(Fv/Fm)、光系统Ⅱ实际光能转换效率(Y(Ⅱ))、光系统Ⅱ相对电子传递速率(ETR)、光化学淬灭系数(qL)和非化学淬灭系数(NPQ)。
1.3.5 干物质量的测定 每处理取3株长势相同植株,用蒸馏水冲洗干净,将地上部与地下部分开后,在烘箱中杀青30 min (105 ℃),80 ℃烘干至恒质量后进行测定,取平均值为植株的干物质量。
1.4 数据分析
图表中数据均为3次重复的“平均值±标准误”。用Excel 2016进行数据处理和制图,采用SPSS 23.0软件进行相关性分析和5%水平的差异显著性分析。
2 结果与分析
2.1 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶绿素含量的影响
由表1可见,就整个生育期而言,各处理甜菜叶绿素含量均呈先升高后降低趋势,CK处理的叶绿素含量在整个生育期均显著高于各盐碱胁迫处理。

表1 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶绿素含量的影响
表1显示,在6月22日,N180、N162、N144和N126处理甜菜叶绿素含量显著高于ALK处理,说明盐碱胁迫下施加生物炭能显著增加甜菜的叶绿素含量。在7月11日,N180、N162、N144和N126处理甜菜叶绿素含量较ALK处理分别提高了31.9%,36.9%,32.4%和19.8%,是各处理叶绿素含量较ALK处理增幅最大的时期。与7月11日相比,N162和N144处理甜菜叶绿素含量在8月5日略有下降,其他处理则仍呈上升趋势。在8月25日,ALK处理甜菜的叶绿素含量与除CK外的其他处理差异不显著,但各处理甜菜叶绿素含量均有明显下降。在9月15日,各处理甜菜叶绿素含量均达到最低值。
2.2 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜RuBP羧化酶活性的影响
由表2可知,CK处理甜菜RuBP羧化酶活性在各取样时期分别较ALK处理高1.8%(06-22),6.7%(07-11),24.6%(08-05),24.7%(08-25)和19.7%(09-15),说明盐碱胁迫会降低甜菜RuBP羧化酶的活性。6月22日,ALK处理甜菜的RuBP羧化酶活性与其他处理无显著差异。8月5日,在盐碱胁迫条件下随施氮量的降低甜菜RuBP羧化酶活性均逐渐降低,其中N180和N162处理均高于ALK处理,而N108处理显著低于ALK处理。在8月25日,ALK处理甜菜的RuBP羧化酶活性显著低于其他处理。在9月15日,CK、ALK、N180、N162、N144、N126处理甜菜的RuBP羧化酶活性均高于8月25日,其中N180、N162、N144、N126处理甜菜的RuBP羧化酶活性分别较ALK处理高16.6%,11.9%,6.3%和3.2%。

表2 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜RuBP羧化酶活性的影响
2.3 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片光合特性的影响
2.3.1 净光合速率 表3显示,随着生育期的推进,甜菜叶片净光合速率呈先升高后降低的趋势,CK处理的净光合速率在整个生育时期均显著高于ALK处理,说明盐碱胁迫使甜菜叶片净光合速率显著降低。随着施氮量的降低,甜菜叶片净光合速率也逐渐降低,除7月11日外,N180和N162处理均显著高于其他减氮处理;除8月5日外,N108处理甜菜的净光合速率在其余各取样时期均低于ALK处理。在6月22日,N180、N162和N144处理甜菜叶片的净光合速率显著高于ALK处理。各处理甜菜的净光合速率均在7月11日达到峰值,此时各处理间净光合速率的变化趋势与6月22日基本相同。
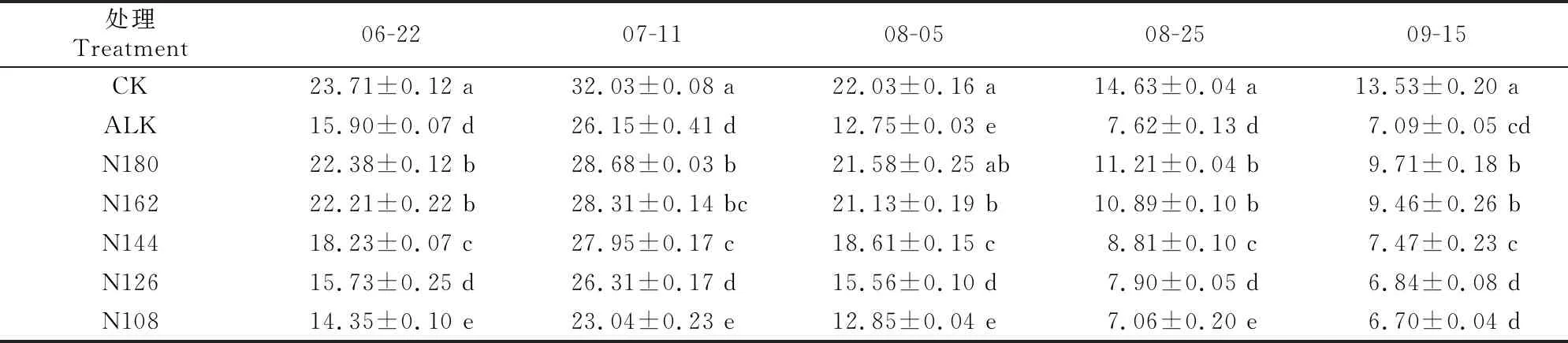
表3 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片净光合速率的影响
2.3.2 气孔导度 由表4可知,甜菜叶片气孔导度随甜菜的生长呈先升高后降低的趋势,在整个生育期内ALK处理均显著低于CK处理,在各取样时期较CK分别下降了28.5%(06-22)、21.0%(07-11)、40.3%(08-05)、31.2%(08-25)和43.6%(09-15),说明甜菜叶片气孔导度在盐碱胁迫下显著下降。在各取样时期,N180、N162和N144处理甜菜叶片气孔导度均显著高于ALK处理;随氮肥施用量的减少,气孔导度逐渐降低,其中N180处理显著高于N162、N144、N126和N108处理。
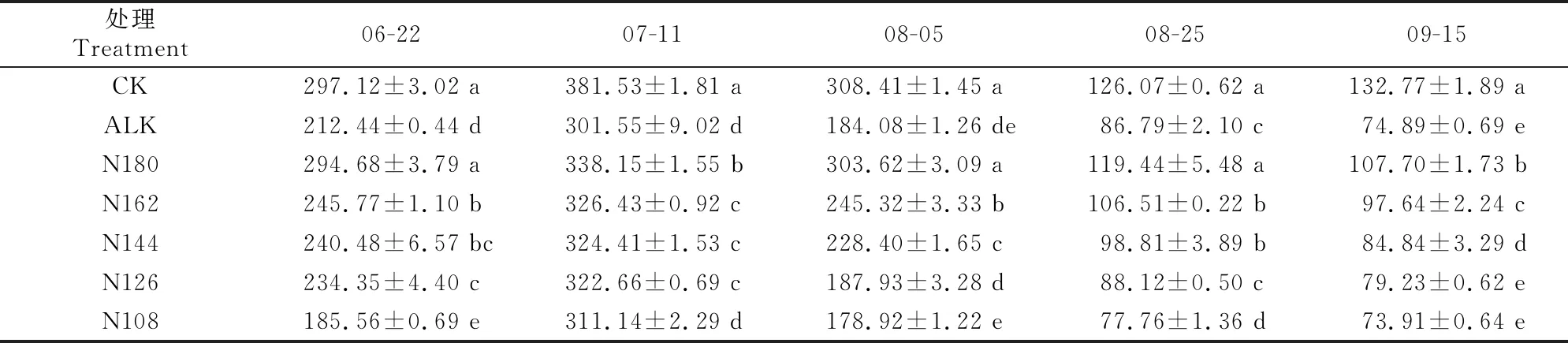
表4 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片气孔导度的影响
2.4 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片叶绿素荧光参数的影响
2.4.1Fv/Fm由表5可知,甜菜的Fv/Fm随生育期的推进呈无规律变化,N180,N162和CK处理在整个生育期均显著高于ALK处理,说明盐碱胁迫显著减低了甜菜的Fv/Fm,生物炭的加入显著增加了甜菜叶片的Fv/Fm。总体上而言,随着氮肥施用量的减少,甜菜叶片Fv/Fm呈逐渐下降趋势,除6月22日外,其余取样时期N180与N162处理相比无显著差异。
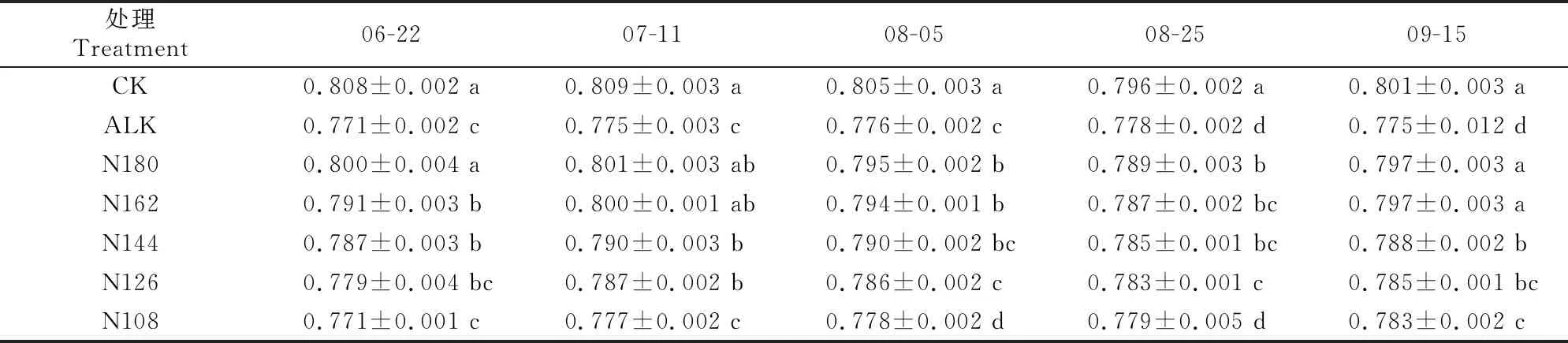
表5 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜Fv/Fm的影响
2.4.2Y(Ⅱ) 由图1可知,随着生育期的推进,各处理甜菜叶片的Y(Ⅱ)均呈先升高后下降的趋势,ALK处理的Y(Ⅱ)在各取样时期较CK处理分别下降了4.7%(06-22),2.0%(07-11),7.1%(08-05),2.7%(08-25)和4.1%(09-15),且差异均达到显著水平,说明盐碱胁迫使甜菜的Y(Ⅱ)显著降低。N180和N162处理甜菜的Y(Ⅱ)在各取样时期均显著高于ALK处理。
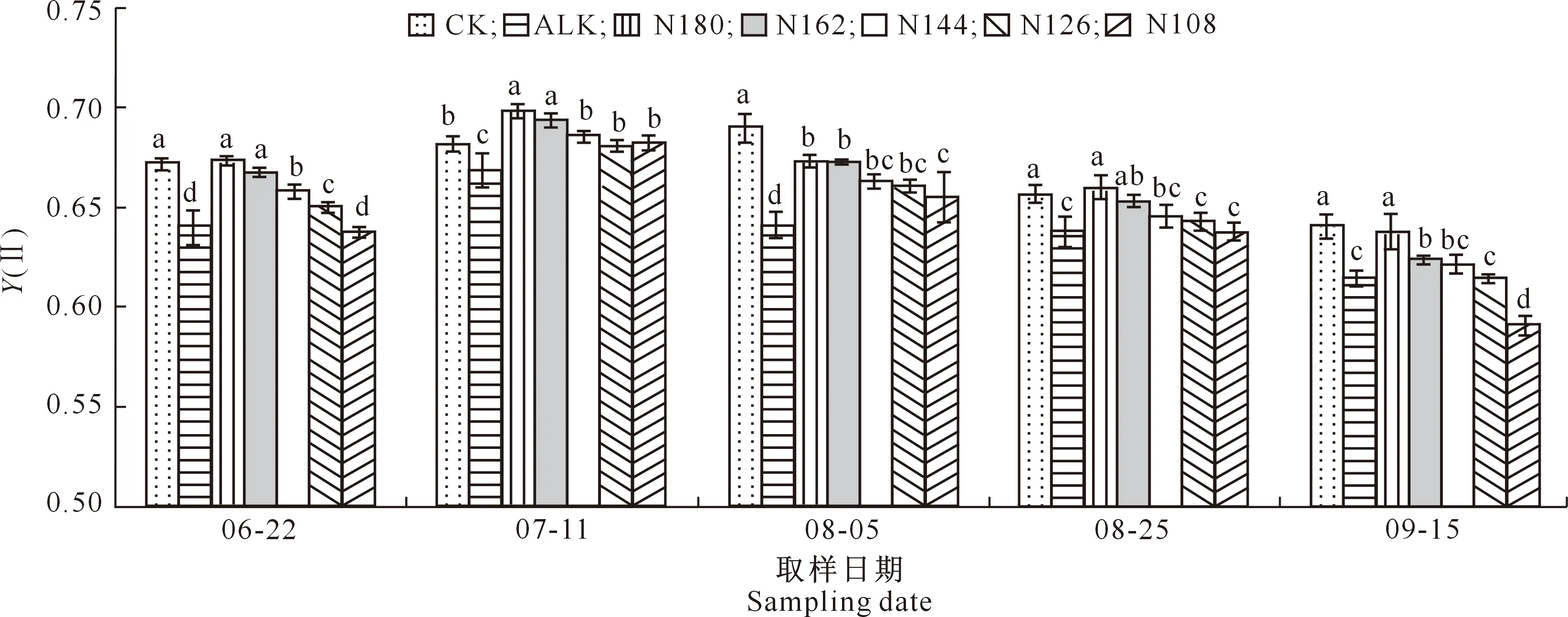
同一取样日期下不同小写字母表示各处理间在0.05水平差异显著。下同
2.4.3 ETR 由表6可知,随生育期的推进,各处理甜菜的ETR总体均呈先升高后降低的趋势。总体而言,盐碱胁迫使甜菜ETR明显降低,适量氮肥条件下施加生物炭可以显著提升甜菜的ETR。6月22日,N180和N162处理甜菜的ETR显著高于除CK外的其他处理。7月11日,各处理甜菜的ETR均达到最大值,此时N180和N162处理显著高于其他处理。8月5日,CK处理甜菜的ETR显著高于其他处理。至8月25日,N180处理甜菜的ETR显著高于其他处理。
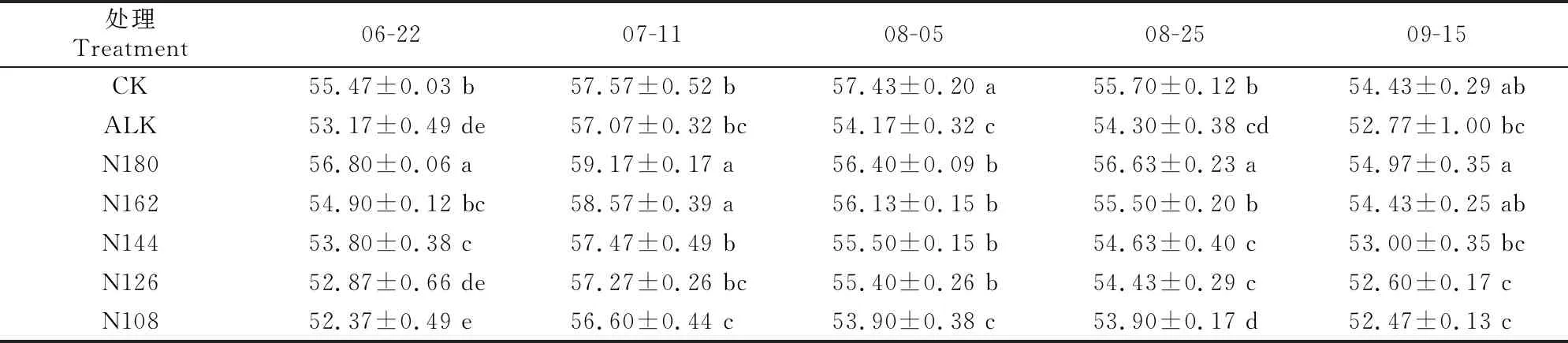
表6 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片ETR的影响
2.4.4qL表7显示,盐碱胁迫下加入生物炭后,随着生育期的推进,各处理甜菜的qL均呈先升高后降低的趋势,在各取样时期ALK处理的qL较CK处理分别降低了14.6% (06-22),14.6%(07-11),5.2% (08-05),10.1% (08-25) 和14.3% (09-15),且差异均达到了显著水平。随着氮肥施用量的减少,甜菜qL逐渐降低。N162和N180处理甜菜的qL在6月22日、7月11日和8月5日无显著差异,而在8月25日和9月15日,以N180处理显著高于N162。

表7 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜qL的影响
2.4.5 NPQ 从表8可以看出,随生育期的推进,各处理甜菜的NPQ均呈先减小后增大的趋势,盐碱胁迫显著增加了甜菜的NPQ,施加生物炭能显著降低NPQ。CK处理与N180处理、N180处理与N162处理相比,甜菜NPQ在各取样时期均无显著差异。在7月11日,除CK处理外其他处理的NPQ均达到最低值,此时ALK和N126处理甜菜的NPQ显著高于其他处理,随后又开始逐渐升高;8月25日,ALK、N144、N126和N108处理甜菜的NPQ显著高于CK、N180和N162处理;至9月15日,N144、N126和N108处理甜菜的NPQ显著高于其他处理。
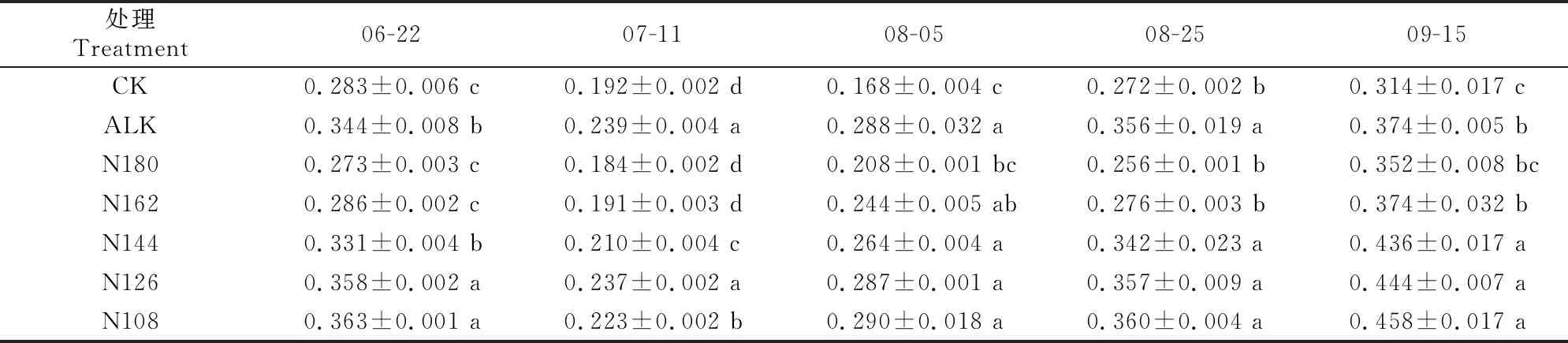
表8 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜NPQ的影响
2.5 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜干物质量的影响
2.5.1 叶片干物质量 由图2可知,各处理甜菜叶片干物质量总体均随生育期的推进呈先增加后减小趋势。盐碱胁迫显著降低了甜菜叶片的干物质量,ALK处理甜菜叶片干物质量在各取样时期较CK处理分别降低了67.5%(06-22),22.3%(07-11),26.1%(08-05),26.1%(08-25)和15.7%(09-15)。N180和N162处理甜菜叶片干物质量在各生育时期也显著高于ALK处理。除N108处理外,其他处理甜菜叶片的干物质量均在8月5日达到最大值,其中以N180处理最大,为11.14 g,较ALK处理高出177%。在8月25日,N108处理叶片干物质量较8月5日略有增长,其他处理则开始下降。

图2 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜叶片干物质量的影响
2.5.2 根系干物质量 由图3可知,随生育期的推进,各处理甜菜根系干物质量均呈逐渐增大趋势,盐碱胁迫显著降低了甜菜根系的干物质量。在各取样时期,N180、N162、N144处理甜菜根系干物质量均显著高于ALK处理;在6月22日、8月5日和8月25日,N180处理甜菜的根系干物质量均显著高于N162处理;在7月11日,N180和N162处理甜菜的根系干物质量无显著差异,而在9月15日,N162处理甜菜的根系干物质量较N180处理高7.2%,且差异显著。
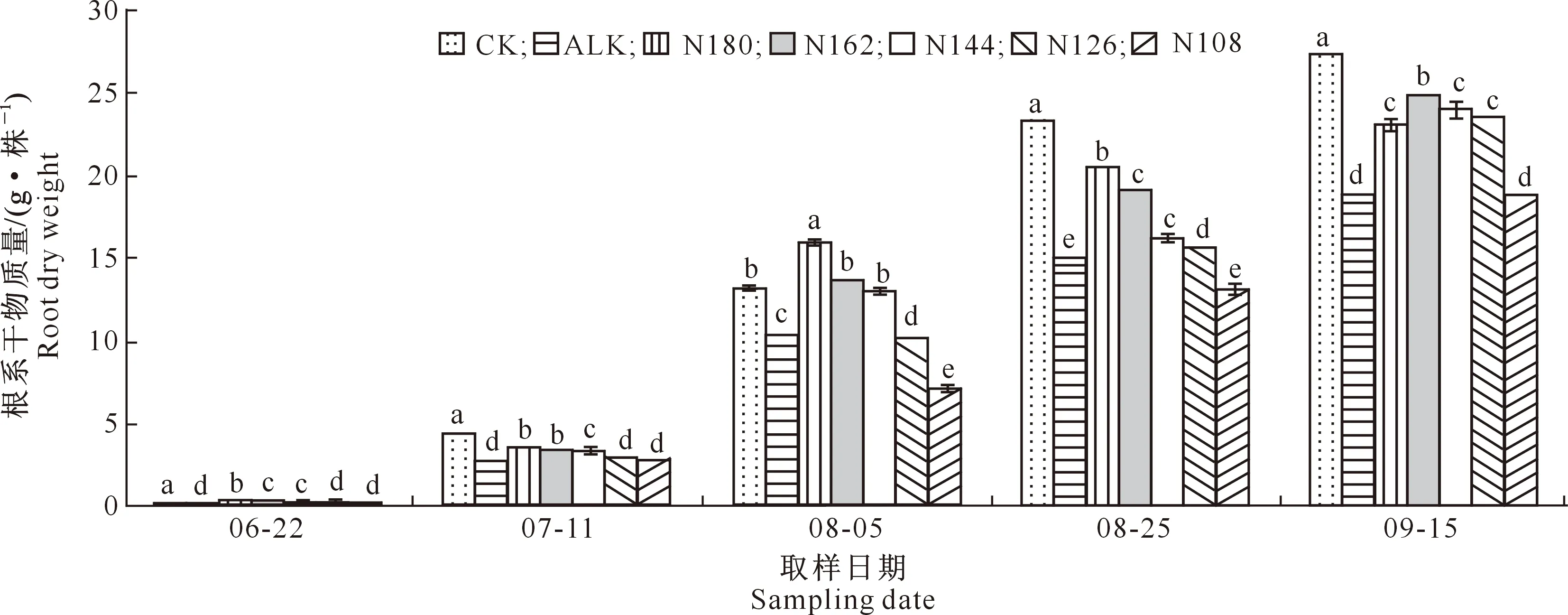
图3 盐碱胁迫下施加生物炭和减少氮肥用量对甜菜根系干物质量的影响
2.6 甜菜叶片干质量与光合重要指标的相关性
在8月5日,除N108处理外,其他处理甜菜的叶片干物质量均达到最大值,说明此时甜菜处于叶丛快速生长期,因此选择8月5日时的各指标参数进行相关分析。叶绿素含量和RuBP羧化酶活性均与甜菜的光合作用相关,甜菜叶片净光合速率、Y(Ⅱ)和qL是光合作用速率与效率的直观表现。由表9可以看出,甜菜叶片的叶绿素含量、RuBP羧化酶活性、净光合速率、Y(Ⅱ)、qL和叶片干物质量之间均存在极显著的正相关关系。

表9 甜菜叶片干物质量与光合重要指标的相关性
3 讨论与结论
光合作用是植物最基本也是最重要的生理代谢之一,其能制造植物所需的有机物,平衡大气中氧和二氧化碳的平衡,在自然界中具有重要意义。盐碱土中过量的Na+导致土壤水势降低,使植物从土壤中吸水困难;植物吸收大量的Na+后不仅使植物对其他养分难于吸收,而且还会导致植物的生理代谢紊乱,降低植物的光合作用。研究表明,高浓度的盐碱胁迫显著降低了植物叶片的叶绿素含量和光系统Ⅱ的光化学效率[14-17],这与本研究结果相一致,其原因可能是过量的盐分导致植物原生质凝聚,破坏了叶绿体的结构,抑制了光合作用的光反应[18]。在盐碱胁迫下枸杞的RuBP羧化酶活性显著降低,使得枸杞的碳同化能力下降,光合作用速率减慢[19],与本研究结果相同,这可能是由于能量供应被限制所致[20]。李帅霖等[21]研究表明,施加2%的生物炭和0.2 g/kg纯N的氮肥可使小麦叶片净光合速率显著提高。其原因可能是由于生物炭疏松多孔的结构能提高土壤的透气性,并使土壤的比表面积增大,有利于植物更容易地从土壤中交换营养物质,为植物提供了良好的生长环境。
本研究结果表明,盐碱胁迫能显著降低甜菜的叶绿素含量,减慢光合作用光反应和暗反应速率及干物质积累速率,而施加生物炭能有效缓解盐碱胁迫对甜菜生长的影响,这与武沛然等[22]关于盐碱胁迫下施加生物炭对甜菜生长的影响研究结果相一致。同时,在减少10%氮肥施用量 (N162) 的情况下,甜菜的光合作用和干物质量与氮肥不减量处理(N180)无显著差异,这可能是由于施加生物炭和适当减少氮肥用量增加了土壤微生物的多样性。有研究表明,生物炭与氮肥配施能使土壤中的微生物生物量碳提高45.1%~208.6%,土壤脲酶活性提高91.7%,还能显著增加土壤微生物数量[23]。这可能是由于生物炭具有较强的吸附性,能吸附土壤中的营养元素和矿质元素[24],有利于土壤中微生物的生长繁殖;土壤脲酶活性的提高和微生物多样性的增加能改善土壤结构,促进养分循环,从而提高了肥料在盐碱土中的利用效率[25],间接提高甜菜的光合作用速率和干物质积累速率。
综上所述,施加3%生物炭能有效缓解盐碱胁迫对甜菜光合作用的抑制。结合施肥成本考虑,在盐碱胁迫条件下施加3%生物炭的同时减少10%氮肥 (N162) 最为合理,但关于盐碱胁迫下施加生物炭和减少氮肥用量对甜菜生长和土壤理化性质的影响及其机理仍需进一步研究。
猜你喜欢
金桥(2021年11期)2021-11-20 06:37:14
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
现代畜牧科技(2021年4期)2021-07-21 06:12:54
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
人民黄河(2020年12期)2020-12-30 12:55:30
山西农业科学(2020年9期)2020-09-14 08:20:08
山西农业科学(2019年12期)2019-12-19 04:03:30
电子制作(2017年10期)2017-04-18 07:23:14
中国糖料(2016年1期)2016-12-01 06:49:04
西藏科技(2016年8期)2016-09-26 09:00:55
