
热处理羊乳酪蛋白结构与抗原性的变化规律
2021-03-02 14:37:24薛海燕樊娇娇贺宝元操歌李晶莹宋宏新
现代食品科技 2021年2期
薛海燕,樊娇娇,贺宝元,操歌,李晶莹,宋宏新
(1.陕西科技大学食品与生物工程学院,陕西西安 710021)
(2.陕西科技大学轻工科学与工程学院,陕西西安 710021)
乳制品含有丰富的蛋白质、脂肪、维生素、矿物质等的营养成分,是人类摄取营养的重要来源之一。酪蛋白是乳中含量最高的蛋白质,约占乳总蛋白质的80%,它在乳中主要以胶束形式存在,是一种高度磷酸化钙结合蛋白[1],酪蛋白主要有αs1-CN,αs2-CN,β-CN和κ-CN四种类型。对于过敏人群来说,酪蛋白则是引起人体过敏反应的主要过敏原之一,如表1中,羊乳酪蛋白中β-CN含量最大,κ-CN也较牛乳含量高,而αs1-CN含量低于牛乳。有研究显示了对低αs1-CN含量的山羊乳有较小免疫反应性[2],故αs-CN对研究羊乳抗原性等方面性质极其重要。而羊乳与牛乳酪蛋白结构不同,受加工影响的结构和抗原性变化亦不同。
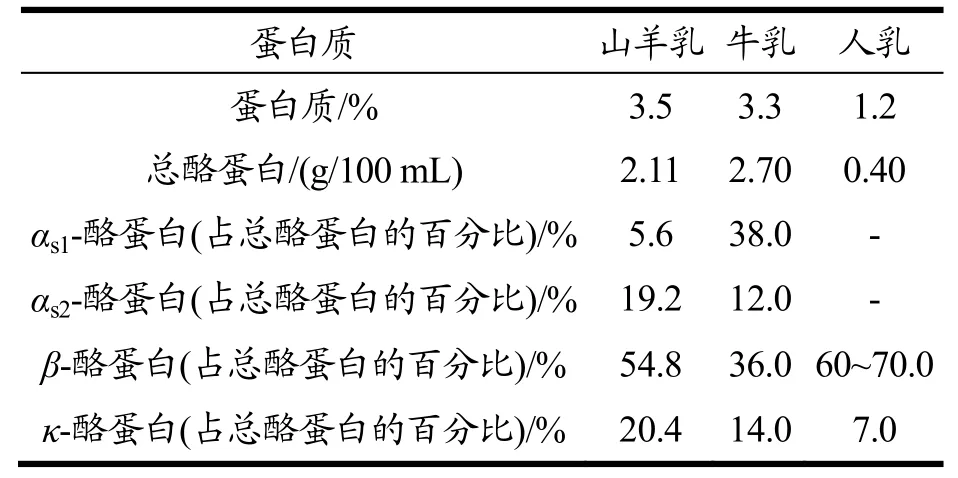
表1 山羊乳、牛乳和人乳中酪蛋白含量差异[3]Table 1 Difference of casein content in goat milk, bovine milk and human milk
乳品加工中,通过一定的加工处理可使蛋白质改性,引起乳品蛋白质的分子结构及理化性质发生改变,有可能会改变乳品的过敏性[4]。主要的处理方法有物理法(热处理[5]、高压处理、辐照处理等)、化学法(糖基化作用等)、酶法和生物法等。许倩[6]等利用动态高压微射流结合加热处理浓缩乳蛋白可降低其中酪蛋白的抗原性,增加α-乳白蛋白的抗原性,但对β-乳球蛋白效果不明显;张微等[7]研究蛋白酶水解作用对牛乳酪蛋白抗原性的影响,结果表明胰蛋白酶能够有效降低牛乳酪蛋白的抗原性。Li[8]等研究发现,经超声处理后的虾肉蛋白,纯化虾原肌球蛋白的IgE结合能力降低了80%。麻小娟[9]对卵白蛋白采用糖基化处理来探索其抗原性及过敏原性,发现随着糖基化的进行,卵白蛋白的抗原性逐渐升高,过敏原性逐渐降低。
而热处理是乳品加工中常用的物理改性方法。在巴氏灭菌奶、UHT牛奶、乳奶、炼乳等乳制品的加工过程中都有涉及加热。热处理能够使蛋白分子结构发生变化,使得其抗原表位被破坏、掩埋、暴露等,最终影响蛋白的抗原性。Bu[10,11]等通过不同温度加热处理乳清分离蛋白,结果发现当加热温度在90 ℃以下时,α-乳白蛋白和β-乳球蛋白的抗原性增加,当温度高于90 ℃时,两种蛋白的抗原性均显著下降。张银[12]等采用热处理与动态超高压加工两种方式结合处理卵白蛋白,结果表明卵白蛋白抗原性随着加热温度的升高而降低,随着压力的升高先降低后回升,并且表明了热处理对卵白蛋白抗原性的影响比超高压大。
关于热处理改变蛋白性质,目前大多是关于蛋白结构变化的研究,在牛乳方面较多关于乳清蛋白中α-La与β-Lg的研究,但关于对羊乳酪蛋白结构及抗原性等的研究尚少。因此,在现有的研究基础上,采用羊乳酪蛋白中的α-CN与β-CN作为研究对象,对其进行不同温度处理,测定其蛋白的结构变化与抗原性变化,并分析不同温度下蛋白的结构变化与抗原性之间的关系,为羊乳的过敏原性研究奠定基础。
1 材料与方法
1.1 原料与试剂
羊乳:陕西金牛乳液有限公司提供;Tris-base;邻甲苯甲醛(DNPH);乙酸乙酯;盐酸胍;乙二胺四乙酸(EDTA);5,5’-二硫代双(2-硝基苯甲酸)(DTNB);1-苯氨基萘-8-磺酸(ANS);胰蛋白酶;胃蛋白酶。
1.2 仪器与设备
超高温高压灭菌,上海申安医疗器械厂;电泳仪,北京百晶生物技术有限公司;PHS-3C型pH计,赛多利斯科学仪器有限公司;高速冷冻离心机,安徽中科中佳科学仪器有限公司;凝胶成像分析系统,南京驰顺科技有限公司;冷冻干燥机,青岛永和创新电子科技有限公司;全波长扫描式多功能读数仪,赛默飞世尔科技有限公司;圆二色光谱仪,英国应用光物理公司。
1.3 方法
1.3.1 羊乳酪蛋白的分离制备
将新鲜羊乳离心脱脂,盐酸沉淀(pH=4.1)羊乳酪蛋白。用0.05 mol/L的NaOH作为溶剂,配置30 mg/mL羊乳酪蛋白溶液,在30 ℃下调pH至12.0,加CaCl2溶液至体系中的Ca2+浓度为0.09 mol/L,调pH至中性,4000 r/min离心15 min,收集沉淀①。将沉淀①于3 mol/L的尿素溶液中完全溶解后,调pH为4.1,离心,收集沉淀②,反复水洗沉淀,透析并冻干得α-CN;将沉淀①溶于水,待完全溶解后调pH 4.1,在4 ℃下冷却3 h,离心,提取上清液,在30 ℃下调pH为4.1,再离心,收集沉淀③,水洗3次,透析并冻干得β-CN。将制得酪蛋白的分离组分在冷冻干燥机中,进行真空冻干,冻干粉末-20 ℃保存备用。
1.3.2 羊乳酪蛋白的热处理
将羊α-CN、羊β-CN用0.01 mol/L pH 7.4的磷酸盐缓冲溶液(PBS)配成1 mg/mL的溶液,依据常见的处理方式温度设定如下:63 ℃(低温保持巴氏杀菌)30 min、80 ℃(高温巴氏杀菌)15 s、100 ℃(煮沸杀菌)300 s、134 ℃(超高温高压灭菌)4 s,不进行热处理作为对照组。加热处理后的样品,取出后瞬间低温冷却放置。
1.3.3 SDS-PAGE检测分子量[13]
将热处理后的酪蛋白进行SDS-PAGE(聚丙烯酰胺凝胶电泳)分析热处理对蛋白质分子量的影响。
1.3.4 酪蛋白的侧链基团修饰变化
1.3.4.1 游离羰基含量的测定[14]
采用DNPH比色法测定热处理后的羊α-CN、羊β-CN样品溶液中游离羰基含量。以不加蛋白样品为空白,在370 nm波长处测量上清液的吸光值,用Bradford法测上清液中的蛋白质含量[15]。按朗伯比尔定律计算羰基含量。
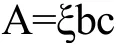
式中:A表示所测样品的净吸光值;ξ为DNPH的摩尔消光系数,值为22000 L/(mol·cm);b为比色皿的厚度;c为游离羰基的浓度,用nmol/g蛋白质表示。
1.3.4.2 自由巯基的测定
参照Ellman’s DTNB法[16]。分别取得热处理前后的1.0 mL的羊α-CN、羊β-CN样品溶液与4.0 mL Tris-Gly缓冲溶液(0.086 M Tris、0.09 M Gly、5 mM EDTA、pH 8.0)混匀,再加入50 μL 4 mg/mL的Ellman’s试剂(4.0 mg DTNB溶于1.0 mL的Tris-Gly缓冲液),37 ℃反应15 min,在412 nm处测其吸光值。
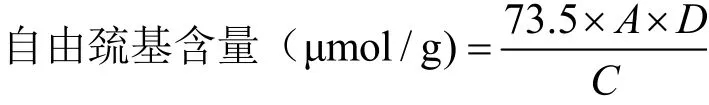
式中:A为吸光度;D为稀释倍数;C为样品浓度(mg/mL)。
1.3.5 蛋白质二级结构的测定
利用圆二色光谱仪对热处理前后的羊α-CN、羊β-CN样品用PBS配置成0.02 mg/mL溶液进行色谱检测。测定条件:扫描范围是210~280 nm,其中石英样品池的光程是0.01 cm,扫描速度是100 nm/min,扫描带宽为0.5 nm,步长为1 nm。通过系统自带的分析软件对结果进行分析。
1.3.6 表面疏水性的测定
采用ANS荧光探针法对热处理前后的羊α-CN、羊β-CN的疏水性进行测定[17]。测定条件为:激发波长为370 nm,扫描发射波长范围为400~600 nm,电压为400 V,狭缝宽度均为10 nm。本实验用荧光强度来表示疏水性的大小。
1.3.7 测定消化后酪蛋白抗原性
模拟婴儿胃肠消化,将热处理后的羊α-CN、羊β-CN调节pH至3,在37 ℃下加入0.3%(质量体积比)的胃蛋白酶,恒温搅拌反应2 h,即开始进行模拟胃液消化。反应停止后立即调节溶液pH至7,加入胰蛋白酶使其浓度为10 mg/mL,恒温搅拌反应4 h,即进行模拟肠液消化。最后用100 ℃沸水浴灭酶5 min停止反应,然后放置冰水冷却,将样品分装后-20 ℃条件下保存待测[18]。
1.3.8 抗原性评估[19]
采用间接竞争ELISA方法测定被处理的α-CN和β-CN的残余抗原性。在间接竞争ELISA体系中待测抗原与包被抗原竞争结合抗体血清,因此被测抗原的抗原性大小与吸光值成反比。羊α-CN、羊β-CN稀释包被浓度为12.5 μg/mL,取100 μL包被在96酶标孔板中4 ℃过夜;隔天洗板后加入待测抗原与羊抗兔IgG预混液进行竞争反应,其中对照组不加待测抗原;1 h后洗板并加入1:4000稀释的羊抗兔IgG-HRP标记物进行检测并显色,终止显色后检测吸光值OD,将梯度浓度对应的OD值转化为B/B0值作为纵坐标(B0为无抗原竞争时的OD值,B为各相应浓度抗原抑制时的OD值),相应浓度的对数是横坐标lg[α-CN](或lg[β-CN]),绘制标准曲线,并进行线性回归拟合,由此计算得待测抗原的浓度,来反映相应抗原性的变化。
1.3.9 数据处理
采用Origin 9.0软件作图,并用SPSS 19软件进行统计学分析。
2 结果与讨论
2.1 分子量分析
如图1为羊乳分离制备的酪蛋白组分电泳图谱,经凝胶成像灰度值分析估算出所分离得到的羊α-CN和β-CN的纯度都在80%以上,市售标准蛋白纯度为70%以上,表明所分离的蛋白较纯,可用于后续研究。
酪蛋白经过热处理、高压脉冲电场、辐射以及超声波手段的处理后,其分子量可能会发生改变。图2为羊乳的α-CN和β-CN经过不同温度处理后对酪蛋白分子量的影响。观察各组条带颜色可知:羊乳的α-CN和β-CN在63 ℃时灰度无明显变化,在温度升高达到80 ℃之后有灰度变浅的现象。电泳图谱中蛋白条带的灰度深浅可以代表进入电泳凝胶内的蛋白含量变化。因为羊乳的酪蛋白热稳定性差,使得酪蛋白发生脱磷酸化反应引起蛋白分子交联形成大分子物质,而无法进入分离胶形成条带[20],导致电泳图所示的条带灰度变浅。这与李林强等[21]对于热处理牛羊乳酪蛋白的SDS电泳结果相似,蛋白经一定程度的热处理均发生交联聚合,且羊乳较牛乳更易发生聚合。
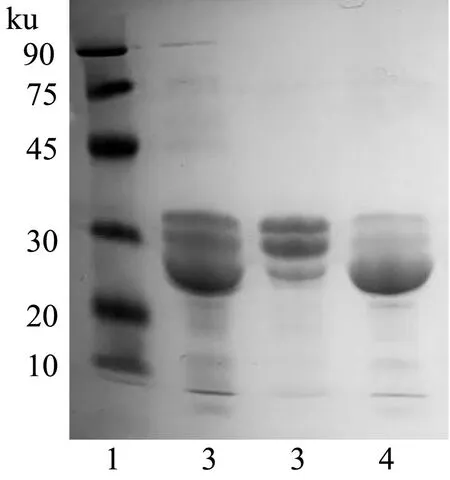
图1 羊奶酪蛋白成分的SDS-PAGEFig.1 SDS-PAGE of the components of casein in goat milk

图2 温度对羊乳中α-CN和β-CN分子量的影响Fig2. Effect of temperature on the molecular weight of α-CN and β-CN in goat milk
2.2 酪蛋白的侧链修饰的变化
2.2.1 游离羰基含量
蛋白质的很多侧链氨基酸官能团都容易氧化形成羰基类衍生物,如肽键的断裂、氨基酸侧链的氧化等,因此游离羰基含量可以被用来判断蛋白质的氧化程度[22-24]。图2为不同温度的处理下,羊乳α-CN和β-CN中游离羰基含量的变化。可以观察出,在温度达到63 ℃时,羊乳α-CN和β-CN的游离羰基含量显著增长,且随着温度的升高不断增多,当温度到达134 ℃时,羊乳的α-CN和β-CN中游离羰基含量与未处理时相比分别增加了134.72%和110.98%,均有极显著增长。这是由于蛋白中的α-CN和β-CN为无规则卷曲蛋白,蛋白中的色氨酸、蛋氨酸、组氨酸等受高温影响最易被氧化形成羰基[25]。羊乳热稳定性低,易在高温条件下被氧化,且温度越高,氧化程度越剧烈。
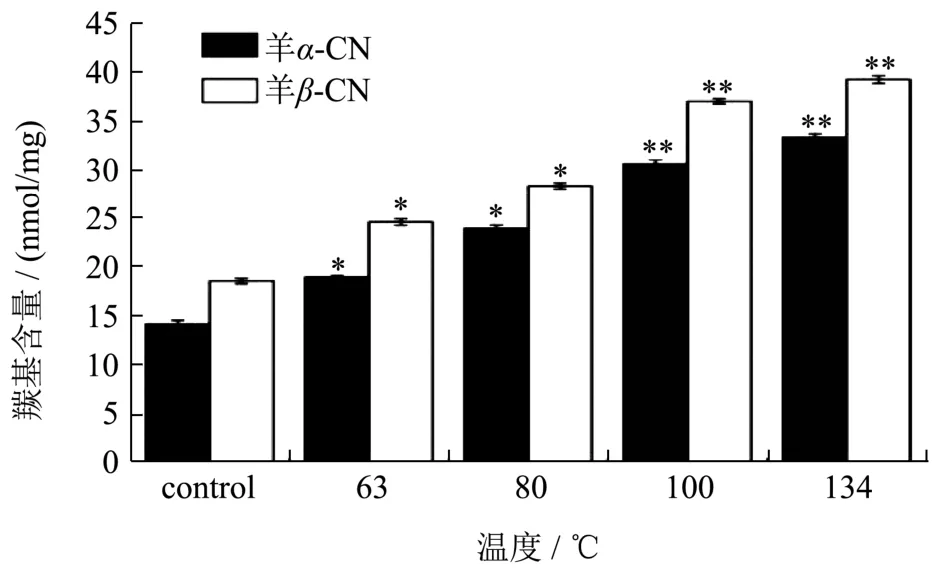
图2 温度对羊乳的α-CN和β-CN中游离羰基含量的影响Fig.2 Effect of temperature on free carbonyl content of α-CN and β-CN in goat milk
2.2.2 自由巯基含量
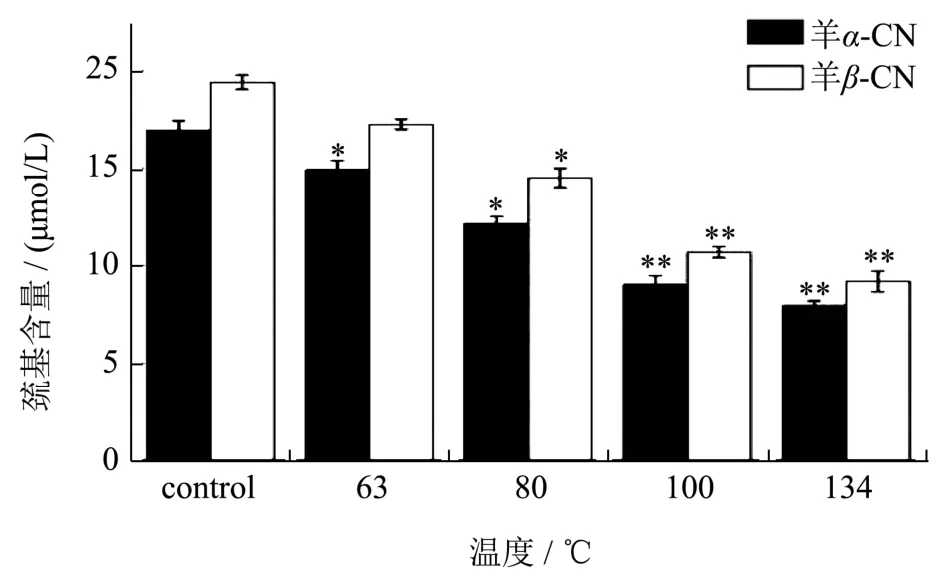
图3 温度对羊乳的α-CN和β-CN中自由巯基含量的影响Fig.3 Effect of temperature on free sulfhydryl content of α-CN and β-CN in goat milk
巯基基团时蛋白质中的重要功能性基团之一,它可以相互交联后形成二硫键,用来稳定蛋白质的高级结构[26,27]。图3是不同温度的处理下,羊乳的α-CN和β-CN中自由巯基含量的变化。由图可知,羊乳的α-CN中自由巯基含量低于β-CN中自由巯基的含量,通过计算得出,当温度加热至63 ℃时,羊乳α-CN中的自由巯基含量有显著下降,在温度达到100 ℃时便有极显著降低,随后下降趋于平缓,当温度为134 ℃时,其自由巯基含量减少了53.03%。然而,羊乳β-CN中的自由巯基含量不及α-CN对温度敏感,当温度达到80 ℃时,自由巯基的含量才开始有了显著降低,当温度到达134 ℃时,自由巯基含量减少了52.65%,与未处理时相比,有了极显著差异。
2.3 酪蛋白二级结构的变化
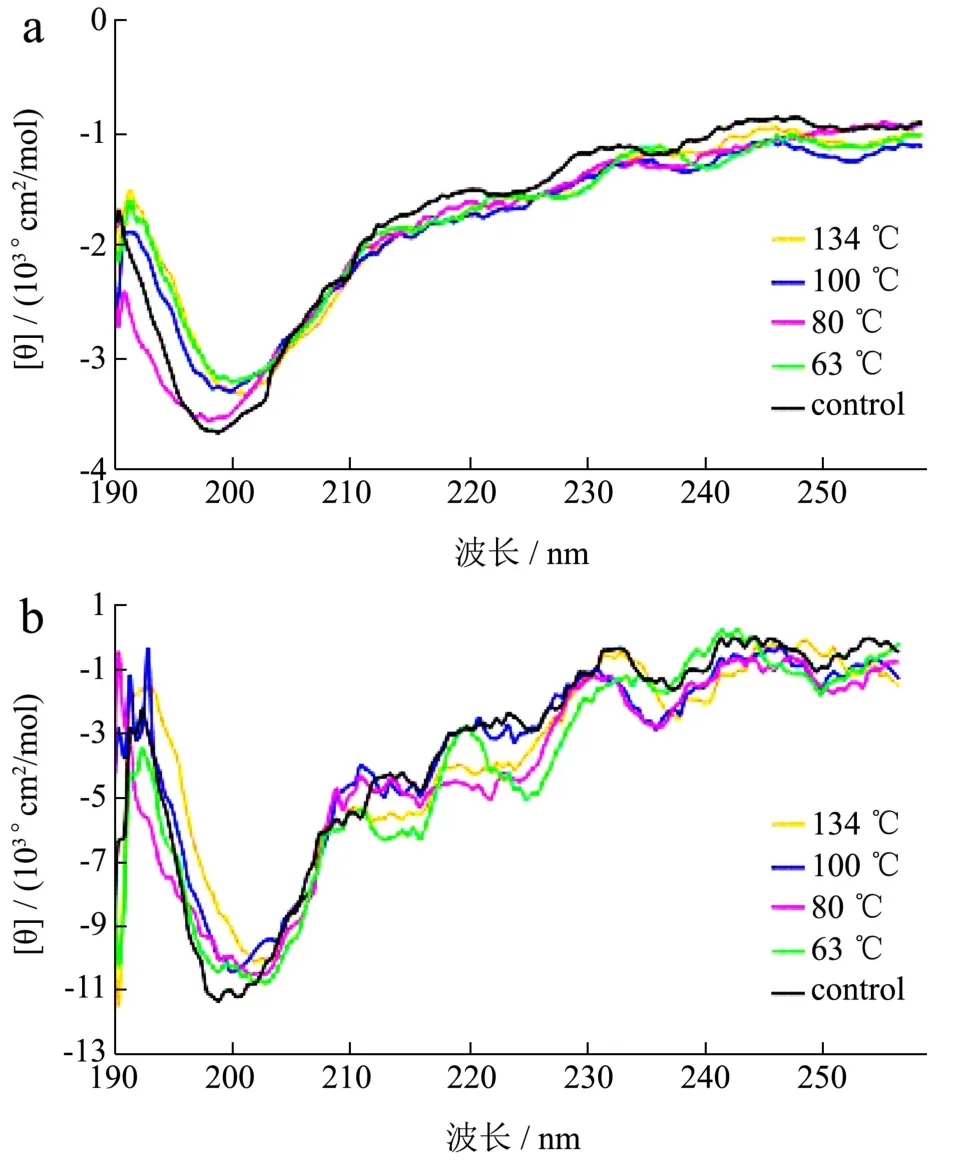
图4 不同温度条件下羊乳α-CN(a)和β-CN(b)的圆二色谱图Fig.4 CD chromatogram of α-CN (a) and β-CN (b) from goat under the different thermal processing conditions
蛋白的二级结构中,肽键的序列都是一定的,这些顺序决定着肽键能级会发生的跃迁情况,圆二色谱的谱带因蛋白的二级结构不同而有所差异,因此,根据圆二色谱可以分析蛋白的二级结构差异。如图4为不同温度条件下羊乳α-CN和β-CN的圆二色谱图。由图4可以看出,在190~250 nm范围内,在206 nm处由负转正有一正谱带,这是β-转角结构[28];在212 nm处和225 nm处的两个负峰是α-螺旋特征峰;在222 nm处还有一个负谱带,是β-折叠特征峰,谱图可发现结构中含有大量无规则卷曲结构[29]。
表2和表3是关于羊乳中α-CN和β-CN的二级结构含量变化。计算发现羊乳α-CN与β-CN与的二级结构含量变化,其中,羊β-CN的α-螺旋结构的初始含量高于α-CN的该结构含量,这或许是由于羊乳本身α-CN的含量就远低于β-CN[30]。且由表格可知,随着温度变化,两种酪蛋白的无规则卷曲结构和β-转角的含量变化均随着温度的升高而减少,当温度达到134 ℃时α-CN的β-转角结构含量减少了85.38%,无规则卷曲减少了51.71%,β-CN的两类结构的含量分别减少了56.14%和94.38%,这表明了随着温度升高羊乳α-CN的结构弹性增加,这或将与它更易被水解有关。α-螺旋结构的含量变化则与上述两种结构变化相反,随着温度的升高而增加,当温度达到134 ℃时两种蛋白分别增加了53.11%和85.50%。该二级结构变化与章宇斌[31]对酪蛋白进行不同温度处理后的CD测定的部分研究结果相似,加热导致αs-及β-CN的螺旋含量逐渐增加。然而,两种酪蛋白的β-折叠结构的含量却随着温度的升高分别在26.52%~28.56%和13.79%~ 21.88%的区间内有着小幅度的无规则的变化,这表明了温度的改变不会对羊乳的该蛋白结构有明显的影响。吴相佚[26]等对浓缩乳蛋白70进行热处理,其蛋白二级结构中α-螺旋和β-转角的含量下降,β-折叠和无规则卷曲含量上升。蛋白的不同可能导致结构的差异,且乳蛋白自身还存在着酪蛋白与清蛋白的交联与聚集,可能导致结果的差异。
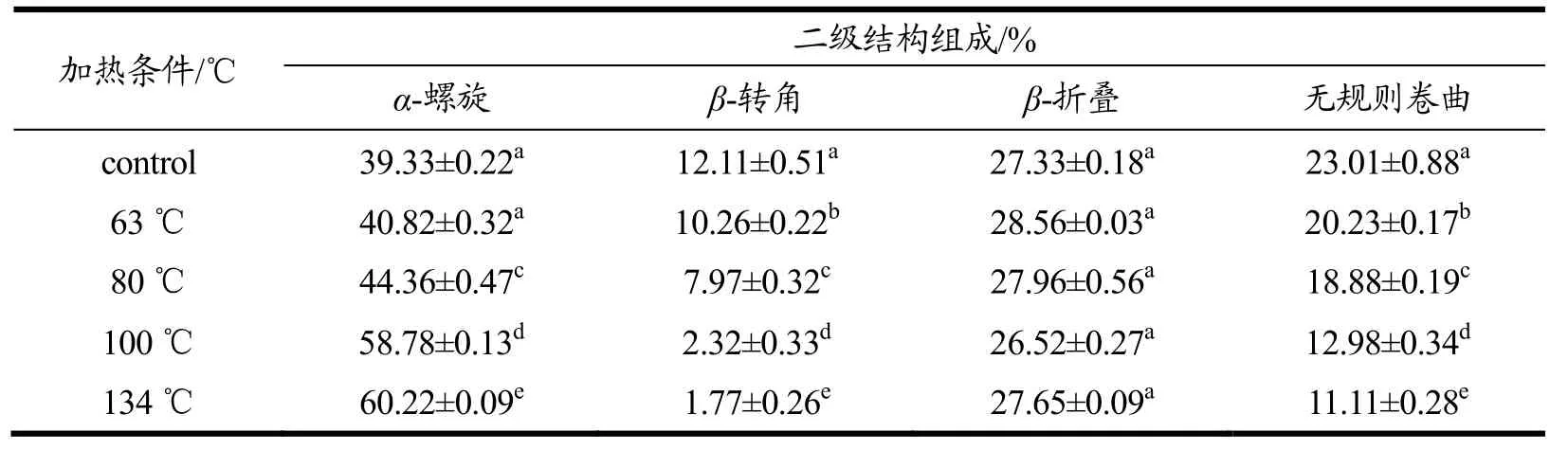
表2 温度对羊α-CN二级结构各组分含量的影响Table 2 Effect of temperature on the content of goat α-CN secondary structure (%)
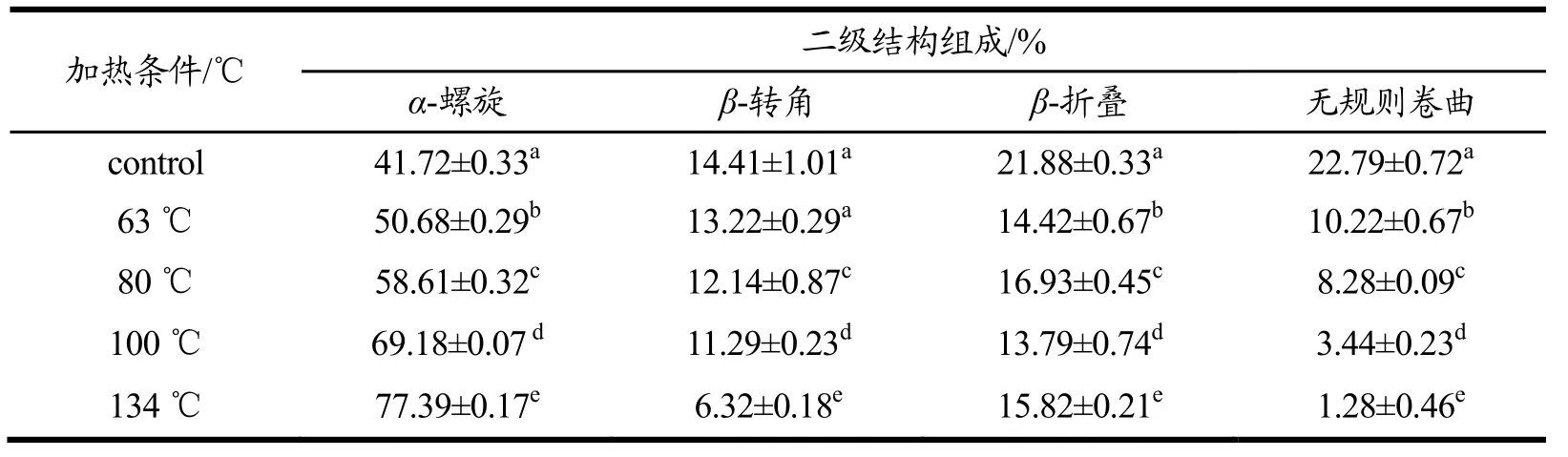
表3 温度对羊β-CN二级结构各组分含量的影响Table 3 Effect of temperature on the content of goat β-CN secondary structure (%)
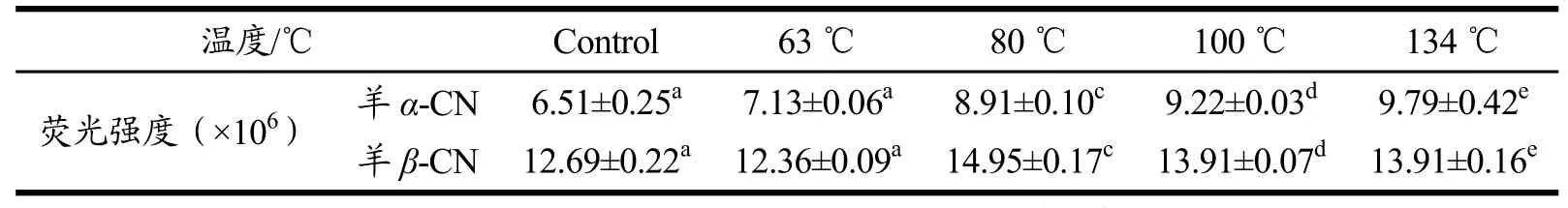
表4 温度对样品荧光光谱分析时最大荧光强度的影响Table 4 Effect of temperature on maximum fluorescence intensity in fluorescence spectrum analysis of samples
2.4 酪蛋白疏水性分析
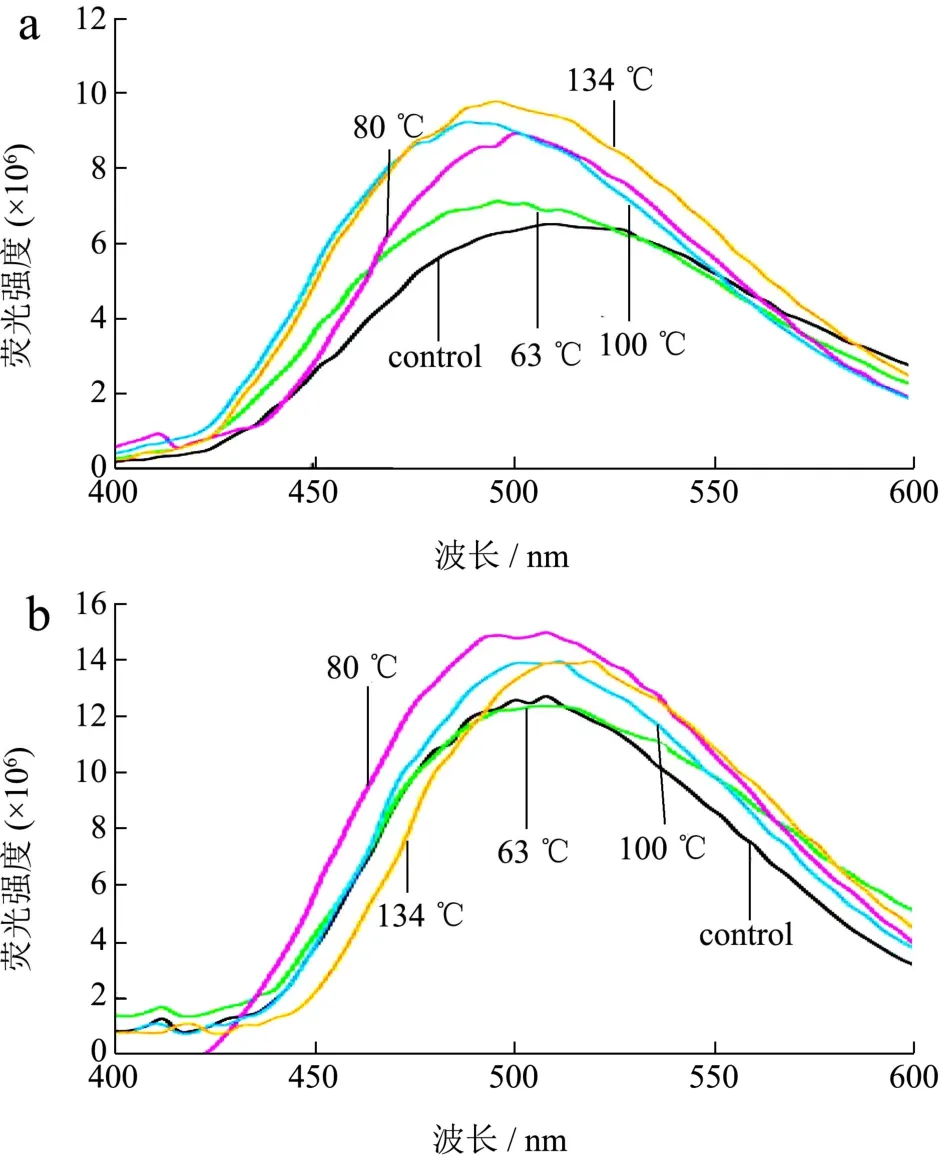
图5 不同温度条件下羊α-CN(a)和β-CN(b)的荧光光谱Fig.5 Fluorescence spectra of α-CN (a) and β-CN (b) from goat at different temperatures
图5是由ANS荧光探针检测法得到的两种酪蛋白不同温度条件下的荧光光谱图。应用荧光探针法来研究蛋白质的构象,在一定范围内,最大荧光强度与蛋白表面的疏水性呈线性关系[32]。
观察谱图可以看出,未经过热处理时,羊乳的α-CN疏水性均小于β-CN的疏水性,这说明羊乳α-CN的疏水性结合位点低于β-CN。结合表4可以看出,随着温度升高,羊乳α-CN的最大荧光强度随之增强,到134 ℃时增加了约50.38%,这是由于温度的升高导致二硫键介导的聚合反应隐藏了部分含有荧光色基团的氨基酸,导致荧光强度增强,同时,蛋白二级构象发生改变,一些集中在α-螺旋、β-折叠等转角处的亲水区域因结构的变化而减少,从而使得蛋白分子内的疏水基团暴露,也可导致荧光强度增强。上述现象可以证明随着温度的升高,酪蛋白结构表面疏水基团会增加,而疏水性的增强或将导致蛋白的解离程度有所增大。又因羊乳酪蛋白的热稳定性弱,且β-CN在羊乳酪蛋白中占有很大比例,所以羊乳β-CN的胶束与ANS结合的亲和力与紧密度相对不稳定,使得最大荧光强度与其他蛋白相比在温度的升高过程中有着摆动性的无规则变化。
2.5 抗原性的变化
通过模拟婴儿的胃肠水解消化,并采用间接竞争ELISA法检测热处理对牛羊乳α-CN和β-CN的抗原浓度变化,来反应其抗原性的变化。图6表示消化前后的抗原性变化。观察发现:在未经过热处理时,羊乳中的β-CN含量高于α-CN,因此图示羊乳α-CN的抗原性低于β-CN的抗原性[33]。在未经过胃肠模拟消化时,羊乳中α-CN和β-CN的抗原性均随着温度的升高而持续性降低,当加热温度为80 ℃时,抗原性有显著性变化,当温度为134 ℃时抗原性有了极显著的降低,α-CN和β-CN的抗原性分别降低了61.80%和69.02%。羊乳的α-CN在模拟胃肠消化之后,其抗原性与相同温度下未消化时的抗原性相比约下降了50%左右,且仅当温度达到63 ℃时抗原性就有显著降低,而羊乳β-CN的抗原性却直至温度达到80 ℃时才有显著性减少。当温度在134 ℃时,两种蛋白的抗原性浓度相似,仅剩7.86~8.54 μg/mL,与未经温度处理的消化前后相比,都有着极显著的降低。这就与前人所得出的羊乳酪蛋白较牛乳在体内来说更易被人体消化吸收的结论相呼应。结合羊乳的α-CN和β-CN的侧链修饰变化、二级结构和疏水性的变化可推断,随着温度升高,羊乳酪蛋白抗原性的减少,除了与其氧化程度(游离羰基含量的增多)、疏水性增加有关,或与二级结构中的β-转角含量和无规则卷曲含量的减少有关,它们随着温度的升高几近消失,这种现象或导致部分抗原结合位点的消失而减少了免疫反应的进行。
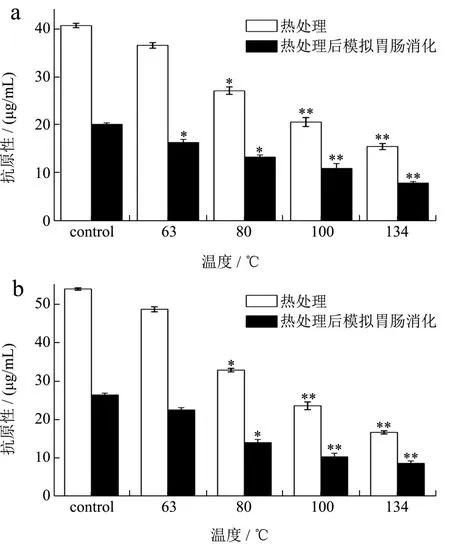
图6 不同温度对羊α-CN和β-CN抗原性的影响Fig6. Effects of different temperatures on the antigenicity of goat α-CN (a) and β-CN (b)
3 结论
热处理羊乳酪蛋白会发生蛋白的交联,导致部分沉降或聚集成大分子物质。同时,样品蛋白被高温所氧化,导致游离羰基含量升高、自由巯基含量减少。且随着温度的不断升高,羊乳的α-CN和β-CN的二级结构中,α-螺旋呈现增加趋势,β-转角和无规则卷曲则减少至几近消失。此外,羊乳α-CN的荧光强度均随着温度升高而增强,而羊乳β-CN的胶束与ANS结合的亲和力与紧密度相对不稳定,故羊乳β-CN的荧光强度呈现无规则变化。温度的升高会引起蛋白的卷曲程度及弹性结构发生改变,导致分子内的疏水基团,二硫键等暴露,蛋白结构发生聚合或交联,部分分子表面的抗原表位被掩埋或被氧化分解,最终导致了被测蛋白抗原性均有所降低。故温度的升高,能够使羊乳α-CN和β-CN的抗原性减弱,且与蛋白的游离羰基含量呈负相关关系,其二级结构中β-转角和无规则卷曲与抗原性变化呈一定正相关,而α-CN的抗原性与疏水性呈反比关系。此外,羊乳α-CN和β-CN经过不同条件的热处理后再经胃肠消化,抗原性再度明显降低。但仅依据单体蛋白研究还不能十分准确的探究其具体规律性改变,在实际加工过程中,蛋白的分子间也存在交联与聚合现象,这还需进一步探究其中的潜在规律。
猜你喜欢
农业工程学报(2022年8期)2022-08-08 01:47:32
中国饲料(2022年5期)2022-04-26 13:42:32
数学物理学报(2021年6期)2021-12-21 06:24:54
江苏农业科学(2020年24期)2020-02-22 07:42:25
中国酿造(2016年12期)2016-03-01 03:08:23
动物医学进展(2015年10期)2015-12-07 05:46:20
食品工业科技(2014年3期)2014-03-22 13:02:16
食品工业科技(2014年15期)2014-03-11 18:17:43
食品科学(2013年17期)2013-03-11 18:26:49
食品科学(2013年15期)2013-03-11 18:25:41
