
谷氨酰胺对冬毛期水貂生长性能、血清生化指标及抗氧化性能的影响
2021-03-01 11:34李柯汝纪君波孙建华徐燕红司志文李文立
中国畜牧杂志 2021年2期
李柯汝,纪君波,孙建华,徐燕红,,司志文,李文立*
(1.青岛农业大学动物科技学院,山东青岛 266109;2.山东省莱阳市农业农村局,山东莱阳 265200;3.山东菁华农牧发展有限公司,山东诸城 262201)
谷氨酰胺(Glutamine,Gln)在动物体内含量丰富,健康状态下Gln 为非必需氨基酸,但在应激或病理状态下,动物内源合成的Gln 不能满足其自身需要,必须由外源补充,又被称为条件性必需氨基酸[1]。许多研究表明,Gln 是肠黏膜上皮细胞和淋巴细胞的主要燃料,在促进受损伤肠道的修复以及维持正常的局部免疫功能中发挥着不可替代的作用[2-4]。Gln 还参与体内谷胱甘肽(GSH)的合成,具有清除体内各种氧自由基和过氧化物对细胞的损害、维持机体内环境稳定的作用[5]。
在人和动物机体中,Gln 是一种活性小肽,具有抗氧化、免疫以及参与某些物质吸收等多种功能[6]。Krebs[7]于 1935 年第一次发现哺乳动物肾脏具备合成和分解Gln 的能力,并强调指出虽然多数氨基酸都具备多种功能,但Gln 的功能最显著。肖英平等[8]研究表明,Gln 能够提高断奶仔猪的肠道消化吸收能力,促进营养物质的消化率。李文立等[9]研究表明,Gln 能够提高肉鸡小肠的吸收能力。此外,Gln 也是细胞内氮的清除剂及生物合成许多重要物质(如核酸、核苷酸、氨基糖和蛋白质)的重要前体[10]。本试验旨在研究Gln 对冬毛期水貂生长性能、血清生化指标及抗氧化性能的影响,为在冬毛期水貂基础日粮中Gln 的应用提供依据。
1 材料与方法
1.1 试验设计 试验选取140 日龄同性别体重相近的冬毛期短毛黑水貂(美国短毛黑与标准貂杂交后裔)120只,共分为5 组,每组24 个重复,每个重复1 只。其中对照组饲喂基础日粮,试验组在基础日粮中分别添加0.2%、0.4%、0.6%和0.8% Gln(由金维氨生物科技有限公司提供,有效含量为99%),基础日粮组成及营养成分见表1。试验在山东省青岛市即墨区的某水貂养殖场进行,试验时间为2018 年9 月24 日—2018 年12 月9 日,预试验1 周,试验期10 周。试验前对棚舍、笼舍、地面等进行全面消毒,全期自由饮水和采食。
1.2 采血及血清制备 试验结束后,各组随机抽取接近于平均体重的公、母貂各6 只。采用剪趾尖法采集血样3 mL,3 000 r/min 低温离心10 min,离心后取上层血清,分装于1.5 mL 离心管中,-80℃冷冻待测。
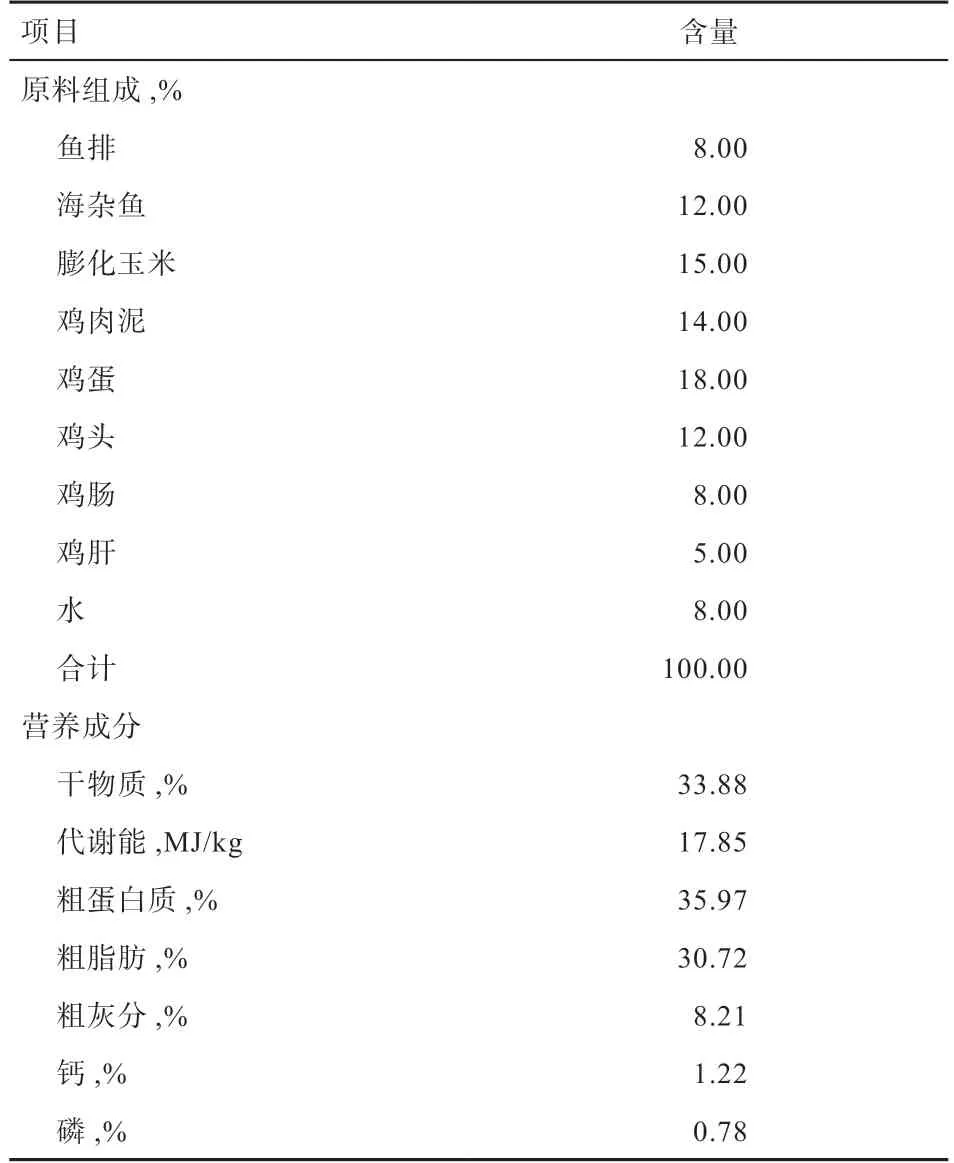
表1 基础日粮组成及营养成分(干物质基础)
1.3 测试指标及方法
1.3.1 生长性能指标 试验期间每天记录给料量和剩料量,分别统计公、母貂的平均日采食量;每周清晨进行空腹称重1 次,分别统计公、母貂的平均日增重;并计算耗料增重比,统计死亡率。试验结束后,各组随机抽取接近于平均体重的公、母貂各6 只处死后,测得鼻尖到尾根的长度为体长,取皮后测得鼻尖到尾根的长度为皮长。
1.3.2 血清生化指标 葡萄糖(GLU)含量采用葡萄糖氧化酶法测定;肌酸激酶(CK)活性采用比色法测定;总蛋白(TP)含量采用考马斯亮兰法测定;尿素氮(UN)含量采用二乙酰一肟法。测定试剂盒购于南京建成生物工程研究所,下同。
1.3.3 血清抗氧化指标 抑制羟自由基(·OH)能力采用比色法测定;总抗氧化能力(T-AOC)采用比色法测定;抗超氧阴离子自由基(O2-)能力采用化学比色法测定;超氧化物歧化酶(SOD)活性采用羟胺法测定;谷胱甘肽过氧化物酶(GSH-Px)活性采用比色法测定;丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定;过氧化氢酶(CAT)活性采用钼酸铵法测定。
1.4 统计分析 采用SPSS 17.0 软件中单因素方差分析(One-Way ANOVA),LSD 法进行多重比较。试验数据以平均值±标准差表示。P<0.05 和P<0.01 分别为差异显著和极显著水平。
2 结果
2.1 Gln 对水貂生长性能的影响 由表2 可知,公貂的平均日采食量、平均日增重、皮长和体长各组间差异均不显著,但4 个试验组的耗料增重比低于对照组(P<0.05)。母貂的平均日采食量、平均日增重、耗料增重比、皮长和体长在各组间差异均不显著。
2.2 Gln 对水貂血清生化指标的影响 由表3 可知,公貂血清中GLU、TP 含量以及CK 活性各组间差异均不显著;但对照组UN 含量高于各试验组(P<0.05),其余各组间差异不显著。母貂血清中GLU、TP 和UN 含量以及CK 活性各组间差异均不显著。因此,水貂基础日粮中添加不同水平的Gln 并不影响母貂血清生化指标。
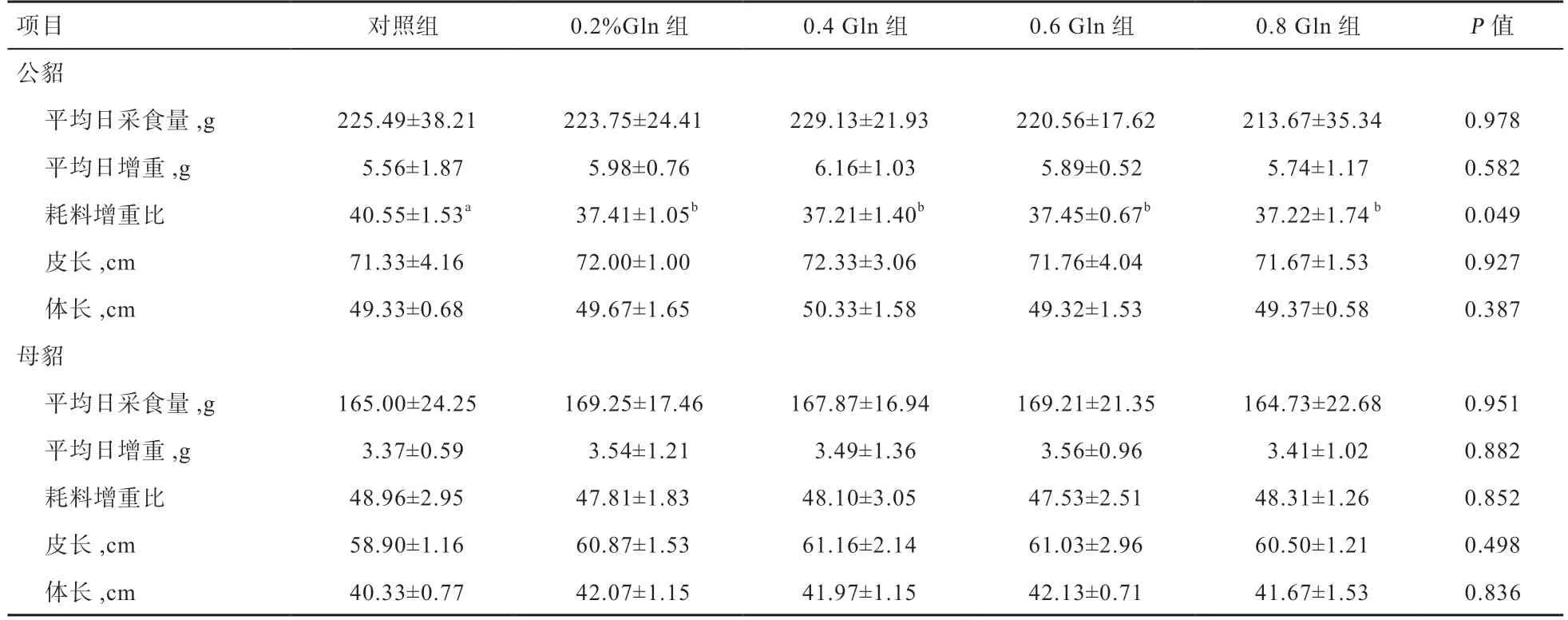
表2 Gln 对公、母貂生长性能的影响
2.3 Gln 对水貂血清抗氧化指标的影响 由表4 可知,公貂血清抑制·OH 能力以及MDA 含量各组间差异不显著。试验组公貂血清抗O2-能力、CAT 活性以及T-AOC 能力各均高于对照组(P<0.05),其余各组间无显著差异。0.4%Gln 组和0.6%Gln 组公貂血清SOD 活性均高于其余各组(P<0.05),0.2%Gln 和0.8%Gln 组高于对照组(P<0.05)。0.4%、0.6%和0.8%Gln 组公貂血清GSHPx 活性高于0.2%Gln 组和对照组(P<0.05),0.2%Gln组高于对照组(P<0.05),其余各组间无显著差异。
由5 表可知,0.6%Gln 组母貂血清抑制·OH 能力高于0.2%Gln 组和对照组(P<0.01),0.6%Gln 组高于0.8%Gln 组(P<0.05),其余各组间无显著差异。母貂血清抗O2-能力以及GSH-Px 活性各组间差异不显著。对照组母貂血清中MDA 含量高于各试验组(P<0.05),各试验组间无显著差异。各试验组母貂血清T-AOC 能力均高于对照组(P<0.01),其中0.6%Gln 组又高于0.2%Gln 组和0.8%Gln 组(P<0.05)。各试验组母貂血清SOD 活性高于对照组(P<0.05),各试验组间差异不显著。各试验组母貂血清CAT 活性均高于对照组(P<0.01),0.2%Gln 组高于0.8%Gln 组(P<0.01)、0.6%Gln 组(P<0.05),其余各组间无显著差异。

表3 Gln 对公、母貂血清生化指标的影响
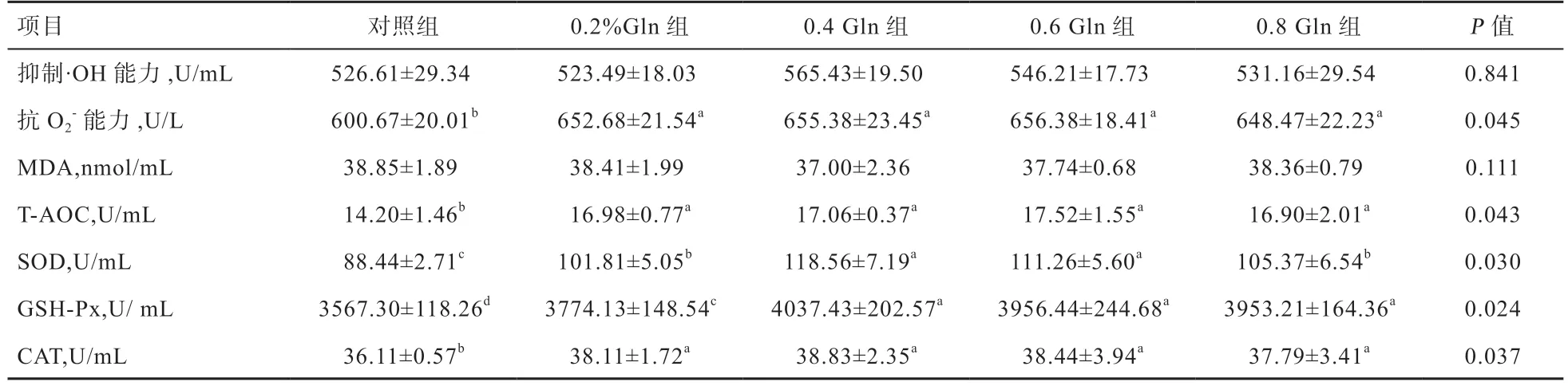
表4 Gln 对公貂血清抗氧化指标的影响
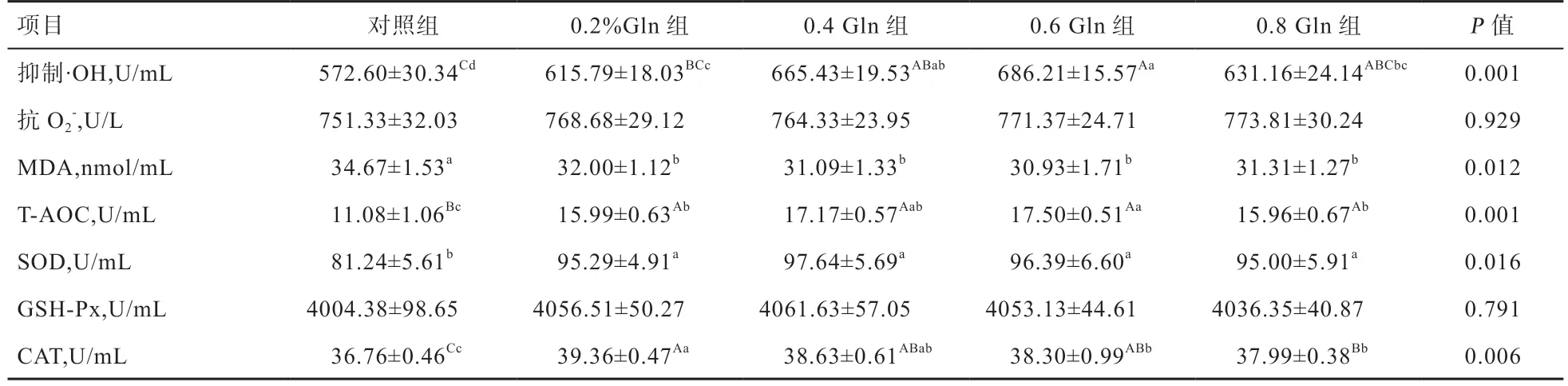
表5 Gln 对母貂血清抗氧化指标的影响
3 讨 论
3.1 Gln 对冬毛期水貂生长性能的影响 Gln 添加水平在动物日粮中的研究有很多。纪君波等[11]研究发现,在育成期水貂基础日粮中添加0.4% Gln 有利于提高水貂的平均日采食量、平均日增重,并显著降低耗料增重比;王录军等[12]研究表明,日粮中添加Gln 对肉兔采食量、日增重和饲料报酬均无显著影响。本试验结果也表明,在水貂基础日粮中添加不同水平Gln 对水貂平均日采食量、平均日增重、皮长和体长均无显著影响,但能显著降低公貂耗料增重比。与育成期相比,冬毛期水貂骨骼生长期已过,由以骨骼发育、器官发育和体增长为主转变为以毛皮生长发育为主,主要是毛皮的生长和脂肪的沉积,致使体增重缓慢[13-14]。
3.2 Gln 对冬毛期水貂血清生化指标的影响 本试验结果表明,不同水平Gln 组水貂血清中的血糖含量无明显变化,所以在冬毛期水貂基础日粮中添加不同水平Gln不能对血清中的血糖含量产生明显影响。TP 在血清固体成分中占比最大,且在水貂体内具有多种功能,一方面作为载体运输物质,维持血管中的正常酸碱度和胶体渗透压;另一方面参与机体免疫防护[1,15-16]。UN 可以反映出机体代谢蛋白质系统的健康程度[2,17-18]。很多研究表明日粮中添加Gln 能够明显提高机体血液中TP 含量,降低UN 含量,但对GLU 含量并没有明显影响[3]。本试验结果表明,虽然在水貂基础日粮中添加不同水平Gln 对公、母貂血清中GLU 含量和CK 活性没有明显影响,但有助于提高TP 含量,并明显降低公貂UN 含量,表明Gln 不仅能够维持机体内代谢环境相对稳定,还有助于提高蛋白质利用率。
3.3 Gln 对冬毛期水貂血清抗氧化性能的影响 T-AOC是衡量机体抗氧化能力的综合性指标,反映机体自由基代谢状态以及对外来刺激的代偿能力[19]。MDA 又是机体氧化应激程度的主要标志物[15]。而机体内抗氧化物酶主要由GSH-Px、CAT 和SOD 组成。研究表明,Gln能够影响机体内GSH-Px、CAT 和SOD 活性,促进GSH 合成,从而提高和维持机体内还原型GSH 水平[18]。而机体内全部自由基都是通过增加抗氧化酶活性来减少氧自由基产生的危害。黄冠庆等[17]研究表明,日粮中添加0.5%和0.8% Gln 能够明显提高30 日龄左右的黄羽肉鸡血液中GSH-Px 活性,并降低MDA 含量,但其对SOD 活性没有明显影响。路静等[20]研究表明,在日粮中添加Gln 能够明显提高肉鸡血清GSH-Px、CAT 和SOD 活性,并且当日粮中Gln 添加水平为1.6%时,其活性最高。本试验结果表明,在水貂基础日粮中添加适量Gln 能够显著提高公貂血清SOD、CAT 及GSH-Px活性、T-AOC 能力、抗O2-能力;显著提高母貂血清SOD、CAT 活性,并显著降低MDA 含量,极显著提高母貂抑制·OH 和T-AOC 能力,且在0.4%添加水平效果最佳。
4 结 论
本研究结果表明,冬毛期水貂基础日粮中添加Gln能降低耗料增重比、MDA 含量以及提高机体的抗氧化性能,且添加水平为0.4%效果最佳。
猜你喜欢
中老年保健(2021年7期)2021-08-22
时代英语·高二(2021年4期)2021-07-29
湖南饲料(2021年3期)2021-07-28
环球时报(2020-11-20)2020-11-20
环球时报(2020-11-12)2020-11-12
环球时报(2020-06-08)2020-06-08
海峡影艺(2019年1期)2019-06-24
农民致富之友(2017年4期)2017-04-10
时尚北京(2016年12期)2017-01-16
食品工业科技(2014年13期)2014-03-11
