
单倍体茄子雄配子发育过程观察及育性分析
2021-02-26 05:15朱海佳崔群香成红林钱丽君柏文军
南方农业 2021年4期
朱海佳 ,崔群香*,成红林,赵 妍,何 芸,邵 英,钱丽君,柏文军
(1.金陵科技学院,江苏南京 211169;2.江宁区农业农村局,江苏南京 211169)
茄子(Solanum melongenaL.)是世界范围内广泛种植的重要蔬菜作物,在亚洲蔬菜生产中具有十分重要的地位[1]。中国茄子栽培历史久远,是茄子次生起源地,茄子类型和品种众多[2]。茄子果实营养丰富,其中富含的维生素E、维生素P 和龙葵碱等成分可以起到抗氧化等保健作用[3]。
自Guha 和Maheshwari[4]在1964 年通过花药培养获得首例单倍体植株以来,运用单倍体技术逐渐成为育种上的热点[5]。在茄子上,Raina 和Iyer[6]首次通过花药培养途径获得单倍体植株。研究表明单倍体植物常常高度不育,如细胞学鉴定表明甜瓜单倍体植株的雄花和雌花均败育[7],西葫芦单倍体植株也出现花粉败育的现象[8]。单倍体具有较高的研究价值,但有性繁殖保存困难,需要加倍成二倍体后才能有效保存和研究利用。茄子单倍体植株加倍困难,尽管能够利用扦插等方式进行繁殖,但繁殖系数很低,且对保存环境要求高,因此极大地限制了茄子单倍体的研究和利用。
研究发现,茄子单倍体植株间性状差异明显,只有部分单倍体植株能够结实,且仅极少量植株的果实中有成熟度较高的不育种子。为了探讨茄子单倍体植株难以座果和种子败育的原因,从而有针对性地采取措施,提高花粉和卵细胞活力、种子活力,实现自交繁殖,有必要对单倍体植株的小孢子产生和雄配子体发育过程进行详细观察分析。
目前,关于茄子单倍体植株减数分裂、雄配子体发育过程及花粉活力的研究未见文献报道。本研究以花药培养再生的茄子二倍体植株84-1自交后产生的单倍体植株D5 为试材,采用冷冻切片法和压片法等对其花粉母细胞减数分裂和雄配子体发育过程进行观察,以揭示其花粉和种子活力降低的原因,试图为单倍体植株的有效利用提供思路。
1 材料与方法
1.1 实验材料
供试材料为本实验室培育保存的单倍体植株D5,该植株春夏季开花但不结实,而可在秋季低温环境下座果。为探索总结从D5 植株获得的减数分裂结束前后的花蕾特征,选择由花药培养直接再生出的单倍体植株D4、D8 和D23 的花蕾,用压片法和染色法进行观察。
1.2 花蕾形态观察
为了探究花蕾形态与细胞发育阶段间的联系,采集D5、D8 和D23 植株上不同大小的花蕾,并将其编号。用游标卡尺测定D5 花蕾总长、萼筒长、花萼基部至抱合的花瓣前端长度、花萼最粗部位的直径、合生花瓣直径等,并通过冷冻切片法观察其细胞发育时期,根据实验结果总结D5 植株的花蕾减数分裂前后的形态特征,并用D4、D8 和D23 植株上的花蕾进行验证。验证时选取不同形态的花蕾,用压片法(方法见“1.5”)观察花药中与生殖相关的细胞发育时期,然后进行对应分析(方法见“1.5”)。
1.3 细胞发育时期鉴定
对D5 各个发育时期的花药进行冷冻切片法观测。冷冻切片法采用新鲜的花药,使用包埋剂将花药覆盖并完全冷冻后,置于切片机上切片,将切片转移到载玻片上,用卡宝品红染色,随后用Nikon 80i光学显微镜观察鉴定。
1.4 花粉活力的测定
取D5 植株上盛开花朵的花粉,置于载玻片上,参照申书兴[9]的方法,用TTC 染色法测定花粉的活力。其中被染成紫红色的花粉为有活力的花粉,无法被染色的花粉为无活力的花粉。计数5 个视野里的花粉粒(总数不少于1 000 粒),根据染色结果计算花粉的活力(染色花粉粒数/花粉粒总数×100%)。
1.5 减数分裂鉴定和染色体计数
使用压片法对D4、D8 和D23 各个发育时期的花药进行观测,并对D5 进行减数分裂鉴定和染色体计数,参照申书兴[9]的方法,采用卡诺固定液固定花药,卡宝品红染色液染色观察。
2 结果与分析
2.1 花蕾形态及细胞发育阶段鉴定
D5花蕾测定结果见表1。观察发现,4号和5号花蕾中,花粉母细胞处于减数分裂时期,6 号之后的花蕾,都处于小孢子发育时期。茄子花粉母细胞减数分裂过程与花蕾大小有密切关系,当花萼基部至合抱的花瓣前端长度比其至花萼筒长度短1~2 mm 时,观察减数分裂效果较好,此时花萼裂片前端基本抱合,仅个别裂片与其他裂片分离。当多数花萼裂片开始分离时,则减数分裂结束,进入小孢子发育阶段。
用D4、D8 和D23 验证的结果见图1 和表2。图1中黑色箭头所示的D4 花蕾中的母细胞处于减数分裂时期,而其余花蕾均已经进入小孢子及随后的发育阶段;白色箭头所示的花蕾尽管与D4 上采摘的最小花蕾大小相似,但其已经完成减数分裂,仔细观察发现其花瓣筒已经露出花萼筒,其余所有花瓣筒超出花萼筒的花蕾均已经完成减数分裂。D8 植株上采摘的1号、2号、3号、7号和8号这5个花蕾中花粉母细胞处于减数分裂时期,其余11 个花蕾均完成分裂;D23 植株上的1 号、2 号、4 号和10 号这4 个花蕾处于减数分裂时期,其余6 个花蕾中的花粉母细胞均完成分裂。形态特征与发育时期的相关性,D4、D8、D23 植株与D5植株类似:花萼裂片前端基本抱合,仅个别裂片与其他裂片分离时处于减数分裂时期;当多数花萼裂片开始分离时,则减数分裂结束,进入小孢子发育阶段。该指标是否适用于二倍体茄子的花蕾有待于验证。
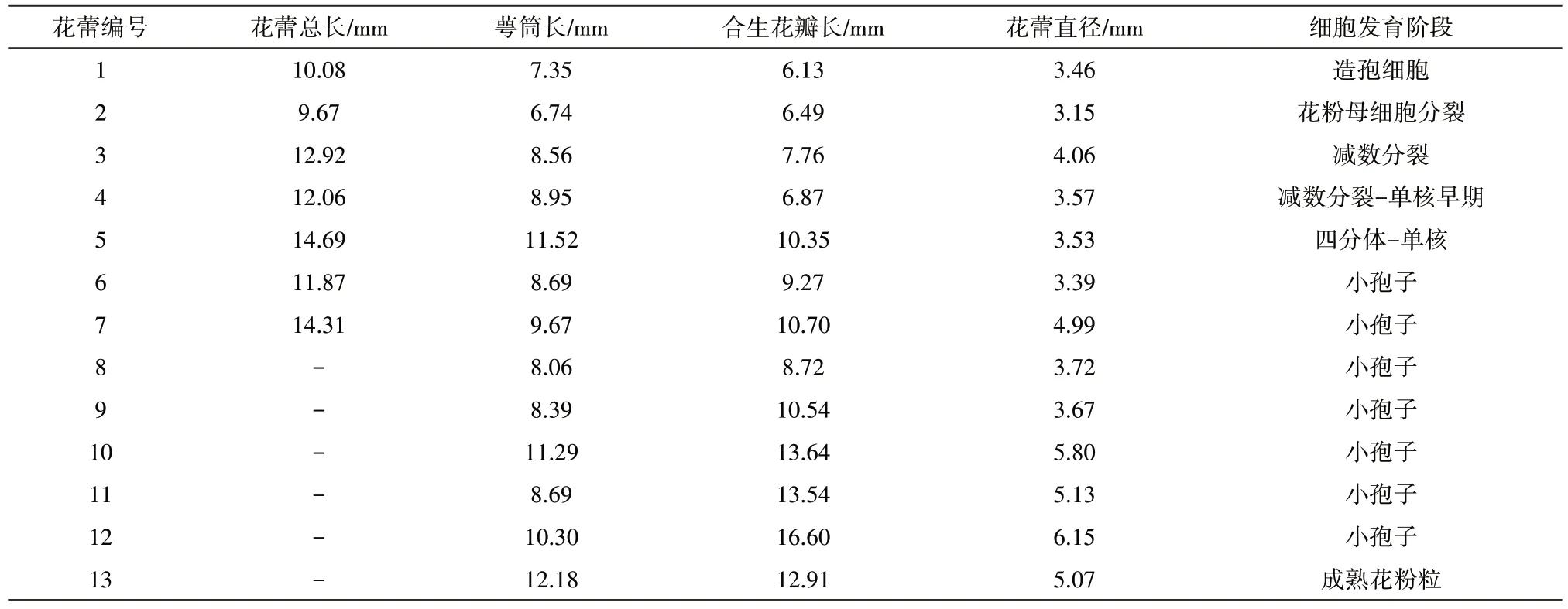
表1 茄子单倍体植株D5不同发育时期的花蕾性状比较

图1 茄子单倍体植株D4、D8和D23的花蕾形态
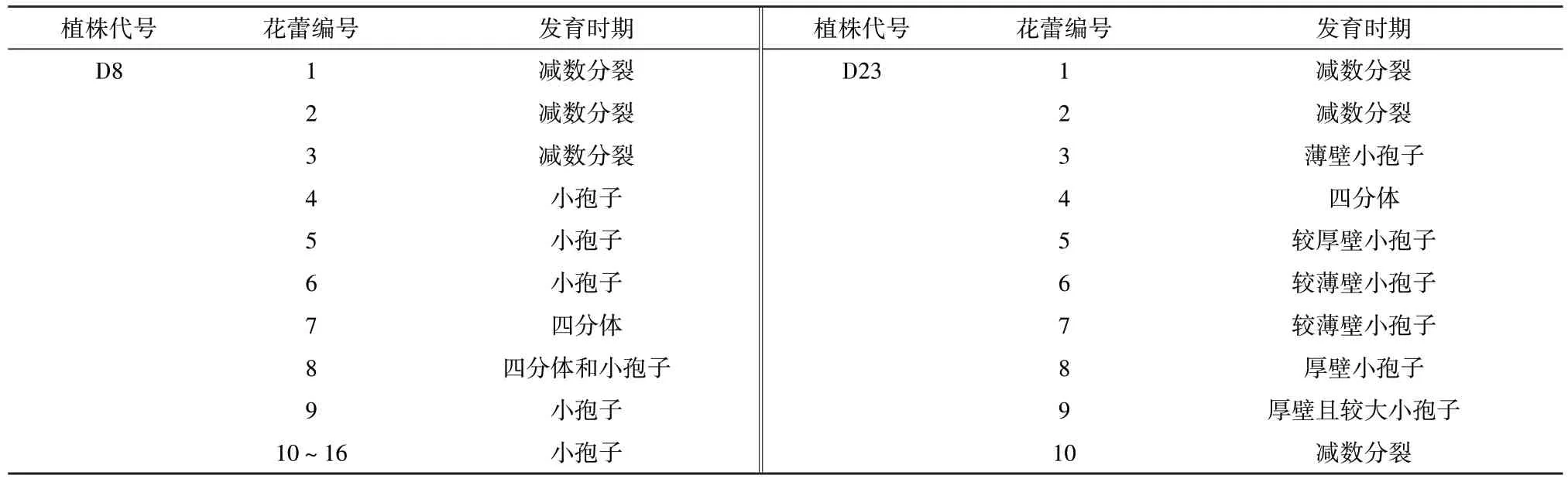
表2 不同单倍体茄子植株花蕾形态与细胞发育时期的关系
2.2 单倍体花药、小孢子和花粉形态鉴定
对花药和花粉进一步观察发现,单倍体的花药结构正常(图2-1),但花粉母细胞由于减数分裂不正常,产生不正常的四分体,甚至“二分体”(图2-2,2-4,2-5)。小孢子多数不能正常发育(图2-3,2-6),仅观察到单核靠边期(图2-7)和少量二核花粉,多数小孢子解体(图2-3),最终只有少量的花粉发育成具有3萌发沟的形态正常的成熟花粉(图2-8)。

图2 单倍体茄子花药、小孢子和花粉粒形态鉴定
2.3 单倍体植株的花粉活力鉴定
花粉经TTC 染色后在光学显微镜下观察发现,花粉粒的大小和形状有明显的不同。其中有活力的花粉粒边缘为红色,中部为黄褐色,而非活性花粉则全部为黄褐色(见图3,箭头示有活力的花粉)。通过计数发现,有活力的花粉占15%,形状和大小差异较大,其结出果实数量少,果实褐变速度快,果实无籽,不能产生后代。
2.4 单倍体植株D5减数分裂过程及染色体数目鉴定
由于冷冻切片采取的花蕾仅切到减数分裂前期、四分体和小孢子时期的花药,因此鉴定染色体数目及对减数分裂整个过程的观察,主要依据压片观察的结果。压片结果显示,单倍体植株D5 染色体复制正常,但联会开始即出现与二倍体不同的现象,中期Ⅰ单价体在细胞中的排列位置分散(见图4-4),但能够清楚地区分出12 个单价体(见图5-1,5-2,5-3),因此D5 植株的体细胞染色体数目为12,证明该植株确实为单倍体。
由图5 可知,单倍体植株D5 减数分裂过程不正常,并不一定产生具有4 个细胞的四分体(图5-14,示5 核花粉母细胞),可能产生各种类型的“四分体”(图5-18)。
2.5 D5植株花粉败育的原因

图4 单倍体植株D5花粉母细胞减数分裂过程

图5 D5植株单倍体减数分裂过程发生败育
在观察单倍体植株D5 的减数分裂过程中发现:中期Ⅰ细胞中有12个单价体,证实该植株确实为单倍体植株;但中期Ⅰ的12个单价体并未排列在赤道面附近,且排列紊乱,染色体聚成2 团或3 团,或各自分散(见图5-1 至5-7);根据中期Ⅱ染色体分布情况(见图5-14,5-15)推测,后期I 以单价体分离为主(见图5-7 至5-12),存在个别单价体染色单体分离现象(见图5-13);单价体分离形式各异,以5/7分离多见,图5-15表示5/7分离的中期Ⅱ,图5-16和图5-17表示后期Ⅱ,显示后期Ⅱ存在非染色单体分离现象,导致四分体中染色体数目严重失衡,产生形态不正常的四分体(见图5-18),甚至是二分体。
从减数分裂中期Ⅰ开始,D5植株的减数分裂就出现不正常,未发现正常的后期Ⅰ细胞,说明植株败育时期在四分体形成以前,因此败育比较彻底。推测有活力的花粉主要来源于后期I 按照12/0 或11/1 分离的花粉母细胞。这两类分离方式占花粉母细胞的比例为2/7,约产生1/7 有活力的花粉,与观察统计出的花粉活力数值15%较为接近。
3 讨论
单倍体是进行体细胞遗传研究和突变育种的理想材料,花药和小孢子培养技术则是获得单倍体的重要途径[10-11]。油菜小孢子培养获得的单倍体可应用于双单倍体育种、诱变育种和转基因育种等研究[12]。相对于正常植株,单倍体植株仅含有半数的染色体,因此单倍体植株比正常植株在形态上较为矮小,且生育率低;同时在后代遗传及生理生化上也表现出一定的差异[13-14]。
张新宇等研究表明山羊豆属植物减数分裂过程存在一定规律,与花蕾大小和颜色密切相关[15]。草本植物关苍术的减数分裂与花蕾大小有密切关系,在花蕾长4~12 mm 时,花粉母细胞进入减数分裂时期[16]。本研究发现茄子花粉母细胞减数分裂过程也与花蕾大小有密切关系,当花柄基部至合抱的花瓣前端长度比花柄基部至花萼筒长度短1~2 mm 时,观察减数分裂效果较好,此期花蕾的形态特征是花萼裂片前端基本抱合,仅个别裂片与其他裂片分离,可以以此作为减数分裂观测取材的依据,也是进行秋水仙素处理花蕾诱导染色体数目加倍的最佳时期;而当多数花萼裂片开始分离时,则减数分裂结束,进入小孢子发育阶段,对于二倍体或多倍体植株,这是采摘花蕾进行花药或小孢子培养的最小花蕾指标,但该指标是否适用于所有不同倍性的茄子植株,还需要进一步验证。研究发现单倍体茄子植株由于减数分裂不正常,不能形成正常的小孢子,因此不适宜进行花药培养或小孢子培养。
本研究所用的D5 单倍体植株春季不结实,至秋季11 月开始坐果,其花粉活力极低(15%)。切片观察显示单倍体茄子的胞质分裂方式属于同时型,后期II 以单价体分离为主,也存在染色单体分离现象,这可能是造成该单倍体植株不育的主要原因。分裂末期常形成大小不一的小孢子,且多数小孢子处于单核靠边期,其后发育即表现不正常,逐渐解体败育,证实单倍体高度不育的特点。张彩霞等研究表明减数分裂过程中多分孢子、落后染色体等异常现象会不同程度地促使小孢子中遗传物质不均衡,从而形成没有育性的孢子,这是引起花粉育性低的重要原因[17]。本研究所用的D5 植株春季不能结实,是否与花粉活力更低有关,需要测定全生育期花粉活力才能验证;另外探讨环境等对花粉活力的影响,并采取措施提高花粉活力,使单倍体植株能够通过有性繁殖加倍为二倍体,是提高茄子单倍体植株保存和利用效率的新思路和新途径,值得进一步研究。
猜你喜欢
现代农业科技(2022年12期)2022-12-13
麦类作物学报(2022年8期)2022-08-08
国际种业前沿动态(2021年1期)2021-03-22
山西农业科学(2020年8期)2020-08-13
江苏农业科学(2020年6期)2020-05-21
三农资讯半月报(2020年5期)2020-03-23
传奇·传记文学选刊(2018年11期)2018-11-26
金山(2018年4期)2018-04-26
天津农业科学(2017年12期)2018-01-10
江苏农业科学(2016年8期)2017-02-15
