
毕节吞天井边缘地带不同郁闭度环境对泥炭藓叶绿素荧光特性及蓄水量的影响
2021-02-26 08:40:32石匡正张朝晖何春梅王智慧
植物研究 2021年2期
石匡正 张朝晖* 何春梅 王智慧
(1. 贵州师范大学贵州省山地环境信息系统与生态环境保护重点实验室,贵阳 550001;2. 贵州师范大学生命科学学院,贵阳 550001)
喀斯特天坑是中国西南喀斯特地形中发现的一种独特且壮观的负地貌,具有相当高的科学和旅游价值,引起了国内外相关研究人员的广泛关注。中国是全球喀斯特天坑家族中发育最完善,种类最齐全,人为干扰程度最深,总量最庞大的国家,是国内外学者前赴后继争相研究的喀斯特王国。研究喀斯特天坑相关植物群落生态、地形地貌、历史演变及其发展现状并对其产生的问题进行探讨,对世界喀斯特环境研究有着极其深远的意义[1~2]。
苔藓植物是自然界中的一大植物门类,是最原始的高等植物,有4.7 亿年的生活史,其生长环境极为丰富多样,几乎遍布在地球上除海洋外的各个角落,具有极其顽强的生命力[3~6]。苔藓植物对整个生态系统具有重要的意义,不仅能丰富物种的多样性,而且对环境污染、矿山找寻等都具有很强的指示作用[7~9]。由于天坑的避难所效应,使得天坑内部的苔藓植物多样性更为丰富,国内外工作目前大都是对天坑内部苔藓植物多样性和环境因子对其影响的研究[10~14],而对坑口苔藓研究甚少。笔者发现,毕节吞天井具有典型的冲蚀型天坑[15]地貌特征,即有地表水流,坑口边缘有大片的泥炭沼泽,属酸性环境,经由长年累月的水流冲蚀作用,且无地下河。而泥炭藓极强的持水特性对自然界的水循环有着相当重要的意义,或许会对天坑形成有影响,因此本研究便以吞天井坑口上游水源天然泥炭沼泽地作为研究背景,以其中生长的泥炭藓为材料展开研究。
泥炭藓科(Sphagnaceae)是苔藓植物门中分布极广的一科,多生长于泥炭沼泽,湿润森林,及高山林下的岩面薄土之上。其生长的土壤及水质呈较强酸性,具有极强的固碳能力,据研究显示,北半球大部分泥潭沼泽地近一半的碳累积都出自泥炭藓的固碳作用[16]。并且泥炭藓作为一种具有超强吸水性的苔藓植物,在医疗卫生、包装运输、环境保护、能源开发及农学和生物学等领域都具有很高的价值[17]。现今泥炭藓又作为一种经济作物的身份出现在耕地和农田中。目前,国外泥炭藓研究众多,如Jonathan Shaw 团队,主要研究方向为苔藓植物的进化和多样性[18~19]。此外,Urban Gunnarsson 曾研究过温度、种间竞争对泥炭藓生长的影响[20];Anna M.Laine曾对泥炭沼泽演替过程中的泥炭藓生长和生态生理学做过研究[21];Sebastian Sundberg 曾研究过以孢子的角度建立泥炭藓的栖息地要求[22];Mulligan R C 曾有研究表明水位是决定泥炭地苔藓植物生长及分布的最重要的环境因素[23]。国内主要是从环境因子和各种胁迫作用对泥炭藓的生长影响的角度展开的研究[24~28]。对于冲蚀型天坑来说,坑口旁地面的水源流入的集中性和水的循环过程是极为重要的,而泥炭藓作为持水性能极强的苔藓植物,对坑口地面外水分循环意义重大[2,29]。不同环境下泥炭藓持水能力的强弱,对生态系统水的分配规律有着重要的影响,本文旨在对喀斯特天坑边缘地带不同郁闭度环境下对泥炭藓叶绿素荧光和蓄水量的影响进行研究探讨,以期对泥炭藓的生长环境做出补充,并为冲蚀型天坑地面外泥炭藓与环境之间的相互作用及其对生态系统水的微循环所带来的重要意义提供参考。
1 材料与方法
1.1 研究样地概况
贵州毕节国家森林公园位于贵州省毕节市境内的乌蒙山区,地处云贵高原东侧梯级斜坡地带,吞天井位于其中的拱拢坪景区内,属北亚热带湿润季风气候区,年平均气温12.8℃左右,冬无严寒、夏无酷暑。吞天井位于景区西北端,为一岩溶奇景——冲蚀型大型天坑。天坑四面环山,坑口南北向约400 m、东西向约150 m,坑深约200 m,地理坐标为27°14′11.01″N,104°59′47.19″E。在吞天井顶部边缘地带,有约6 000 m2的天然野生泥炭沼泽地,其覆盖度由落花溪上游一直延伸到吞天井口。
1.2 研究方法
1.2.1 野外工作
在吞天井顶部边缘地带泥炭沼泽地藓类植物众多,以泥炭藓科(Sphagnaceae)为优势科,其种类分别有粗叶泥炭藓(Sphagnum squarrosum)、秃叶泥炭藓(S.obtusiusculum)、广舌泥炭藓(S.russowii)、泥炭藓(S.palustre)。其中又以泥炭藓为优势种。根据泥炭藓生长环境郁闭度大小,分别以郁闭度为0%,30%,60%,90%(分别为开阔湿地,矮灌木丛边,矮灌木丛下,华山松(Pinus armandii)林下的小水泽)进行取样,并将郁闭度0%环境设为对照组(CK)。每个郁闭度下设5 个1 m×1 m 小样地,使用10 cm×10 cm 的样方框,在每个样地随机取十份样,并分别称取其鲜重(注:取样前当地连续3 d为晴天)。
1.2.2 室内工作
标本采集的同时利用CI-340 便携式叶绿素荧光仪测定了各个郁闭度下泥炭藓的荧光参数,且每个郁闭度下随机测3 次。随后将采回的标本分别装入塑料网袋完全浸泡入水中2 h 至最大饱和度,并悬挂直至不再有水滴下,称重计算饱和含水量。接着将其放入干燥培养箱,70℃恒温进行烘干处理30 h,计算干物质重量和含水量。
1.2.3 数据处理和分析
利用SPSS Statistics 21 对不同郁闭度环境下初始荧光(Fo)、最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)、非光化学淬灭系数(NPQ)、光化学淬灭系数(qP)、光合电子传递速率(ETR)和蓄水量的数据进行分析,使用单因素方差分析(One-way ANOVA),分析不同郁闭度环境对各个指标的显著性影响,最小显著性差异法(LSD)进行两两比较,显著性水平设为0.05;再用Student-Newman-Keuls 法进行所有各组均值间的配对比较,即将各组均值按从大到小的顺序排列,寻找最极端差异。并利用Arc GIS10.2、Origin 9.0和Excel 2010绘制图表。
相关计算公式:
自然吸水率:
饱和吸水率:
自然蓄水量:
饱和蓄水量:

2 结果与分析
2.1 郁闭度对泥炭藓荧光参数的影响
结果显示泥炭藓F0,Fm均随郁闭度的增大而增大(见图2)。对于F0,其中30%郁闭度下比对照增大5.79%,差异显著(P<0.05),60%郁闭度下比对照增大12.61%,差异极显著(P<0.01),90%郁闭度下比对照增大17.51%,差异极显著(P<0.01),且各个郁闭度梯度之间差异都显著(P<0.05)。对于Fm,30%郁闭度下比对照增大2.79%,差异不显著,60%郁闭度下比对照增大3.91%,差异显著(P<0.05),90% 郁闭度下比对照增大9.68%,差异极显著(P<0.01),梯度之间仅60%~90%差异显著(P<0.05),相比增大5.56%。
泥炭藓Fv/Fm的值呈现先降后保持稳定的趋势(见图3),郁闭度为30%时比对照减小1.45%,差异不显著,60%和90%郁闭度下都比对照减小4.35%,差异显著(P<0.05),差异显著且在30%到60%郁闭度变化下,差异发生显著性变化,相比减小2.94%(P<0.05)。
泥炭藓qP 随着郁闭度的增大而逐渐增大,其升幅先陡进而接近平缓,再变陡(见图4)。在郁闭度为30%、60%和90%下变化相比对照皆有极显著性差异(P<0.01),分别增大88.24%、94.12%、170.59%。且在60%~90%郁闭度变化下,差异显著(P<0.05),增大39.39%。泥炭藓NPQ随着郁闭度的增大而逐渐减小,降幅由陡最终趋于平缓(见图4)。在郁闭度为30%、60%和90%下变化相比对照皆有极显著差异(P<0.01),分别减小17.2%、46.5%、49.04%。且在30%~60%郁闭度变化下,有极显著差异(P<0.01),减小35.38%。泥炭藓ETR随着郁闭度的增大而逐渐增大,呈现增幅先陡后缓再变陡的趋势(见图4)。在郁闭度为30%和60%下变化相比对照皆显著(P<0.05),分别增大29.9%和36.71%,在90%下变化相比对照有极显著差异(P<0.01),增大69.23%,且在60%~90%郁闭度变化下,差异显著(P<0.05),增大23.79%。
2.2 郁闭度对泥炭藓吸水率、蓄水量的影响
结果显示,泥炭藓自然吸水率随着环境郁闭度的增大呈现逐步上升的趋势,且在90%郁闭度下增幅最为显著,30% 郁闭度下比对照增大1.67%,60% 和90% 郁闭度下分别比对照增大30.92%和83.64%(见表2),有极显著差异(P<0.01);饱和吸水率随着郁闭度增大也呈升高趋势,同样在90%郁闭度下增幅最为显著,30%郁闭度下比对照增大2.86%,60%和90%郁闭度下比对照分别增大38.66%和24.47%(见表2),有极显著差异(P<0.01);且在90%郁闭度下自然吸水率与饱和吸水率相差很小,其余都有明显不同。
泥炭藓的自然蓄水量和饱和蓄水量都随着郁闭度的增大呈升高趋势,在90%郁闭度下增幅最为显著。且在90%郁闭度下自然蓄水量与饱和蓄水量相差很小,其余都有明显不同。自然蓄水量在30%、60%和90%郁闭度下比对照分别增大65.27%、126.42%和322.56%,有极显著差异(P<0.01);饱和蓄水量在30%、60%和90%郁闭度下比对照分别增大75.65%、89.5%和187.92%(见表2),有极显著差异(P<0.01)。
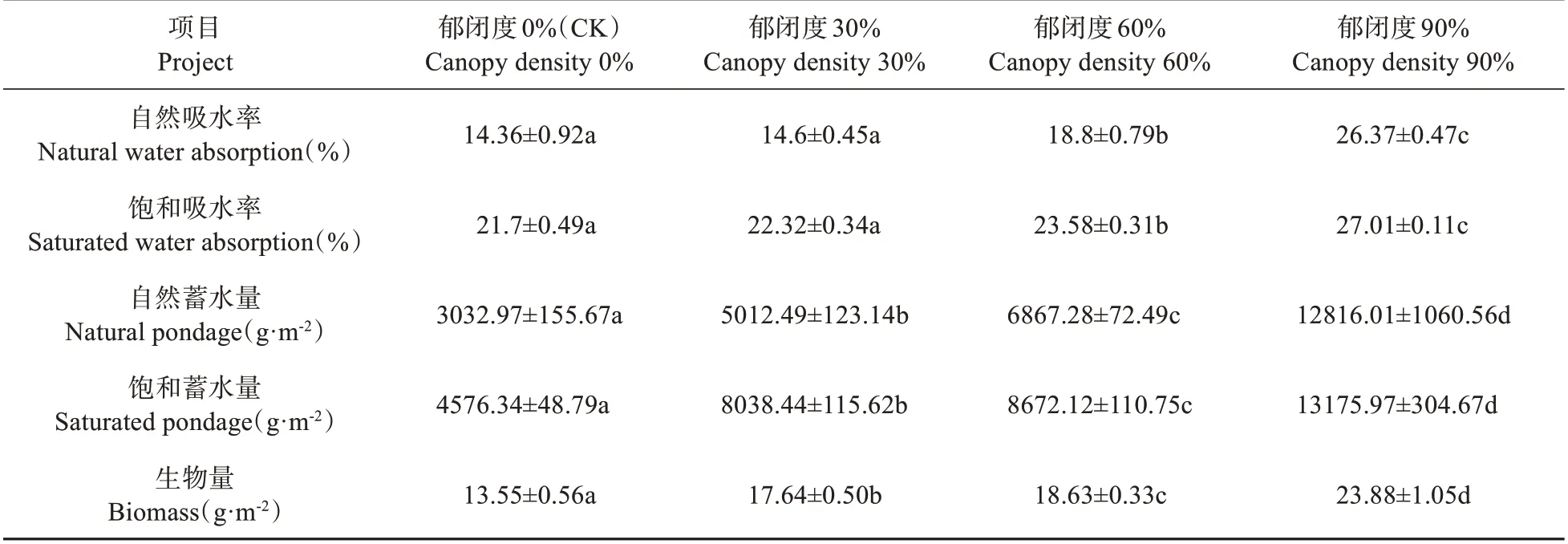
表1 泥炭藓在不同郁闭度环境下的吸水率、蓄水量和生物量的变化(平均值±标准偏差)Table 1 Changes in water absorption,pondage and biomass of S.palustre in different canopy density(mean±SD)
3 讨论
3.1 不同郁闭度环境下生长的泥炭藓荧光特性的不同
植物的光合作用是植物生存和发展的基础,决定了植物生产力的大小,而叶绿素荧光参数能够灵敏、无损地展示出植物光合功能对环境变化的响应,其参与了植物光合作用的整个反应过程,包含了一系列植物生理响应机制,对我们研究植物生长有着极其重要的指示意义[30]。不同的植物对其生活的环境要求不同。郁闭度环境不同,其光照强度,环境温度,空气潮度、土壤湿度乃至可利用土壤肥力都不相同。进而使得植物光合作用强度和叶绿素荧光特性都有变化。如遮荫处理导致甜菜树叶片PSⅡ最大光化学效率(Fv/Fm)和实际光化学效率(ФPSⅡ)下降[31];遮阴后耐弱光基因型马铃薯具有较高的净光合速率、较低的光补偿点、较低的CO2补偿点、较快的光诱导反应速度和较高的非光化学热耗散能力[32]。
本研究中,泥炭藓的初始荧光(Fo)和最大荧光(Fm)都随着郁闭度的增加而显著增大,说明中度遮阴有利于泥炭藓的叶绿素积累,从而促进其生长,而在重度遮阴环境下,由于其生长水位极大而使得其生长依旧最好,这与Mulligan R[23]的研究相吻合。杨冠松等曾对不同遮阴程度下的甜菜树叶绿素荧光进行研究,发现(Fv/Fm)在80%遮阴环境下显著降低,即发生了光抑制[31];范贝贝等曾研究黄绿型和红型两种泥炭藓在践踏过程中(Fv/Fm)的变化,前者无显著变化,后者略有下降,从而推断黄绿型泥炭藓具有较强的践踏耐受性[33]。本研究中泥炭藓最大光化学效率(Fv/Fm)在郁闭度为60%时略有降低,接着又趋于不变,可能是由于泥炭藓具有一定的耐光性,全光或遮阴并不会对其产生胁迫作用,在郁闭度为60%时略有降低的原因可能是灌木林与其的竞争作用,导致土壤水分和营养元素的吸收减少,所以产生了胁迫作用,之后又因为其良好的耐受能力而维持不变。这与资源胁迫假说及马进泽的研究相吻合[24,34~35]。泥炭藓良好的抗胁迫能力,或许正是其在各种环境下能生存的原因,奠定了其能始终维持自身生物特性和对生态系统水的微循环的调节作用。
叶绿素荧光光化学淬灭系数(qP)表示植物将光能转化为化学能的能力和效率,非光化学淬灭系数(NPQ)则表示PSⅡ所吸收的光能不能用于光合电子传递,而是以热能的形式消耗掉。梁文华[36]曾做过遮阴对金叶风箱果叶绿素荧光参数的影响,发现在80%遮阴环境下,电子传递速率明显升高,而热耗散的量子比率由0.36 下降至0.25,唐星林[37]则发现当遮阴程度为全光,50%遮阴,78%遮阴时,闽楠电子传递速率分别为52.46、55.30、71.34,呈正相关,本研究中,随着郁闭度的增大,泥炭藓光化学淬灭系数(qP)由0.17 升至0.46,光合电子传递速率(ETR)由5.72 升至9.68,都显著增大,非光化学淬灭系数(NPQ)由1.57 降至0.8,显著减小,说明遮阴能提高泥炭藓的光能利用效率,并通过降低热耗散比例从而适应弱光生境。
3.2 不同郁闭度环境下生长的泥炭藓吸水率和蓄水量的不同
苔藓植物因具有极强的持水特性,对整个喀斯特森林地带的生态水循环有着重要的生态学意义,而泥炭藓作为吸水能力最强的苔藓植物,研究其区系吸水率和蓄水量有助于我们认识喀斯特高原森林生态系统水的微生态循环过程与分布规律[38]。
随着生长环境郁闭度的增大,泥炭藓的吸水率与蓄水量都发生了显著增大,且在全光无遮蔽物与30%郁闭度环境下其自然吸水率与饱和吸水率相差最大,说明持续的强光照射会降低泥炭藓的自身蓄水量,并在一定程度上破坏其吸水性能。而遮阴的确有效地促进了其生物量的累积,从而增大其蓄水量。在郁闭度为90%下,因其生长环境为华山松林下的小水泽,有丰富的自然水分条件,故拥有极大的自然和饱和蓄水量,且二者相差无几。在毕节吞天井顶部的开阔湿地中,常年杂草丛生,且阳光直射程度高,泥炭藓生长虽具有一定规模,但生物量较小且植株较为矮小细弱,而在井口的泉边华山松林下,泥炭藓的植株生长的更加粗壮高大则为本文的研究结果提供实际的依据。而不同郁闭度下泥炭藓的持水能力对地表径流水的分配规律有着重要意义,从而对吞天井的形成产生影响。
4 结论
泥炭藓具有良好的抗光胁迫能力,可在各种光照条件下正常生存,但是郁闭度在30%~90%的环境下更有利于其叶绿素的积累,促进光合作用的进行,进而产生更大的生物量和蓄水量。且在泥炭藓周围生长的其他种群的植物会与其产生竞争关系,分享土壤水分和营养元素,从而影响泥炭藓的生长,更多的水分有助于它的生长。泥炭藓的生长及其与环境间的相互作用可能对吞天井外部地面的生态系统水循环产生更有益的影响,而泥炭藓极强的吸水性在减缓地表径流上具有重要作用,这或许对减慢吞天井这一冲蚀型天坑的形成有着极为重要的意义。
猜你喜欢
新疆地质(2024年1期)2024-04-17 08:55:27
今日农业(2021年15期)2021-10-14 08:20:06
检察风云(2021年13期)2021-09-14 09:19:30
福建林业(2020年5期)2020-03-18 08:23:02
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
中国三峡(2018年11期)2018-01-30 15:25:17
干旱地区农业研究(2016年2期)2016-04-25 01:15:51
防护林科技(2015年12期)2016-01-13 11:49:24
小主人报(2015年3期)2015-02-28 20:42:09
