
天山林区土壤真菌多样性及其群落结构
2021-02-25 06:56王诗慧常顺利张毓涛
生态学报 2021年1期
王诗慧,常顺利,*,李 鑫,张毓涛
1 新疆大学资源与环境科学学院绿洲生态教育部重点实验室, 乌鲁木齐 830046
2 新疆林科院森林生态研究所, 乌鲁木齐 830063
真菌能够分解有机质、参与物质和能量循环,是土壤微生物多样性的重要组成部分。土壤真菌的种类结构可以作为评价土壤生态状况的重要生物指标,而土壤真菌群落的结构又受植物物种及其根系分泌物的影响[1-3]。在全球尺度上,土壤真菌丰富度与植物多样性存在直接的相关关系[4];青藏高原高寒草原土壤真菌的α多样性和β多样性与地上植物的α多样性和β多样性呈现出强烈耦合关系,地上植物多样性是制约土壤真菌多样性的首要因素[5]。而土壤真菌也是植被参与生态系统循环的重要推动者,通过共生或者病原方式与植物构成密切的联系。因此,了解土壤真菌群落结构对于探索植物繁育、疾病防治、评价生态系统功能、维护生态系统平衡等方面至关重要。
通常,土壤真菌群落基于传统的培养方法进行分析,已经清楚地证明了植物对真菌群落的依赖性[6]。然而,传统培养所能得到的土壤真菌数量非常少,有许多真菌无法直接被分离培养。高通量测序就是从土壤中直接提取总DNA富集后进行测序,具有高效、信息全面等特点,为土壤微生物的研究提供有利手段[7,8]。
天山山系是亚欧大陆最大的东西走向的独立山系之一,天山林区在涵养水源、调节气候、固碳释氧等方面具有非常重要的生态系统功能[9]。天山林区以雪岭云杉(Piceaschrenkiana)纯林为主,在涵养水源、积累营养物质、保护生物多样性等方面具有重要的生态功能。雪岭云杉林缘、林窗及林下的灌木主要有栒子(Cotoneastermelanocarpus)、异果小檗(Berberisheteropoda)、密刺蔷薇(Rosaspinosissima)、金丝桃叶绣线菊(Spiraeahypericifolia)、新疆方枝柏(Sabina pseudosabina)、锦鸡儿(CaraganaFabr)、刚毛忍冬(Lonicerahispida)等,林下土壤类型为典型山地灰褐色森林土[10]。雪岭云杉一直被报道存在老龄化严重,幼苗成活率低等更新不良的问题,目前关于天山植物的研究大多基于种群空间变化或生态化学计量等方面的研究[11-12],对植物与土壤真菌多样性及其群落结构的关系研究较少。基于此,本研究使用Illumina MiSeq 高通量测序技术,分析了天山不同植被类型的根际土与非根际土中真菌群落组成及多样性,探讨了不同植物土壤微生物的群落动态及优势菌属,以期为探寻影响云杉生长的优势菌属、对天山植物的保护及经营管理提供理论基础。
1 材料与方法
1.1 实验材料
本文的土壤样品采集于天山森林生态系统定位研究站(87°07′—87°28′E,43°14′—43°26′N,驻地海拔1908 m),年均气温2—5℃,年降水量400—600 mm,土壤类型为灰褐色森林土。选择6种植被为研究对象取土壤样品,分为别为5种灌木的根际土与非根际土,不同生境下云杉的根际土与非根际土,共12个处理样品,每个处理3次重复取样,共计36个测试土样。天山北坡为阳坡,5种灌木分布在海拔1600—1760 m 之间;采样时选取海拔、林分密度基本相同的灌木纯林,设置10 m×10 m的样方,每种灌木设置三个样方,样方内采用5点法取样,去除表面凋落物后用抖落法取灌木根际土样;云杉林分布于海拔1760—2800 m之间,阴坡的云杉生长状况优于阳坡,用以上相同方法取云杉的根际土与非根际土,但云杉林窗面积较小且没有植物生长,一个林窗为一个样方,3个样方之间间隔20 m以上,去掉表面凋落物后挖到云杉根系时采集根际土,未挖到根系的表层土为非根际土。将同处理土样混合均匀并取出可见的植物根系和动植物残体并保存于灭菌的密封袋冷藏带回,带回实验室后立即过2 mm土壤筛编号(表1)并分为两份,一份用于土壤理化性质的测定,一份用于土壤真菌DNA提取。
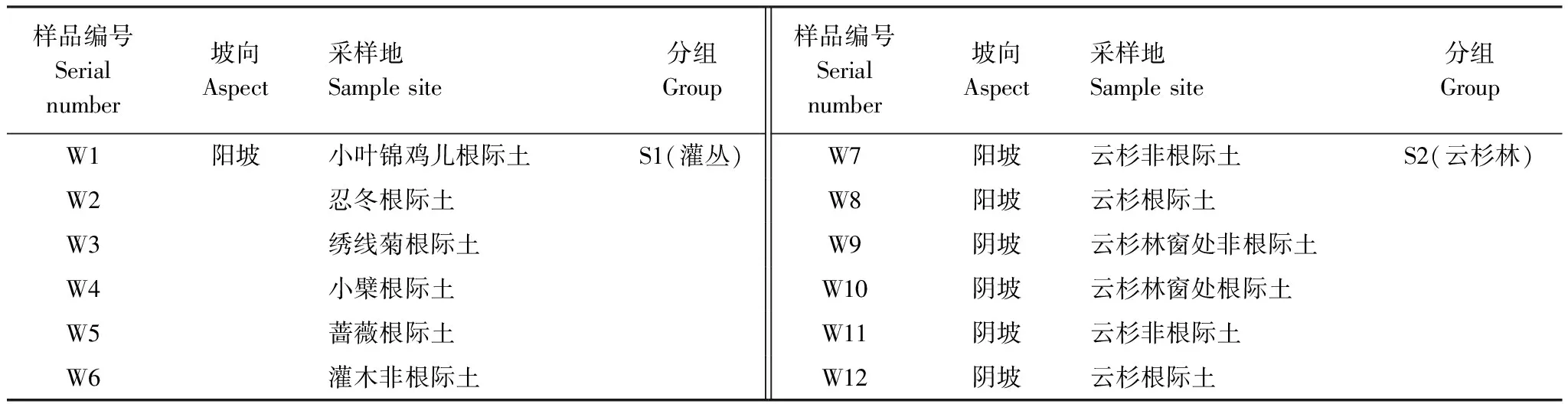
表1 样品编号
1.2 测序分析
采用MoBio试剂盒法提取土壤样品DNA,之后用1%的琼脂糖凝胶电泳检测DNA的纯度和浓度。稀释后的基因组DNA为模板,对真菌ITS1区域进行PCR扩增,引物序列为ITS1:5′-TCCGTAGGTGAACCTGCGG-3′, ITS4:5′-TCCTCCGCTTATTGATATGC-3′;使用New England Biolabs 公司的Phusion® High-Fidelity PCR Master Mix with GC Buffer和高效高保真的酶进行PCR,确保扩增效率和准确性。PCR产物用2%浓度的琼脂糖凝胶电泳进行检测,选择主带大小在400—450 bp之间的序列,割胶回收目标条带。由诺禾致源生物科技有限公司使用Illumina公司TruSeq DNA PCR-Free Library Preparation Kit建库试剂盒进行文库的构建,构建好的文库经过Qubit定量和文库检测,合格后使用HiSeq进行上机测序。
1.3 土壤理化性质分析
土壤样品风干后过0.149 mm筛,土壤总有机碳(Total Organic Carbon,TOC)采用重铬酸钾氧化-外加热法测定;土壤全N(Total Nitrogen,TN)采用扩散法;土壤全P(Total Phosphorus,TP)采用酸溶-钼锑抗比色法;土壤全K(Total Potassium,TK)采用 NaOH 碱熔-火焰光度计测定法。
1.4 数据分析与处理
使用Excel 2017对土壤样品中C、N、P、K 的数据进行分析;对ITS1区进行测序,下机所得原始数据(Raw data)经滤除后得到高质量数据(Clean data),使用FLASH v1.2.7软件、Trimmomatic v0.33软件、UCHIME V4.2软件获得样品的有效数据(Effective data)并在97%的相似度水平下进行聚类获得OTUs,每个OTU认为可代表一个物种。使用Qiime软件计算微生物群落的α多样性指数,群落多样性用Shannon指数和Simpson指数表示,Chao指数表示土壤微生物群落的丰富度,Coverage指数表示测序深度。利用样本间生物学距离进行β多样性分析。选取OTUs的代表性序列用对不同样品内微生物进行物种注释分析,获得分类学信息并分别在界(Kingdom)、门(Phylum)、纲(Class)、目(Order)、科(Family)、属(Genus)、种(Species)分类水平统计个样本的群落组成,利用Qiime生成不同分类水平上的物种丰度表,再使用R语言工具绘制样品各分类学水平下的群落结构图、Venn图、稀释曲线图。使用 Canoco 5中的冗余分析(Redundancy Analysis,RDA),探讨不同环境因子对真菌群落的影响。
2 结果与分析
2.1 不同植被类型土壤理化指标
对两组样品的土壤理化性质进行测定,天山土壤有机碳含量较高,天山云杉林下土壤有机碳、全氮、全磷含量高于灌丛,云杉与灌木土壤理化性质差异并不明显。
2.2 山林区土壤样品的测序结果
滤除低质量和短序列的读数后共得到957120条序列,为研究样品的物种组成多样性,对所得优质序列按≥97%的相似度进行OTU聚类,得到14121个OTUs,数据有效率为95.03%—98.68%,灌丛(S1)和云杉林(S2)得到的有效序列分别为97.19%和97.18%,说明原始数据利用率较高,绝大部分都可以用于后续的数据分析。随着测序数量的增加,所有样品的稀释曲线的斜率逐渐上升,最终都趋向于平缓(图1),且alpha多样性的文库覆盖度都大于0.99(表4),说明测序数据可以反映样品中真菌群落的实际情况。

表2 天山林区不同植被类型土壤理化指标

图1 不同植被类型土壤所得OTU稀释曲线图
2.3 不同植被类型下土壤真菌的多样性
反映土壤真菌的多样性的Shanno指数、Simpson指数和反映物种丰富度的Chao1指数、ACE指数表明,真菌群落结构和丰富度在不同植被类型中差异显著(表3)。总体来说,灌丛的真菌多样性和丰富度高于云杉林,非根际土中真菌的多样性和丰富度高于根际土,其中小叶锦鸡儿(W1)的土壤真菌多样性和丰富度都显著高于其他植被类型,云杉林窗处根际土(W10)的真菌多样性和丰富度最低。云杉林中,阴坡的土壤真菌群落结构和丰富度较高,阳坡次之,林窗最低。

表3 不同植被类型土壤真菌α多样性统计分析
2.4 不同植被类型土壤真菌群落OTU特异性分析
使用韦恩图分析天山林区土壤真菌的物种组成(图2)。灌丛(S1)和云杉林(S2)分别检测到2989和2458个OTU,两组共有OTU数量为1723个,占各组OTU的57.64%和70.09%。S1中,W1—W6共有的OTU数量为201,各自特有的OTU占总OTU的17.91%、11.08%、5.69%、8.61%、4.61%、3.35%,非根际土(W6)中的真菌多样性要低于根际土。S2中,W7—W12共有的OTU数量为139个,各样品特有OTU数目占总OTU的4.97%、7.69%、17.74%、2.41%、13.54%、10.87%,W10特有的OUT数最少,W9最多。灌丛和云杉林的根际土中,共有的OTU数目为148,特有OTU数目W1>W2>W12>W4>W3>W8>W5>W10;由以上可知,云杉林根际土中特有OTU数量少于灌木;灌丛根际土和云杉林窗的根际土中特有OTU数量大于非根际土;阳坡和阴坡的云杉非根际土中特有的OTU数目都大于根际土。
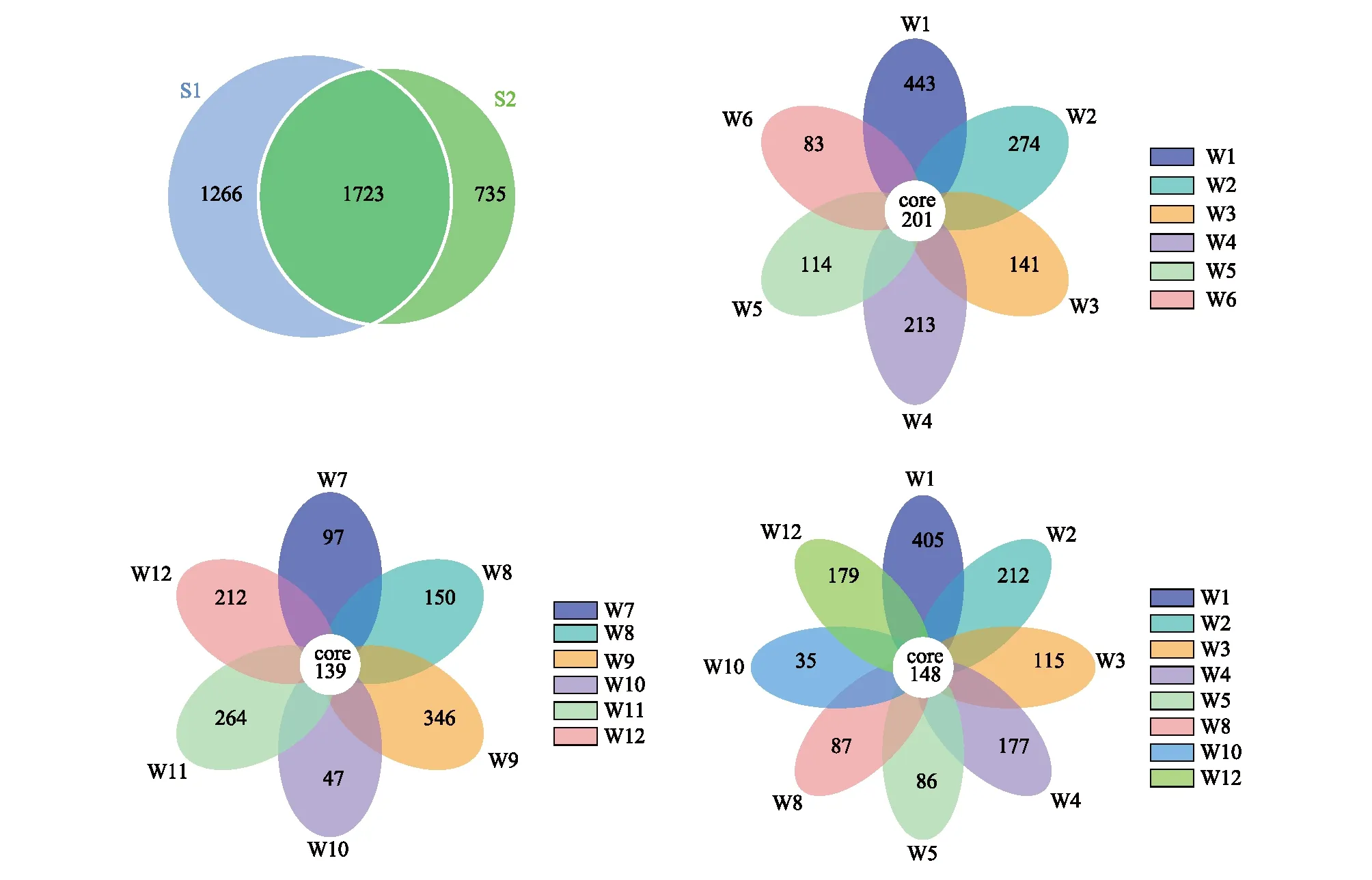
图2 不同植被类型土壤真菌OTU数量韦恩图
2.5 不同植被类型下土壤真菌群落组成
对得到的OTU序列进行注释,分为7个门、33个纲、109个目、255个科、444个属、677个种。通过对样品进行种群组成分析,可以反映其在不同分类学水平上的群落结构,有利于分析样品之间的差异,从而分析其群落结构组成的特点。菌群在门水平上占比最多的是子囊菌门(Ascomycota)57.69%,其次为担子囊菌门(Basidiomycota)30.47%,接合菌门(Zygomycota)9.35%、壶菌门(Chytridiomycota)1.2%和球囊菌门(Glomeromycota)1.1%(图3)。其中子囊菌门(Ascomycota)和担子囊菌门(Basidiomycota)在灌丛和云杉林两个组中分别占比为62.48%、27.94%和52.89%、32.98%,且在11个样品中相对丰度均大于12%,显著高于其他真菌,是优势菌门;云杉林中的担子囊菌门(Basidiomycota)和接合菌门(Zygomycota)占比数量显著高于灌丛;子囊菌门(Ascomycota)在W10中丰度最大、担子囊菌门(Basidiomycota)在W8和W12中丰度较大、接合菌门(Zygomycota)在W9中丰度最大。
图3分别显示样品中丰度排名前10的纲(Class)水平和属(Genus)水平的群落结构分类比较结果:相对丰度较高的前10个纲种类为粪壳菌纲(Sordariomycetes)、伞菌纲(Agaricomycetes)、未定接合菌纲(Incertae_sedis_Zygomycota)、座囊菌纲(Dothideomycetes)、锤舌菌纲(Leotiomycetes)、盘菌纲(Pezizomycetes)、古菌纲(Archaeorhizomycetes)、散囊菌纲(Eurotiomycetes)、银耳纲(Tremellomycetes)、壶菌纲(Chytridiomycetes)(图3),其中粪壳菌纲(Sordariomycetes)和伞菌纲(Agaricomycetes)丰度最高,在灌丛和云杉林中分别达到27.09%、26.04%和26.84%、31.45%。
从属的分类水平分析(图3),相对丰度较高的为:帚枝霉属(Sarocladium)、鬼笔属(Lysurus)、镰刀属(Fusarium)、赤霉属(Gibberella)、丝盖伞属(Inocybe)、Microidium、大环柄菇属(Lepiota)、微皮伞属(Marasmiellus)、盾盘菌属(Humaria)、Rhytidhysteron。灌木和云杉在属水平上的优势菌群有明显差异,帚枝霉属(Sarocladium17.86%)、鬼笔属(Lysurus5.35%)、Microidium(3.18%)是灌丛中的优势菌属;云杉林中相对丰度较高的属有镰刀属(Fusarium4.41%)、赤霉属(Gibberella4.70%)、丝盖伞属(Inocybe4.56%)。TOP35在各组之间分布差异明显,其中,W1中丰度较高的有环柄菇属(Lepiota)、帚枝霉属(Sarocladium)、镰刀属(Fusarium);W2中丰度较高的有帚枝霉属(Sarocladium)、环柄菇属(Lepiota)、Microidium、被孢霉属(Mortierella);W3中丰度较高的有鬼笔属(Lysurus)、帚枝霉属(Sarocladium)、被孢霉属(Mortierella);W4中丰度较高的有帚枝霉属(Sarocladium)、Microidium、Rhytidhysteron;W5中丰度较高的有帚枝霉属(Sarocladium)、湿伞属(Hygrocybe)、被孢霉属(Mortierella);W6中丰度较高的有帚枝霉属(Sarocladium)、Articulospora、枝孢属(Cladosporium);W7中丰度较高的有微皮伞属(Marasmiellus)、赤霉属(Gibberella)、被孢霉属(Mortierella)、欧立菌属(Oliveonia);W8中丰度较高的有丝盖伞属(Inocybe)、拟锁瑚菌属(Clavulinopsis)、帚枝霉属(Sarocladium);W9中丰度较高的有Boeremia、丝盖伞属(Inocybe)、Halokirschsteiniothelia;W10中丰度较高的有镰刀菌属(Fusarium)、赤霉菌属(Gibberella)、根霉属(Rhizopus);W11中丰度较高的有盾盘菌属(Humaria)、被孢霉属(Mortierella)、单端孢属(Trichothecium);W12中丰度较高的有丝盖伞属(Inocybe)、单端孢属(Trichothecium)、被孢霉属(Mortierella)。云杉根际土中共有的所占比例较多的属有丝盖伞属(Inocybe)、被孢霉属(Mortierella)、赤霉属(Gibberella)、镰刀属(Fusarium),这些属可能与云杉的生长有密切的关系。
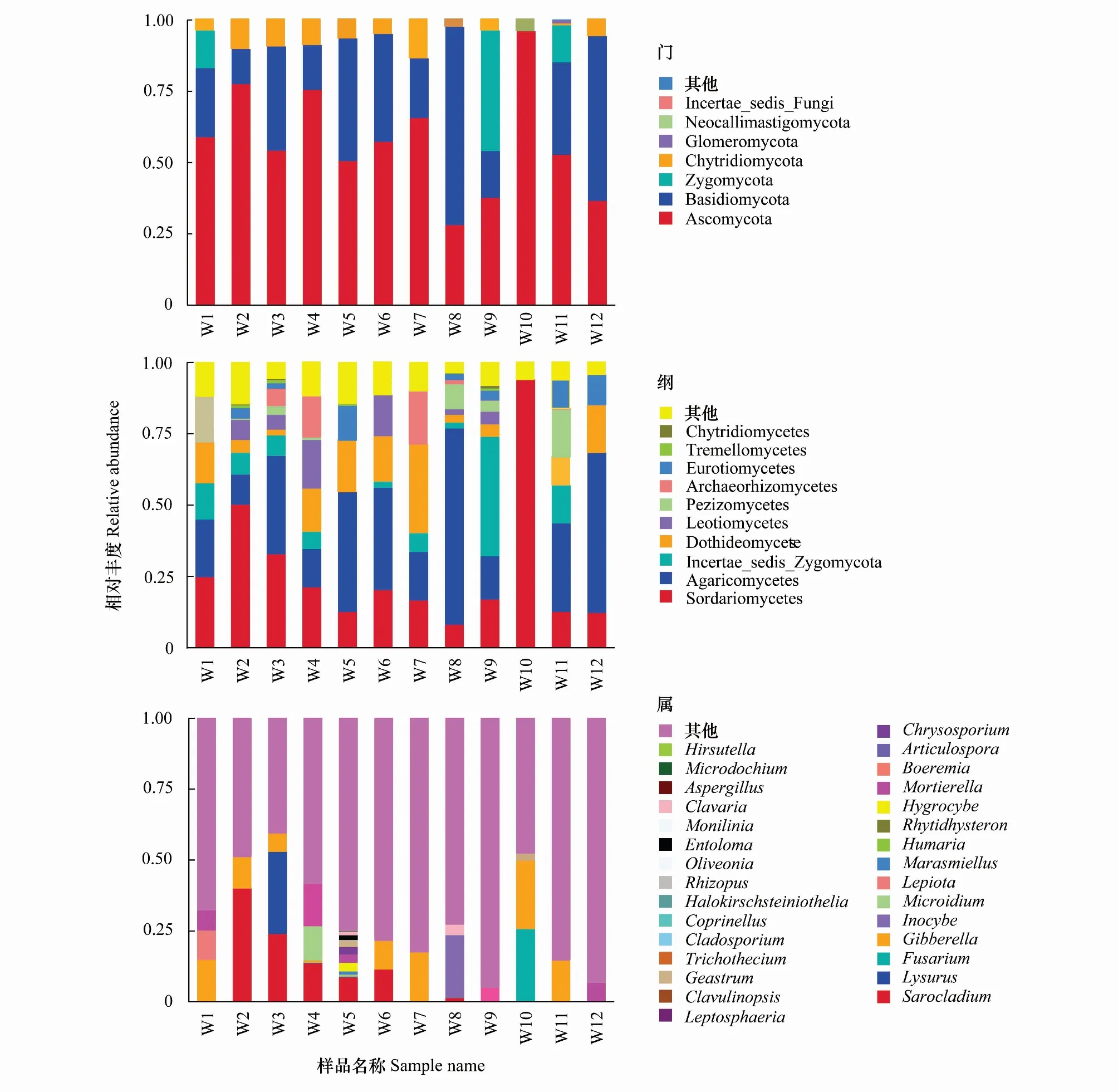
图3 天山林区土壤真菌群落在分类水平的相对丰度
2.6 不同植被类型土壤真菌群落差异分析
通过β多样性的分析结果,可以看出样品间的群落结构差异是否与生物学分组一致。基于OTU水平对天山植物根际土壤样品进行UPGMA(图4)和PCoA(图5),结果表明不同植被类型的真菌群落相似性差异较大。基于加权的Unifrace 距离矩阵进行分析,将灌丛(S1)和云杉林(S2)分为两组,其真菌群落多样性差异显著。根据样品间的相似关系,真菌群落可以分为三组:林窗根际土(W10)的真菌群落组成与其他土样差异较大,被单独列为一组;小叶锦鸡儿(W1)和阳坡非根际土(W7)的菌群结构相似,林窗的非根际土(W9)和林下的非根际土(W11)群落结构相似,它们与阳坡和阴坡的根际土群落结构也较为靠近;忍冬(W2)、小檗(W4)、蔷薇(W5)、绣线菊(W3)、灌木的非根际土(W6)被分为一组。基于加权的PCoA分析,横纵坐标表示OTU水平真菌群落结构的变异分别为35.31%和17.34%,由图7可以看出,PC1维度上可以将灌丛和云杉林两组分开,即PC2是造成云杉和灌木真菌群落产生差异的主成分。在PC2维度上,云杉林的非根际土与几种灌木根际土的距离较近,表明云杉的非根际土与灌木的土壤真菌群落结构较为相似。
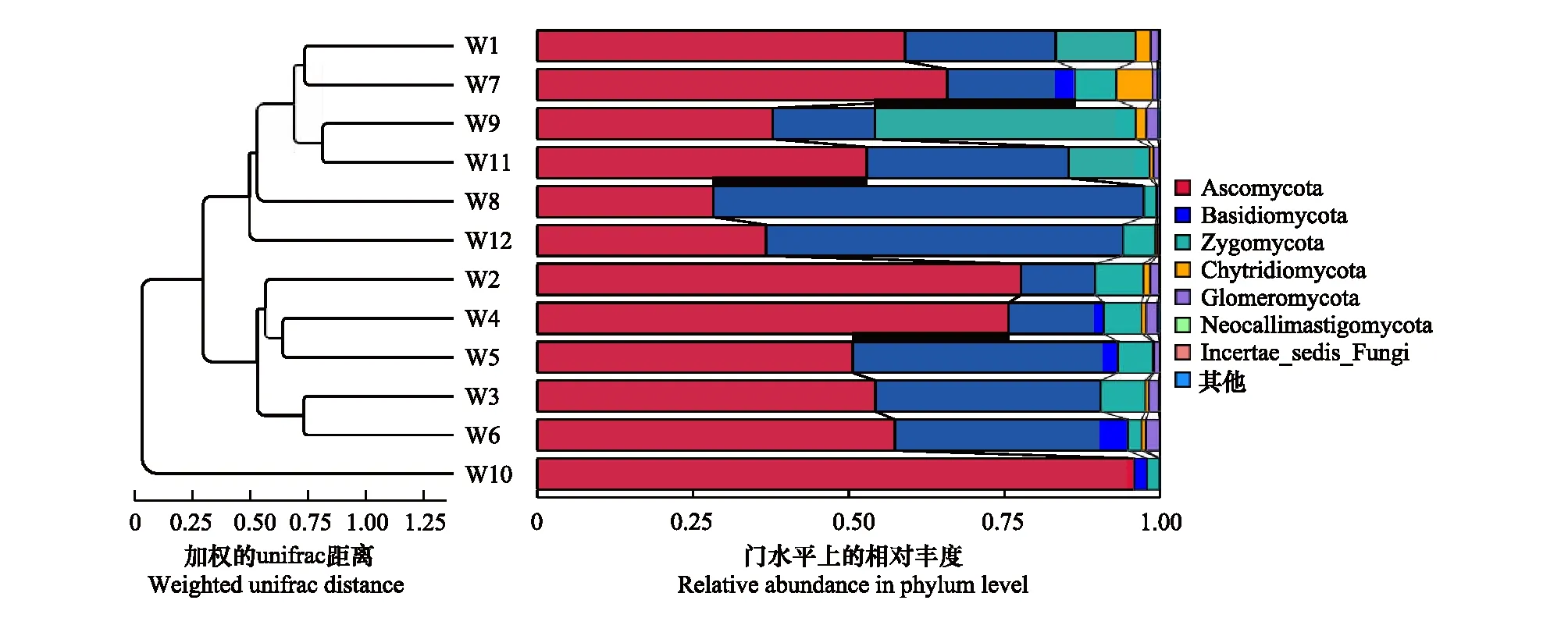
图4 不同植被类型土壤真菌群落结构的UPGMA聚类分析

图5 不同植被类型土壤真菌群落结构的PCoA
2.7 土壤营养元素对真菌群落的影响
将土壤中的10个优势菌属与环境因子进行冗余分析(RDA)可发现(图6),第一轴可以解释所有信息的29.15%,土壤有机碳、全磷、全氮是影响真菌群落的主要因子,几种土壤理化性质呈正相关关系;第二轴解释率为9.81%,主要影响因子为全钾,总解释率达到38.99%,4种环境因子对优势菌属的影响从到到小为:TOC>TP>TN>TK。帚枝霉属(Sarocladium)、Rhytidhysteron、鬼笔属(Lysurus)、Microidium、大环柄菇属(Lepiota),与全钾含量成正相关,与全氮、全磷和有机碳含量呈负相关;微皮伞属(Marasmiellus)、丝盖伞属(Inocybe)、盾盘菌属(Humaria)与土壤全氮、全磷、有机碳含量呈正相关,与全钾含量呈负相关;赤霉属(Gibberella)、镰刀属(Fusarium)与土壤全钾、有机碳含量呈正相关,与全氮、全磷和有机碳含量呈负相关。
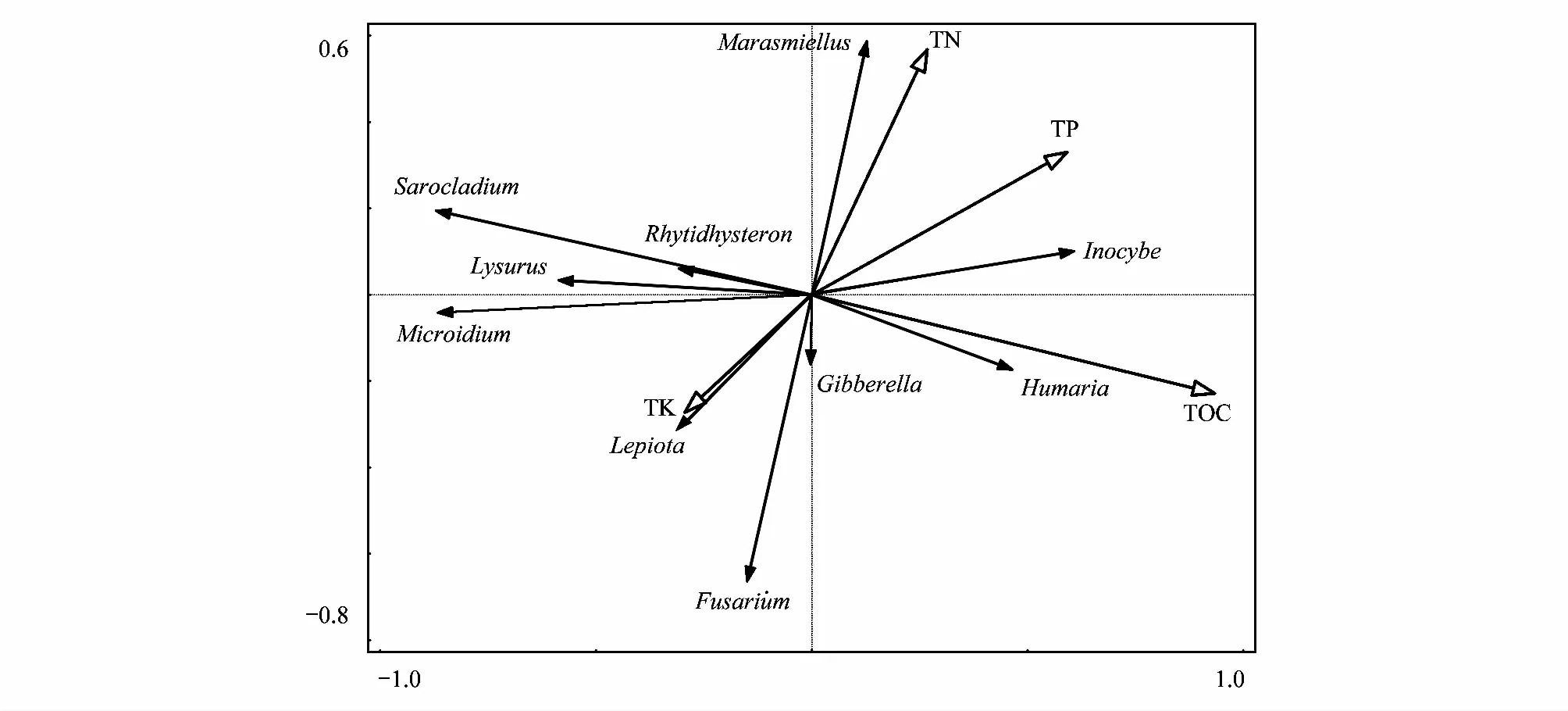
图6 优势菌属与土壤营养元素的冗余分析
3 讨论
3.1 天山不同植被类型对土壤真菌多样性的影响
植物生长过程中从土壤环境吸收水分和营养物质的同时也会释放大量分泌物,种类繁多的分泌物影响根际土壤环境的同时也影响了根际土壤的微生物多样性[13]。植被物种越丰富,其根系分泌物种类越多,同时植被凋落物增加促进养分循环,林下土壤微生物多样性指数也就越高[14-15]。本文中灌丛的OTU数量、物种数目、真菌的多样性和丰富度都多于云杉林,可能是因为几种灌木混杂生长,林下植被多样性要高于纯林的云杉且凋落物种类更为丰富,土壤微生物生存环境较好,微生物存活率较高[16]。云杉作为针叶树种,凋落物中所含诸如木质素、丹宁以及树脂等有机组分不容易被分解,土壤微生物生长繁殖所需的有效可利用底物十分缺乏,这些不利条件将进一步限制云杉土壤微生物的生长和繁殖[17]。与其他研究相同[18],与灌木相反云杉根际土中真菌的多样性和丰富度都低于非根际土且阴坡较高,阳坡次之,林窗最低;阴坡比阳坡更适宜云杉的生长,云杉林下、林窗很少有植物生长,被认为可能是由于根系分泌一些化感物质抑制了其他植物的生长,这些化感物质同时也会对云杉的种子萌发和早期幼苗的定居产生影响。几种灌木的土壤真菌群落多样性之间有一定的差异,但差异较小;云杉中除了林窗根际土,其他土样间真菌群落多样性差异较小,微生物群落呈现出高度的局域生物多样性。云杉的非根际土与灌木根际土的土壤真菌群落结构较为相似,与孙棋棋等人发现真菌群落在同地区林草地间无显著差异相似[19],可能是因为真菌群落具有一定的内稳态。而云杉的根际土和灌木的根际土相比真菌群落差异较大且多样性较少,可能是由于云杉根际富集了特定的真菌群落,且对其他土壤真菌有一定程度的抑制作用。
3.2 天山不同植被类型对土壤真菌群落结构的影响
在森林土壤中,50%—70%的腐殖质来源于根或者与根相关的微生物[20], 根际微生物可影响植物对养分的获取,进而影响植物对生物和非生物胁迫的响应策略以及植物的生产力和多样性[2,21],同时植被类型也显著影响微生物群落的碳源利用能力、群落组成及其活性[22-23]。菌根共生体是自然生态系统中十分重要的生命有机体,陆生维管植物中只有8%为非菌根植物,云杉为天山北坡单一树种,是外生菌根依赖型树种[24],灌木大多数则为丛植菌根型。外生菌根主要隶属于担子囊菌门(Basidiomycota)、子囊菌门(Ascomycota)和接合菌门(Zygomycota)、以担子囊菌最多;丛植菌根隶属于球囊菌门(Glomeromycota)。本文检测到的丰度较高的5门真菌群落分别为子囊菌门(Ascomycota)、担子囊菌门(Basidiomycota)、接合菌门(Zygomycota)、壶菌门(Chytridiomycota)和球囊菌门(Glomeromycota)。辽东山区的红松人工林中,子囊菌门与担子囊菌门占比达25.16%和63.51%[24];青藏高原冻土区垂直剖面中子囊菌门与担子菌门组成比例分别为75.3%和24.7%[25];与呼伦贝尔针茅草原土壤真菌群落结构等以上研究相同[26],天山植被中子囊菌门与担子囊菌门是绝对的优势菌群。它们在土壤养分循环中起着重要作用[27],能够降解木质素和角质素等难分解的物质,担子囊菌主要降解木质素且受植被类型的影响程度较大[28-29]。灌木与云杉在门水平上群落组成差异不大,但丰度存在一定的差异,这与张俊忠等[30]对于东祁连山高寒草地4种不同草地类型土壤真菌群落研究中得出的结论一致。
寄主植物和环境因子对土壤真菌群落有明显影响,土壤理化指标对真菌群落结构的影响中表明,有机碳和全磷对真菌群落结构的影响较为明显,微皮伞属(Marasmiellus)、丝盖伞属(Inocybe)、盾盘菌属(Humaria)赤霉属(Gibberella)、镰刀属(Fusarium)与有机碳含量呈正相关。天山雪岭云杉林中相对丰度较高的属有镰刀属(Fusarium)、赤霉属(Gibberella)、丝盖伞属(Inocybe);灌丛中相对丰度较高的有帚枝霉属(Sarocladium)、鬼笔属(Lysurus)、Microidium;灌丛和云杉林共有的所占比例较多属为赤霉属(Gibberella)、镰刀属(Fusarium)和微皮伞属(Marasmiellus)。镰刀属丰度最高,但一些菌种是主要致病菌[31]。许多镰刀属可以分泌聚酮(polyketides),类萜(terpenoids)和非核糖体肽(nonribosomalpeptides)等特定的植物毒性代谢物或其他生物活性代谢物[32-33],产生如小麦赤霉病、水稻恶苗病、马铃薯块茎干腐病和松树溃疡病等极具破坏性的病害[34],也有一些镰刀菌能产生赤霉素和纤维素酶等物质促进植物生长[13]。丝盖伞属在云杉中丰度较高,是常见的外生菌根,可以与植物形成菌根,促进营养元素吸收,提高植物的抗逆性[35]。被孢霉属可以抑制一些致病菌,在一定程度上还能缓解土壤负面状态对土壤酶活性和作物生长的影响,是一种具有潜在生物防治意义的真菌,有研究发现其可能是内生真菌的常见类群[36],根际土壤中被孢霉属的相对丰度可用来推测根际抵抗根腐病的能力的强弱[37-38]。研究中的菌根真菌较少而某些致病菌占优势,这些菌株可能限制了菌根真菌对植物的侵染,使幼苗难以定殖,从而导致天山云杉的更新问题。丝盖伞属为云杉的优势外生菌根菌属,可为天山雪岭云杉森林的菌根化育苗提供帮助。
4 结论
天山不同植被类型土壤真菌群落中灌木土壤真菌多样性高于云杉,但多样性无显著差异。土壤有机碳和全磷对土壤真菌群落结构影响最大。丝盖伞属、被孢霉属、赤霉属和镰刀属在土壤真菌中丰度较大为优势菌属。
猜你喜欢
纺织学报(2022年8期)2022-08-26
浙江农业科学(2022年7期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
黄河之声(2021年12期)2021-10-25
现代农村科技(2020年7期)2020-12-19
农村实用技术(2020年6期)2020-12-15
花卉(2020年6期)2020-01-07
意林·全彩Color(2019年11期)2019-12-30
天然产物研究与开发(2018年3期)2018-05-07
启迪与智慧·教育版(2018年12期)2018-02-26
