
薄毛海绵杜鹃结实和萌发特性随环境的变化
2021-02-25 06:56:26杨紫玲喻舞阳郑维列王国严李江荣
生态学报 2021年1期
杨紫玲,喻舞阳,郑维列,罗 建,王国严,5,*,卢 杰,李江荣
1 西藏高原森林生态教育部重点实验室, 林芝 860000
2 西藏农牧学院高原生态研究所, 林芝 860000
3 西藏林芝高山森林生态系统国家野外科学观测研究站, 林芝 860000
4 西藏农牧学院资源与环境学院, 林芝 860000
5 成都理工大学生态资源与景观研究所, 成都 610051
杜鹃属(RhododendronL.)植物广布于北半球温带地区,其花色艳丽,具有较高的水土保持和观赏与药用价值[1-3]。全球约有1000种杜鹃花,中国的杜鹃花种类占世界种类的半数以上,除宁夏和新疆以外均有野生杜鹃花种质资源分布,主要集中在西南地区,其中西藏共有野生杜鹃花170种左右,约占世界的1/5,特有种37种,是我国杜鹃花最大的分布中心之一[1,4],是开展杜鹃花生态学研究的理想区域。
薄毛海绵杜鹃(Rhododendronaganniphumvar.schiaopeplum)在藏东南地区大面积连续分布,是色季拉山林线(4320 m)的指示种和优势种[5-7],对林线地区水土保持具有重要意义[8]。作为高山林线优势植物,认识薄毛海绵杜鹃对高山环境特有的适应性[9]以及对不同环境因子的响应特征,是理解该区域林线形成和变化机制的基础。
植物的果实和种子是物种繁殖系统中受遗传控制较强的两个性状,在强大的选择压力之下,会表现出对环境独特的适应性[10-11]。结实特性不仅是植物种群生殖生态学研究的重要内容[10],有助于认识种群在特定环境的繁殖投入等生态策略,也是理解植物种群更新和繁殖适合度的基础[12]。种子作为植物的生活史中唯一有移动力能力的阶段[13],也是种群在特定环境下成功延续和发展的重要生活史阶段[14]。
高山杜鹃多以种子为主要繁殖方式,研究其结实和种子萌发特征对于认识种群的更新策略、对高寒环境的适应机制以及群落演替规律具有重要意义。然而,对西藏高海拔(高山)杜鹃的种实特性研究资料较少[15-16]。本研究选择青藏高原东南林线地区优势植物薄毛海绵杜鹃为对象,研究薄毛海绵杜鹃结实和萌发特性随海拔(4183—4673 m)、坡度、坡向等环境梯度的变化,旨在阐明不同海拔薄毛海绵杜鹃结实和萌发特性与环境之间的关系,认识该种群在藏东南地区的分布和繁殖状况,为高山杜鹃的种质资源保护和气候变化背景下藏东南林线动态预测提供基础资料。
1 材料与方法
1.1 研究地概况与实验材料
研究地位于青藏高原东南部的色季拉山(西藏林芝高山森林生态系统国家野外科学观测研究站附近)。色季拉山系念青唐古拉山向南延伸的余脉,属湿润山地暖温带、半湿润山地温带气候[17],该区域杜鹃花资源丰富,面积可达1000 km2以上[18]。薄毛海绵杜鹃(又称裂毛雪山杜鹃)隶属常绿杜鹃亚属(Subgen.Hymenanthes(Blume)K.Koch)常绿杜鹃组(Section.PonticumG.Don),原产芒康、察隅、米林,果实粗短而直立或略弯,花冠粉红色至白色,是高山林线灌丛优势植物[19]。
2017年10月5日—10日在色季拉山沿海拔梯度设置八个样点(4183、4270、4330、4390、4480、4577、4673、4724 m),在每个海拔带随机选取>5株薄毛海绵杜鹃,共采摘果实数量>100个,带回实验室常温条件(约15 ℃)下自然晾干备用。薄毛海绵杜鹃采种地环境数据详见表1。
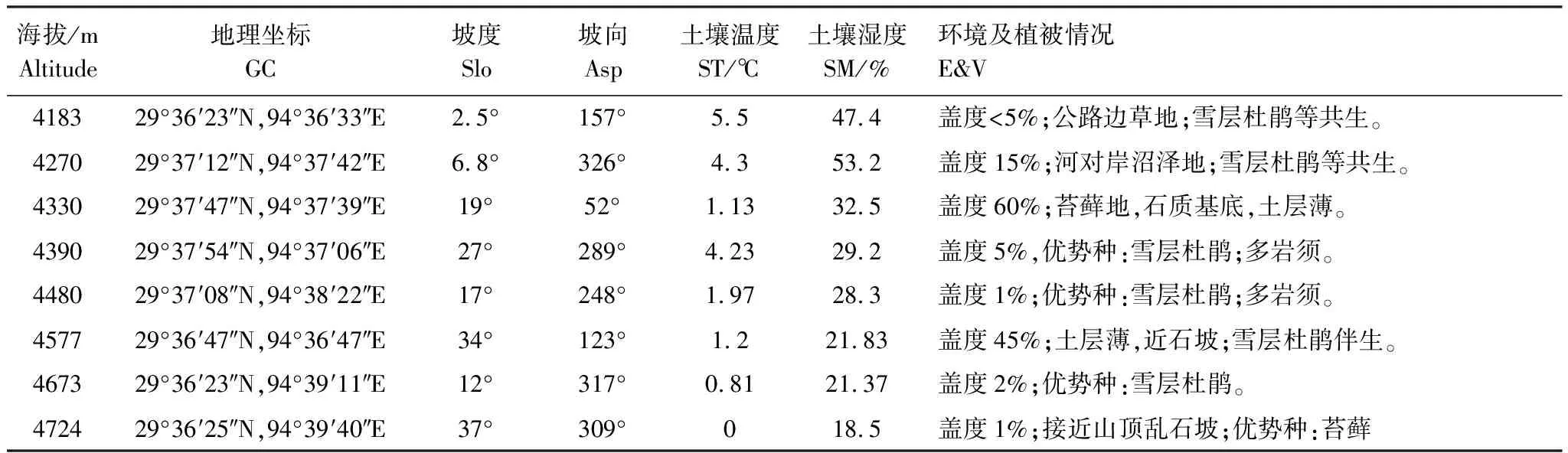
表1 采种地环境数据
1.2 实验方法
1.2.1结实特性实验
将果实根据海拔分为8个组,每组设置4个重复,每个重复随机选取25个果实。用电子天平称量果实的干重;用游标卡尺进行果实的长宽测量;再随机从25个果实中随机选3—5个健康的果实对种子数量进行计数;每个海拔随机取种50粒,分别称重,重复三次,其平均重量乘以20为种子的千粒重。
1.2.2种子萌发特性实验
设置5/1、15/2、25/5 ℃三个温度梯度,每个温度设置光照(12 h光照/12 h黑暗)和黑暗两个处理。3个温度梯度分别代表野外5月和10月(5/1 ℃)、9月(15/2 ℃)、6月至8月(25/5 ℃)。每个海拔选取600粒外观良好的种子,每25粒种子为一组放入垫有双层滤纸(滤纸已用铅笔标号)的培养皿中,4个重复为一个处理。在培养皿中加入适量蒸馏水(约2 mL),每4个培养皿装入一个自封袋密封,黑暗处理的4个培养皿用以两层锡箔纸封窗,之后分别放入3个温度梯度的人工气候培养箱内进行培养。
2017年11月3日将种子放入培养箱培养,随后每7 d观察一次(每周五),持续6周,每次观察时将已经发芽的种子(以胚根突破种皮为标准)挑出培养皿,根据需要补充蒸馏水,保持滤纸湿润、清洁。黑暗处理的种子萌发的观测在暗室中的安全绿光LED灯(中国深圳万科,光谱输出为515 nm—530 nm,吸收峰值为520 nm)下进行。每次记录种子发芽数以及出现的特殊情况(如是否发霉等),最后一次观察时统计未发芽种子数(本研究的目标是检验新成熟种子的萌发能力与环境的关系,因此对未发芽种子不进行是否有活力试验)。
1.2.3数据分析
采用广义线性模型(GLMs)检验各环境因子及其交互作用对种实性状以及萌发率的影响(对所有模型的主因子和交互因子执行递归法模型筛选,直到模型仅包含显著的主因子/交互作用(P<0.05)),用卡方(X2)检验显著性。其中萌发率服从二项分布,果实重量、长、宽、长宽比、种子千粒重服从正态(高斯)分布,种子数量服从泊松分布。利用Tukey检验比较不同海拔果实和种子性状的平均值之间的显著差异性。
数据分析在R(3.5.1)中进行。二项分布误差和广义线性模型基于lme4包、MASS包、lmerTest包、plyr包、car包进行;Tukey检验基于multcomp包进行。文中和图形中的数据采用原始数据的均值(±se)展示。
2 结果分析
2.1 果实与种子表型性状
薄毛海绵杜鹃的果实为蒴果,干燥后呈棕褐色,因长宽比的差异,形状由椭圆形至倒卵形,果柄朝上。不同海拔的薄毛海绵杜鹃果实和种子在数量性状上存在显著差异(P<0.05,种子千粒重除外)。种子细小扁平为粉末状,颜色随海拔的变化由黑棕色(4183—4480 m)过渡至黄色(4527—4724 m)。果实干重(25个果实)为(2.13±0.18)g—(4.03±0.06)g,海拔4724 m样点果实最重,海拔4183 m样点果实最轻。果实长度为(10.50±0.25)mm—(14.91±0.28)mm。果实宽度为(4.52±0.09)mm—(6.22±0.16)mm。4183、4673、4724 m的果实长宽比均在2.30以上,果实外形较瘦长,其他几个海拔的杜鹃果实长宽比较小,长宽比均在2.30以下,果实外形较肥硕。单果种子数最少为(73.50±7.69)粒(海拔4183m样点处),单果种子个数最多为(435.25±50.8)粒(海拔4673 m处)。种子千粒重为(0.05±0.01)g—(0.10±0.01)g,4183—4577 m种子千粒重差异不大,而海拔4673 m和4724 m的种子千粒重较小,分别为(0.05±0.005)g、(0.06±0.01)g。
2.2 果实重量随环境因子的变化
海拔、坡度、土壤湿度显著影响果实重量(P<0.05,表3)。果实重量随着海拔、坡度升高而增加,随土壤湿度增大果实重量减小(图1),果实重量最小值(2.13±0.18)g出现在海拔4183 m、坡度较小(2.5°)和土壤湿度47.4%环境条件下,而果实重量最大值(4.03±0.06)g在海拔4700 m、坡度37°和土壤湿度18.5%环境条件下(图1)。

图1 果实重量随海拔、坡度、土壤湿度的变化
2.3 果实长度、宽度、长宽比随环境因子的变化
海拔、坡向和坡度交互作用显著影响果实长度(P<0.01,表3),其他环境因子间无显著交互作用(P>0.05)。果实长度随着海拔升高而增加,最大值(14.91±0.28)mm出现于海拔4724 m处;最小值(10.50±0.25)mm出现于海拔4270 m处(图2)。
坡度、坡向以及土壤湿度显著影响果实宽度(P<0.05,表3)。果实的宽度随坡度的增大而增加,随坡向和土壤湿度的增大而减小,最大值(6.22±0.16)mm出现于海拔4724 m处;最小值(4.52±0.09)mm出现于海拔4183 m处(图2)。

表3 环境因子对薄毛海绵杜鹃单果个数、种子千粒重以及对果实的重量、长度、宽度与长宽比的影响
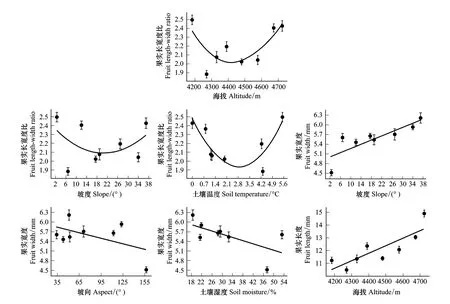
图2 果实长度、宽度、长宽比随显著相关环境因子的变化
而海拔、坡度、土壤温度、坡向和坡度交互作用显著影响长宽比(P<0.05,表3)。果实长宽比随海拔、坡度、土壤温度的增加而呈现两端大中间小的“U”型变化,最大值(2.50±0.05)mm出现于海拔4183 m处;最小值(1.88±0.04)mm出现于海拔4270 m处(图2)。
2.4 单果种子数随环境因子的变化规律
除土壤温度(P=0.725)外,海拔、坡度、坡向、土壤湿度和坡向与坡度的交互作用显著影响单果种子数(P<0.05,表3)。单果种子数随海拔升高而增加;随坡向增大而减少,且呈现阴坡(315°—45°)>半阴坡(45°—90°和270°—315°)>半阳坡(90°—135°和225°—270°)>阳坡(135°—225°)的规律;随坡度和土壤湿度的增大单果种子数呈先减少后增加的趋势。单果种子数最大值为(435.25±50.8)粒,出现于海拔4673 m处;最小值为(73.50±7.69)粒,出现于海拔4183 m处(图3)。
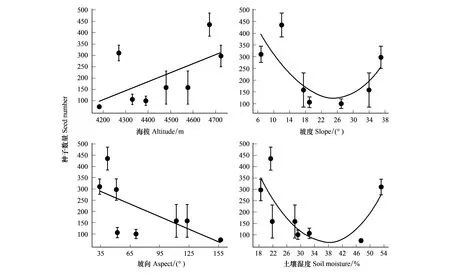
图3 单果种子数随海拔、坡度、坡向、土壤湿度的变化
2.5 种子千粒重随环境因子的变化
坡度(P=0.068)、坡向(P=0.779)、土壤温度(P=0.308)、土壤湿度(P=0.452)对种子千粒重无显著影响,只有海拔因子显著影响种子的千粒重(P<0.05,表3)。海拔4390 m种子的千粒重最大,为(0.10±0.01)g;海拔4673 m最小,为(0.05±0.005)g,且总体呈随海拔的增大而千粒重减小的趋势(图4)。
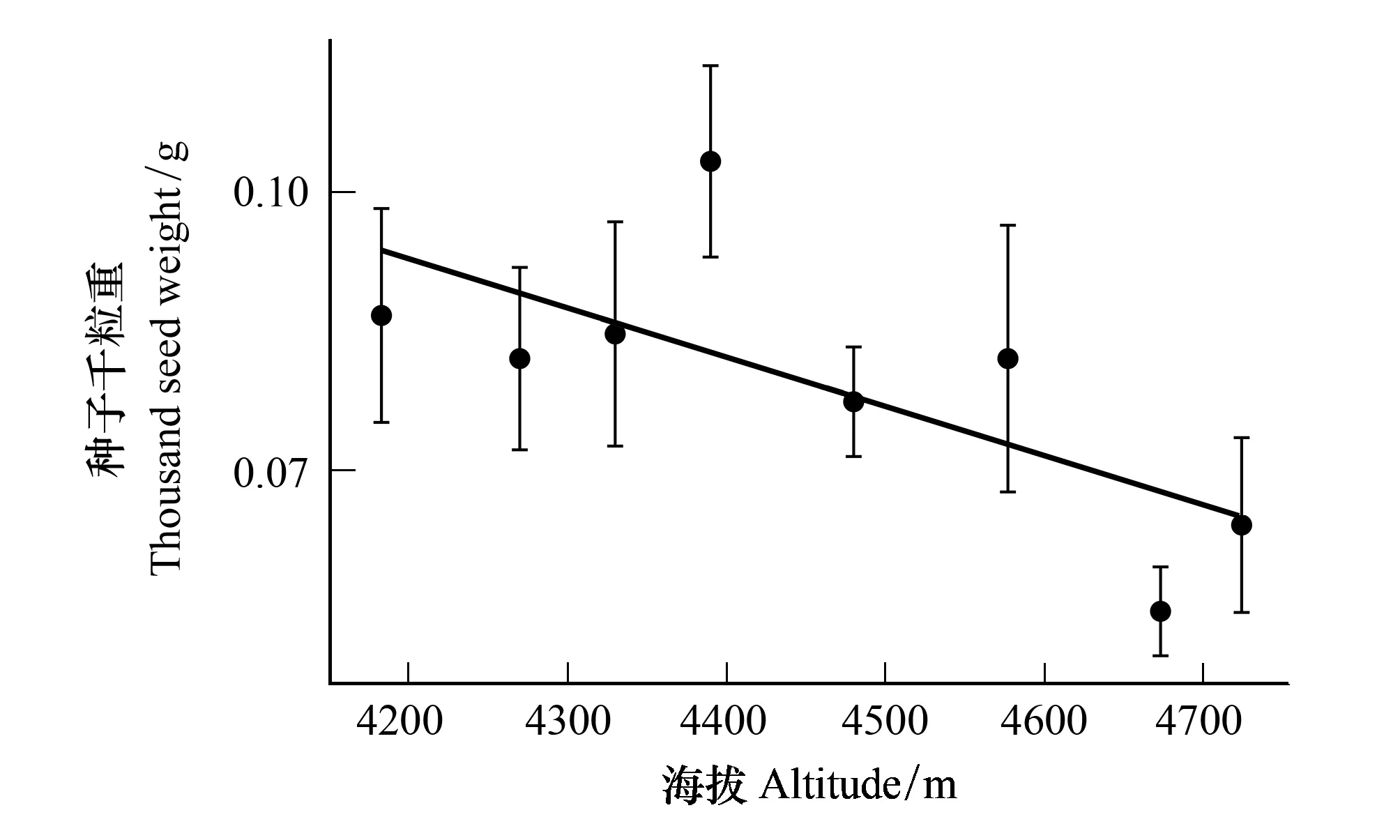
图4 种子千粒重随海拔的变化
2.6 种子萌发对母体环境和培养光照、温度的响应
除海拔显著影响薄毛海绵杜鹃的种子萌发(P<0.01)外,其他因子对薄毛海绵杜鹃的种子萌发无显著影响(P>0.05),种子萌发率随着海拔的升高呈现出两端低中间高的倒“钟型”变化趋势(图5)。
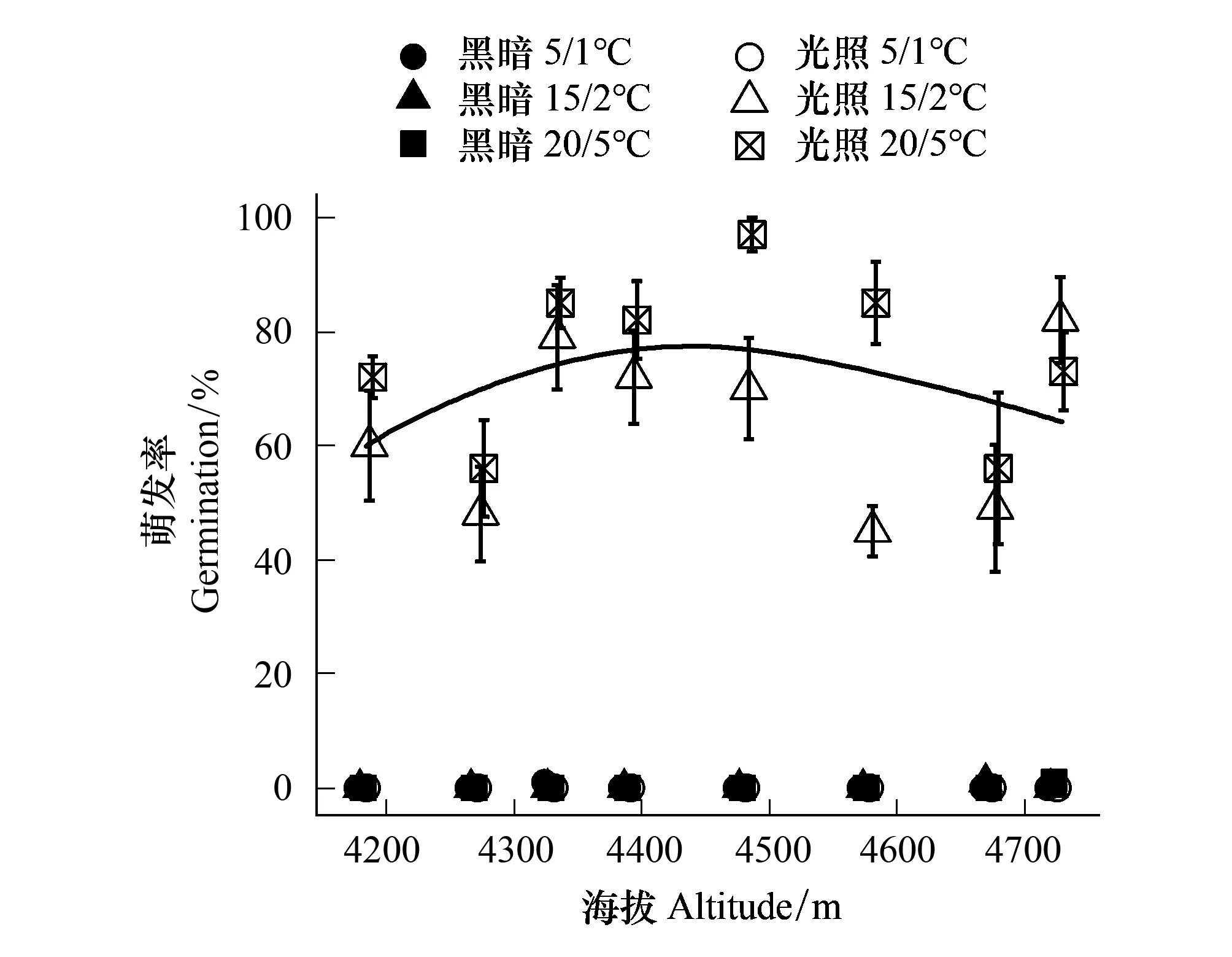
图5 海拔、温度和光照海拔薄毛海绵杜鹃种子发芽率的影响
种子的萌发实验在模拟日光照(12 h/12 h)和持续黑暗两种条件下进行,结果显示薄毛海绵杜鹃的种子在黑暗条件下发芽率较低,且在有光条件下,第二周开始出现萌发,在第六周时达到46.2%。
在5/1 ℃条件下8个海拔的种子均不萌发;在15/2 ℃条件下,海拔4183m的种子从第4周开始出现萌发,且随着海拔的升高,种子起始萌发时间缩短为3周;25/5 ℃条件下,种子起始萌发时间在高海拔4673 m、4724 m和低海拔4183 m时均为3周,其余中海拔均为从第2周开始萌发;海拔为4673 m和4724 m时,两个可萌发温度(15/2 ℃、25/5 ℃)对种子萌发起始时间没有影响。
除了海拔4724m的种子外,其他海拔的种子萌发率在20/5 ℃下的比15/2 ℃下高,且相同培养温度条件下海拔间的种子萌发率存在显著差异(P<0.05)。在25/5 ℃条件下,最高种子萌发率出现在海拔4480 m(97.0%)处,最低出现在海拔4270 m和4673 m(56.0%)处,其中在海拔4330、4390、4480、4577 m处种子萌发率均高于80.0%;在15/2 ℃条件下,种子萌发率最高出现在海拔4724 m(81.0%)处,最低出现在海拔4577 m(45.0%)处,其中萌发率在海拔4270 m(48.0%)—4330 m(79.0%)与海拔4673 m(49.0%)—4724 m(81.0%)之间存在显著差异。
3 讨论
3.1 结实特性
不同环境条件下薄毛海绵杜鹃的果实重量、形状之间存在显著差异(表2),随着海拔升高和坡度增加,果实更重更大;坡向越小、土壤湿度越低时果实更重更大,最高海拔(4724 m)所采果实最大最重,长(14.91±0.28)mm、宽(6.22±0.16)mm、重(4.03±0.06)g。即在高海拔、坡度大、坡向小、土壤湿度低的区域,薄毛海绵杜鹃果实干重较重、长宽也较大。单从果实的重量、形状来看,生活在较强胁迫(高海拔)条件下的薄毛海绵杜鹃繁殖投入比较大。恶劣环境条件下,常绿植物在长期进化过程中形成了加大繁殖投入的生存策略,即将同化的资源更多地分配到繁殖器官(果实和种子)中[20],进而可以产生更多的种子来完成繁殖过程。薄毛海绵杜鹃在高海拔胁迫环境(如气温低,土壤湿度低、土壤温度低等)条件,通过提高自身的繁殖能力来结又大又重的果从而产生更多的种子使种群得到稳定发展。
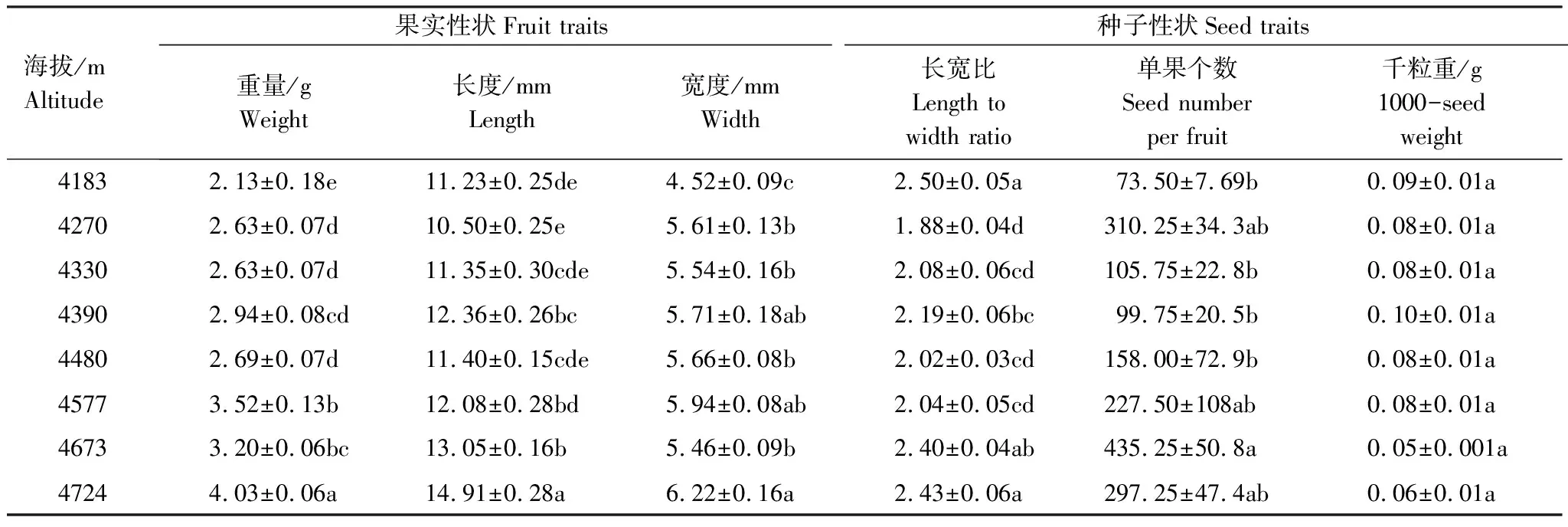
表2 不同海拔样点的果实和种子性状
海拔越高种子的千粒重越小,单果种子数也越多(P<0.05,图3,图4)。可能与高海拔生长季更短有关,导致种子生长所需的营养生长和发育时间减少[21],陈学林研究[22]也发现较短的生长季使种群更趋向于产生重量小的种子,但本研究中在高海拔较大繁殖分配策略的驱使下,杜鹃以更大量的种子来弥补和权衡。另外,小种子较大种子而言有更远的传播距离[23-25],这可能是高山植物应对环境胁迫、维持种群稳定发展的对策,从而保证有足够的种子数量、较强的种子扩散传播能力以及增加种群更新成功几率。
坡向是影响薄毛海绵杜鹃种子数量的主要环境因子(坡向和坡度之间有较强的交互作用,P<0.05),除阴坡的单个果实种子数显著大于阳坡外,还表现出坡度较大或较小时的单果种子数显著大于坡度适中时单果种子数的规律。这与阴坡的水分、土壤养分条件要比阳坡好有关,生活在阴坡的薄毛海绵杜鹃具有更强的繁殖能力。该结果也与郭其强等人调查[26]所得的分布规律一致。
在土壤湿度较高或较低时虽然单果种子数较多(P<0.05,图3),但结合萌发实验结果来看较高或较低的土壤湿度条件产生的种子萌发力低。低海拔4270 m的薄毛海绵杜鹃种群生活在河边沼泽地,水分优越(土壤湿度53.2%),单果种子数可达(310.25±34.3)粒,但种子的萌发率却较低(< 60%)。而土壤湿度在28.3%—32.5%时单果种子数较少(99.75±20.5)粒—(158±72.9)粒,但种子的萌发力却较高(> 60%)。可能因为种子在成熟的过程中如果受到水分胁迫(湿度过低或过高),导致种子的活力较低,萌发率不高[27],故生长在土壤湿度较适中环境里的薄毛海绵杜鹃种子的萌发能力较高,种子更为健康,种群发展更稳定。同时,生活在土壤湿度较高或较低的薄毛海绵杜鹃通过加大繁殖投入(即产生大量的种子)的调整使适合度最大化[28-29]以保障幼苗能成功建植,来维持种群的生存。
3.2 萌发特性
薄毛海绵杜鹃种子的发芽能力与母体植株海拔密切相关。海拔作为间接影响因子,影响了温度从而影响了薄毛海绵杜鹃种子的萌发能力。同高海拔、低海拔的种子相比,中间海拔的种子萌发率更高、萌发速率更快,可能是由于植物个体对低温的适应性导致了同种之间种子萌发能力的差异。在西藏色季拉山,海拔4300—4500 m是薄毛海绵杜鹃的主要分布范围[6],从本研究结果来看,不同温度条件下所产种子的萌发能力是决定薄毛海绵杜鹃分布范围的重要因素之一。
种子的萌发要求种子必须有活力且已打破休眠,其他环境需求如温度、光照等也是影响种子萌发的重要因素[30-34]。而种子的萌发特性恰恰反映了物种对生存环境的适应能力[35]。
光是植物的生命活动中必不可少的因子之一[32],本研究中,种子经过了六周的培养,薄毛海绵杜鹃种子在光照条件下能发芽,萌发率为(46.25±0.37)%,而黑暗条件下种子极少萌发(萌发率<1%)。张茂林[36]也发现光照是影响杜鹃花种子萌发的重要环境因素。在温带的高海拔地区,由于光照受到冬季积雪和夏季太阳辐射的强烈影响,光是反映种子在土壤中的位置/雪埋深度的空间信号[37],光照成为影响薄毛海绵杜鹃种子萌发的一个重要环境因子。薄毛海绵杜鹃的蒴果中种子数量多,但种子个体呈粉末状,Milberg[38]提出种子萌发对光的需求与种子质量共同进化,小种子的萌发更加依赖光照。薄毛海绵杜鹃的种子属于需光性种子,对光的响应有利于种子等待冬春季融雪或土壤干扰后,接近地面(有光照)再萌发,降低小种子在冬季较深雪埋环境或较深的土埋条件下萌发但出土失败的概率,有利于储存能量有限的小种子的幼苗建成。
温度显著影响薄毛海绵杜鹃种子萌发率和萌发速率,除在5/1 ℃条件下8个海拔的种子均不萌发,而在15/2 ℃(萌发率63.0%)和25/5 ℃(萌发率75.8%)条件下种子均有大量萌发,且适当的高温可加快种子的萌发进程。薄毛海绵杜鹃的萌发温度需求与大白杜鹃(R.decorum)和秀雅杜鹃(R.concinnum)的生长需求相似[39-40],其最适宜的生长温度为 15—25 ℃,在 5 ℃条件生长趋于停滞,因而适宜在比较温暖的春夏季萌发。薄毛海绵杜鹃种子萌发对温度的响应,调控种子在高寒地区相对高温条件下萌发(春夏季),而在低温条件下(秋冬季)不萌发,这有利于高寒植物种子避免在种子成熟的晚秋、冬季寒冷条件下萌发,从而保证幼苗的存活,是种子萌发阶段对高寒环境的适应。
4 结论
本研究通过调查和实验阐明了海拔梯度上(4183—4673 m)薄毛海绵杜鹃结实和萌发特性与环境之间的关系,主要结论如下:
(1)在高海拔低温胁迫环境下,薄毛海绵杜鹃的生殖投入加大,果实大且质量重,而种子数量多但质量轻,该特征有利于种子的扩散和种群在胁迫环境下繁衍成功;
(2)坡向是影响单果种子数量的主要环境因子,阴坡单果种子数显著大于阳坡,生活在阴坡的薄毛海绵杜鹃的繁衍能力更强;
(3)较高和较低土壤湿度都会影响薄毛海绵杜鹃的种子质量,中等土壤湿度(28.3%—32.5%)薄毛海绵杜鹃种子萌发能力最强;
(4)薄毛海绵杜鹃的种子是需光性种子且适宜在温暖的春夏季萌发,体现出种子萌发特性对高寒地区萌发季节的筛选功能,是一种对高寒地区生长季短、春季雪埋等特殊生境的适应策略。
薄毛海绵杜鹃作为高山林线的一种指示灌丛,海拔4330—4480 m的薄毛海绵杜鹃与其他海拔的薄毛海绵杜鹃相比,无论是结实特性或萌发特性其性状指数均较高,可知海拔4330—4480 m为薄毛海绵杜鹃种群的最适分布范围。全球变化背景下,未来藏东南的暖湿化可能会导致薄毛海绵杜鹃种群的分布区向高海拔扩张。
猜你喜欢
中国农业科学(2022年13期)2022-07-26 09:55:28
天津农林科技(2020年3期)2020-08-13 03:17:10
测绘学报(2019年11期)2019-11-20 01:31:42
电子制作(2019年15期)2019-08-27 01:12:12
考试周刊(2016年11期)2016-03-17 05:11:15
高原山地气象研究(2016年4期)2016-02-28 13:53:39
高原山地气象研究(2016年4期)2016-02-28 13:53:37
安徽农业科学(2015年25期)2015-12-22 06:21:16
土壤与作物(2015年3期)2015-12-08 00:46:55
植物营养与肥料学报(2012年3期)2012-10-26 02:54:52
