
Effects of temperature on photosynthetic performance and nitrate reductase activity in vivo assay in Gracilariopsis lemaneiformis (Rhodophyta)*
2021-02-22 02:00ZhihaiZHONGZhengyiLIULongchuanZHUANGWanlinSONGWeizhouCHEN
Zhihai ZHONG , Zhengyi LIU, Longchuan ZHUANG , Wanlin SONG, Weizhou CHEN ,
1 Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, China
2 Shantou University, Shantou 515063, China
Abstract Gracilariopsis lemaneiformis is an economically-valued species and widely cultured in China at present. After being acclimated to different growth temperatures (15, 20, 25, and 30 °C) for 7 days, the relative growth rate (RGR), nitrate reductase activity, soluble protein content and chlorophyll a fluorescence of G. lemaneiformis were examined. Results show that RGR was markedly affected by temperature especially at 20 °C at which G. lemaneiformis exhibited the highest effective quantum yield of PSII [Y(II)] and lightsaturated electron transport rate (ETR max), but the lowest non-photochemical quenching. Irrespective of growth temperature, the nitrate reductase activity increased with the incubation temperature from 15 to 30 °C. In addition, the greatest nitrate reductase activity was found in the thalli grown at 20 °C. The value of temperature coeき cient Q10 of alga cultured in 15 °C was the greatest among those of other temperatures tested. Results indicate that the optimum temperature for nitrate reductase synthesis was relatively lower than that for nitrate reductase activity, and the relationship among growth, photosynthesis, and nitrate reductase activity showed that the optimum temperature for activity of nitrate reductase in vivo assay should be the same to the optimal growth temperature.
Keyword: chlorophyll a fluorescence; Gracilariopsis lemaneiformis; growth; nitrate reductase activity
1 INTRODUCTION
Nitrate assimilation starts when alga takes up nitrate from the external medium. Nitrate reductase (EC 1.6.6.1) reduces nitrate to nitrite by using NAD(P)H as the electron donor (Dovis et al., 2014). Nitrite is then transported into the chloroplast, and subsequently reduced to ammonium, in which nitrite reductase (NiR) uses reduced ferredoxin as an electron donor (Chow et al., 2007; González-Galisteo et al., 2019). The overall rate of nitrate assimilation is often limited by the first reduction step of nitrate reductase activity. This step serves as a key point in nitrogen metabolism (Crawford and Arst, 1993; Campbell, 1999), and can be used as an index to nitrate assimilation in the field (Collos and Slawyk, 1977).
Nitrate reductase activity and photosynthesis are regulated by various environmental factors, including temperature, light, nitrate, iron, and other regulators of algal growth (Eppley et al., 1970; Lapointe et al., 1984; Gao et al., 1992; Young et al., 2007; Chen et al., 2018). Among them, temperature is a key physiological factor on the algal growth, distribution, and reproduction by affecting the sensitivity of the main cellular components (proteins and membranes). The temperature responses of species involve mainly three types: genetic adaptation (in thousands of millions of years), phenotypic acclimation (in hours to days), and short-term physiological regulation (in seconds to minutes). Besides, temperature is also considered to function by regulating C and N assimilation strongly associated with the photosynthesis and enzyme activities (Berry and Bjorkman, 1980; Gao et al., 2017, 2018). Furthermore, previous studies have pointed out that the metabolism of photosynthetic products is closely associated with nitrate reduction in photosynthetic tissues (Gao et al., 1992). The reason is that nitrite reduction needs reducing power and energy provided by photosynthesis (Chow et al., 2013; Varela et al., 2018). However, there are few studies about the regulation of temperature on photosynthesis and nitrate reductase activity by phenotypic acclimation.
In vitro and in vivo assays of nitrate reductase activity are two methods to measure nitrate reductase activity. The in vitro assay first ruptures the cells, then extracts and stabilizes enzyme, and at last tests the enzyme (Dovis et al., 2014). The in vivo assay to increase the membrane permeability, increase the rate of substrate (NO3ˉ), NADH (nicotinamide adenine dinucleotide (NAD) + hydrogen (H)), and reaction production (NO2ˉ) in and out of the cells. The in vivo assay is faster and easier, and the more important is that it represents nitrate reductase activity with the current level of cellular NADH (Chow et al., 2004; Dovis et al., 2014). Previous studies have mainly concentrated on the in vitro assay (Chow et al., 2001, 2004; Vona et al., 2004), but in vivo assay is scarcely discussed (Corzo and Niell, 1991). Only few papers concerned the optimum temperature for the nitrate reductase activity assay in vivo (Corzo and Niell, 1991; Zou, 2005; Teichberg et al., 2007; Cabello-Pasini et al., 2011; Chen et al., 2015), most of those studies reported that the incubation temperature for the in vivo assay should be 30 °C or room temperature, regardless of the species and the actual physiology of the alga in the field. When nitrate reductase is used as an index of the nitrate assimilation, the optimum temperature for the in vivo assay and the relationship between photosynthesis and nitrate reductase activity are important for evaluating C and N assimilation (Kristiansen, 1983).
Gracilariopsis lemaneiformis (Rhodophyta) is an economic macroalgae cultured in a large-scale in China for providing quality raw material for agar industry and feed for abalone aquaculture (Yu and Yang, 2008; Gu et al., 2017; Chen et al., 2018; Liu et al., 2019). By absorbing and utilizing nutrients from the seawater, G. lemaneiformis acts as ideal biofilters to control eutrophication, and improve the health and stability of the marine ecosystem (Yang et al., 2005). As a result, much attention has been paid to the aquaculture techniques and the ecophysiology of this alga (Yang et al., 2006). However, little research focused on the effects of temperature on nitrate reductase activity and photosynthesis in G. lemaneiformis. In this study, the relationship among photosynthesis, growth, content of soluble protein, and nitrate reductase activity assay in vivo were explored and the optimum temperature for in vivo assay of nitrate reductase activity was determined.
2 MATERIAL AND METHOD
2.1 Plant material
Gracilariopsis lemaneiformis was sampled from Shen’ao Bay (23.46°N, 117.09°E), Nan’ao Island, Shantou, China. Samples were kept at 5 °C, and transported to the laboratory in 4 h. The algae were then stored in a glass aquarium tank containing filtered natural seawater (salinity: 28, temperature: 20 °C) under irradiance of 120 μmol photons/(m2·s), and in 12 h L:12 h D photoperiod scheme for 3 days. Healthy thalli were selected for subsequent experiments.
2.2 Experimental design
Thalli of approximately 4 g fresh weight were placed in flasks containing 1-L sterile natural seawater (salinity: 28; NO3ˉ: 17.22 μmol/L; NO2ˉ: 1.62 μmol/L; NH4+: 1.92 μmol/L; PO43ˉ: 0.29 μmol/L) enriched with 100 μmol/L NaNO3and 10 μmol/L NaH2PO4. The irradiance was 120 μmol photons/(m2·s) and 12 h L: 12 h D photoperiod scheme. Triplicate cultures were grown at four different temperatures (15, 20, 25, and 30 °C), and culture medium was renewed every day. Algae remained in culture for 7 days prior to experimental work.
2.3 Relative growth rate (RGR)
The fresh weight of the alga was measured once a day. The relative growth rate (RGR) was calculated as follows: RGR=ln( Wt/ W0)/ t, where W0is the initial fresh weight and Wtis the final fresh weight after t days. Before weighting the algae, samples were softly blotted using filter paper to remove excess water.
2.4 Determination of nitrate reductase activity
The nitrate reductase activity assay method was modified according to the in vivo method described by Corzo and Niell (1991). At the end of the culturing period (7 days), when illuminated for 4 h (Lopes et al., 1997), G. lemaneiformis samples were cut into 3-cm-long segments by scissors, then incubated in culture seawater for 1 h to minimize the cutting damage (Zou, 2005). Approximately 0.2 g of healthy algae were selected at random (Chen et al., 2015), and then put into test tubes containing 5 mL of the reactive medium (pre-cooled at 15, 20, 25, and 30 °C, individually). The reactive medium was made using 0.1 mol/L pH 7.9 phosphate buffer, 1 mmol/L EDTA, 0.1% 1-propanol, 300 μmol/L NaNO3and 10 μmol/L glucose. Subsequently, the medium was infused with N2gas for approximately 2 min to remove oxygen to prevent nitrite from being oxidized to nitrate, medium was then sealed and wrapped using aluminium foil before a 2-h incubation at 15, 20, 25, and 30 °C. After the incubation, the reaction was completed by removing the thalli from the reactive medium. Approximately 1 mL of the resulting medium was then added to a mixture of 1 mL of 1%w/v sulphanilamide and 1 mL of 0.2% w/v n-(1-napthyl) ethylenediamine dihydrochloride. The absorbance of the medium was then determined at 543 nm. Nitrite concentration in the medium was calculated using a standard curve. The nitrate reductase activity was expressed in μmol/(NO2·h·g FW).
The temperature coeき cient Q10 was used to analyze the relationship between temperature and nitrate reductase activity (Rasmusson et al., 2019), and values of Q10 were calculated across temperature intervals (15-30 °C) as: Q10=( V2/ V1)10/(T2-T1), where V1and V2are nitrate reductase activity at different temperatures, T1and T2(in °C).
2.5 Soluble protein determination
Soluble protein was extracted by grinding 0.1-g (fresh weight) thalli in 0.1-mol/L phosphate buffer (pH=7.0) in a mortar on ice. Cell debris was removed by centrifuging for 15 min at 5 000 r/min in 4 °C, and the samples were then assayed according to the modified method of the binding of Coomassie Brilliant Blue G-250 (Bradford, 1976; Read and Northcote, 1981). Approximately 0.1 mL of extract was obtained into the 5 mL of mixture (Coomassie Brilliant Blue G-250) and the absorbance was determined at 595 nm.
The mixture: 100 mg of Coomassie Brilliant Blue G-250 was dissolved in 50 mL of 95% ethanol, and then added with 100 mL of 85% (w/v) phosphoric acid. The solution was diluted to a final volume of 1 L using distilled water.
2.6 Chlorophyll a fluorescence parameters
Chlorophyll a fluorescence parameters were determined using a pulse amplitude modulated fluorescence monitoring system (Maxi-Imagine-PAM, Heinz Walz, Effeltrich, Germany). Samples were placed in dark at different incubation temperatures (15, 20, 25, and 30 °C) for 10 min before beginning the measurements. The effective quantum yield of PSII [Y(II)] is Y(II)=( Fm′- F0)/ Fm′ (Genty et al., 1989), and the non-photochemical quenching (NPQ) is NPQ=( Fm- Fm′)/ Fm′, where Fmis the maximal fluorescence induced by a saturation pulse from a dark adapted sample, F0is the minimal fluorescence level measured at measuring light the low frequency, and Fm′ is the maximal fluorescence level induced by a saturation pulse from algae in active light (111 μmol photons/(m2·s)).
The rapid light curves (RLCs) can be obtained by a series of 20 s light exposures with increasing irradiance (1, 21, 56, 111, 186, 281, 336, 396, 461, 531, and 611 μmol photons/(m2·s)). The parameters of the RLCs were calculated following the photoinhibiton (Eilers and Peeters, 1988) models as follows:
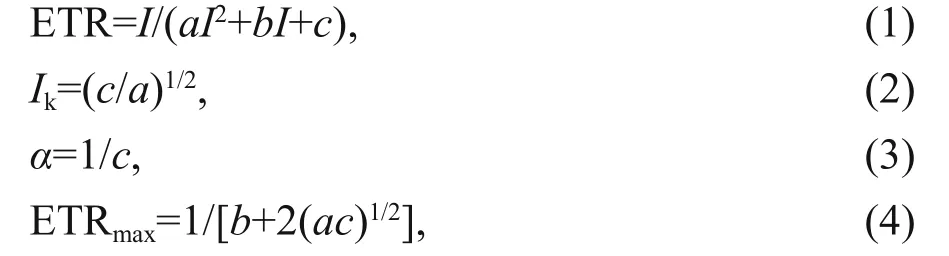
where ETR is electron transport rate, ETRmaxis the light-saturated electron transport rate, α is the electron transport eき ciency, I is the incident irradiance, Ikis saturated irradiance, and a, b, and c are the adjustment parameters.
2.7 Data analysis
ETR was turned into a C-fixation rate according to Silva and Santos (2004), and the nitrate reductase activity was changed to N-incorporation rate according to Collos and Slawyk (1977).
Significance among treatments was tested using the one-way analysis of variance (one-way ANOVA) in SPSS (Version 19). The significant level was set at 0.05. All data were expressed as the mean ± standard deviation (SD, n=3).
3 RESULT
3.1 Growth
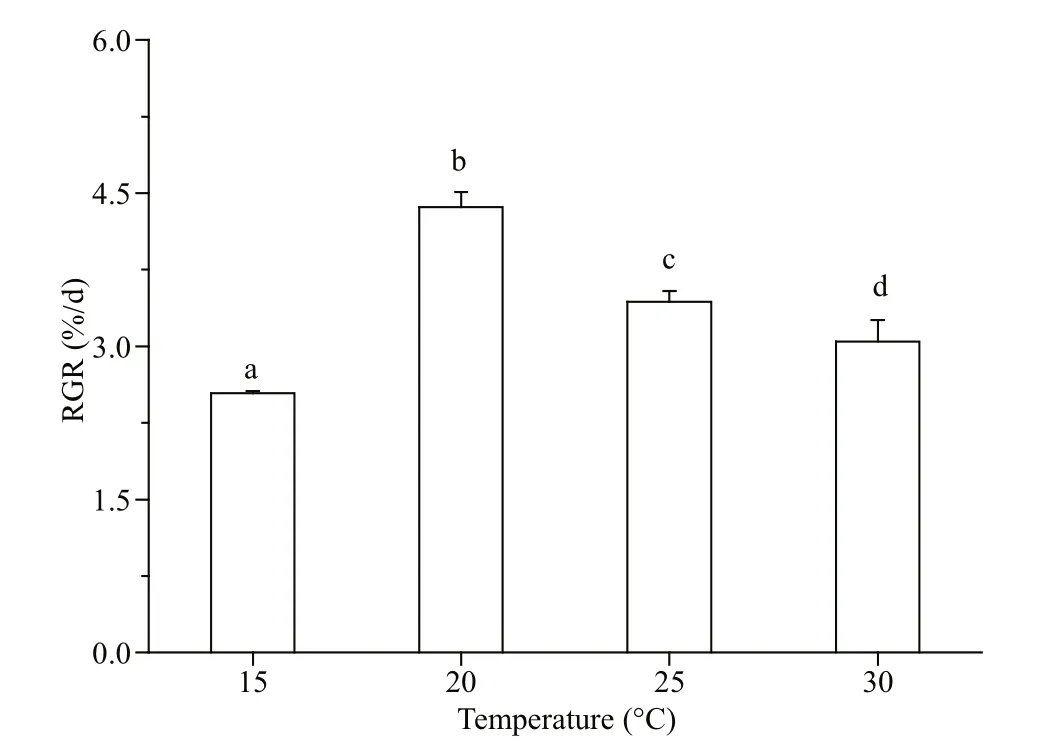
Fig.1 The relative growth rate (RGR) of G. lemaneiformis grown at temperatures of 15, 20, 25, and 30 °C
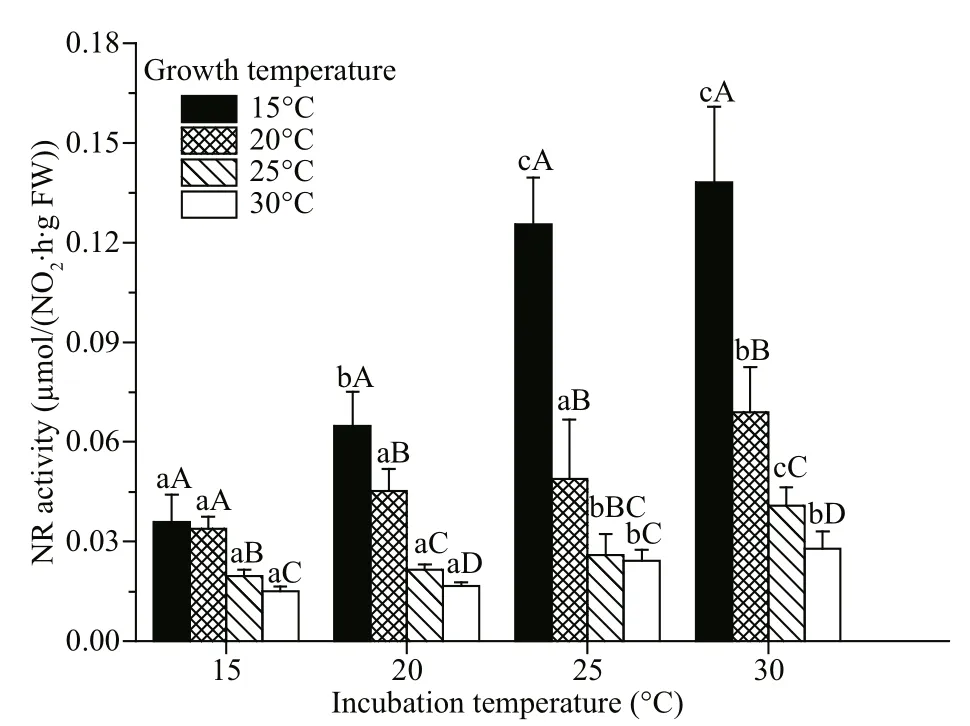
Fig.2 The nitrate reductase activity of G. lemaneiformis determined under different incubation temperature
Gracilariopsis lemaneiformis was cultured for 7 days at 15, 20, 25, and 30 °C, separately. The maximum RGR (4.36%/d) ( P <0.05) occurred at 20 °C. The RGR decreased when the temperature was higher than 20 °C ( P <0.05) (25 °C: 3.43%/d; 30 °C: 3.05%/d; Fig.1). The appropriate temperature for algal growth was between 15 and 25 °C, with the optimum temperature being 20 °C.
3.2 nitrate reductase activity and soluble protein contents
The nitrate reductase activity was determined at different incubation temperatures (15, 20, 25, and 30 °C) and the instantaneous responses of nitrate reductase activity to temperature were detected.Figure 2 illustrates the variation of nitrate reductase activity as a function of temperature. Irrespective of growth temperature, the maximum nitrate reductase activity of algae occurred at 30 °C. Maximum nitrate reductase activity occurred at 15 °C, and decreased with increasing of temperature (Fig.2). After comparing nitrate reductase activity determined at the growth temperature, we found that maximum nitrate reductase activity occurred at 20 °C (Table 1), which is in accordance with the results of algal growth (the maximum RGR occurred at 20 °C).
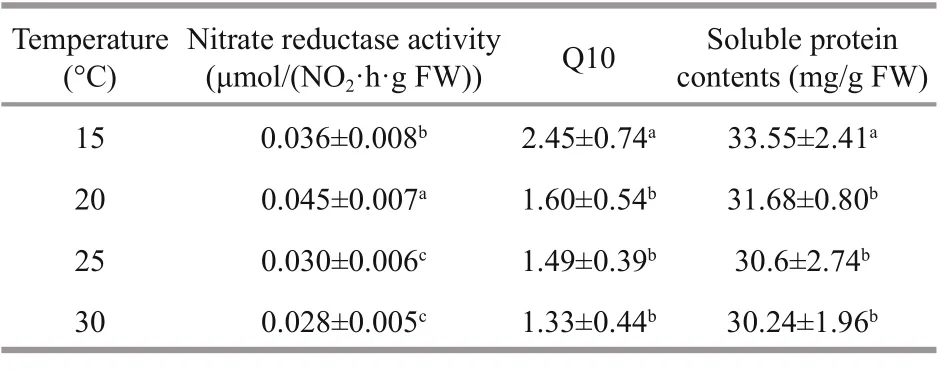
Table 1 Nitrate reductase activity, Q10, and soluble protein contents of G. lemaneiformis growing at 15, 20, 25, and 30 °C
For the nitrate reductase activity measurements, the Q10 values differed substantially with the growth temperature. The Q10 value at 15 °C was highest among all the growth temperatures. No significant ( P >0.05) was found over the range from 20 to 30 °C (Table 1).
Increasing temperatures led to lower soluble protein contents, indicating that lower temperature promoted the accumulation of soluble protein (Table 1).
3.3 Changes in PSII photosynthetic capabilities
After G. lemaneiformis had acclimated to different growth temperatures for 7 days, chlorophyll fluorescence parameters were determined by Maxiimagine PAM. Figure 3 shows the changes of the two photosynthetic properties of PSII, effective quantum yield of PSII [Y(II)] and NPQ. The highest Y(II) (about 0.36) was found at 20 °C, which was significantly greater than that at other temperatures ( P <0.05) (15 °C: 0.28; 25 °C: 0.28; 30 °C: 0.24) (Fig.3a). Contrary to the tendency of changes about Y(II) to temperature, the lowest NPQ occurred at 20 °C. Higher NPQ was found at 25 and 30 °C, which was almost twice the value at 20 °C (Fig.3b).
3.4 Rapid light curve (RLC) of G. lemaneiformis
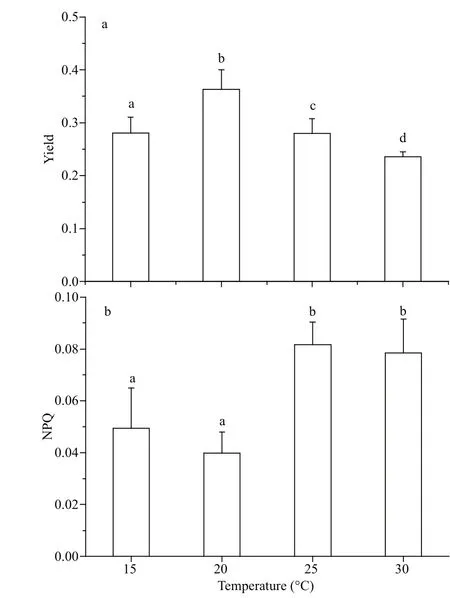
Fig.3 Changes of yield (a) and NPQ (b) of G. lemaneiformis grown at 15, 20, 25, and 30 °C, individually, under 111 μmol photons/(m2 ·s)
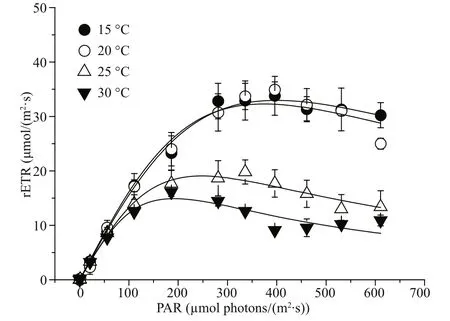
Fig.4 Rapid light curve (RLC) of G. lemaneiformis grown at 15, 20, 25, and 30 °C
The rapid light curve shows significantly different responses of photosynthetic performance to temperature ( P <0.05) (Fig.4). At 20 °C, the lightsaturated electron transport rate (ETRmax) reached its maximum value, which was significantly greater( P <0.05) than other temperatures’. The increasing temperature led to a gradual decrease in ETRmaxto a lower extent. When the growth temperature increased to 30 °C, ETRmaxdecreased to about two fifths of the value at 20 °C ( P <0.05) (Table 2). However, irrespective of the growth temperature, the electron transport eき ciency (α) showed no significant difference ( P >0.05). The saturation light ( Ik) of G. lemaneiformis cultured at 15 and 20 °C was almost equal, but was significantly greater than that at 25 °C (almost 1.7 times), and was even twice as much as the value at 30 °C ( P <0.05).
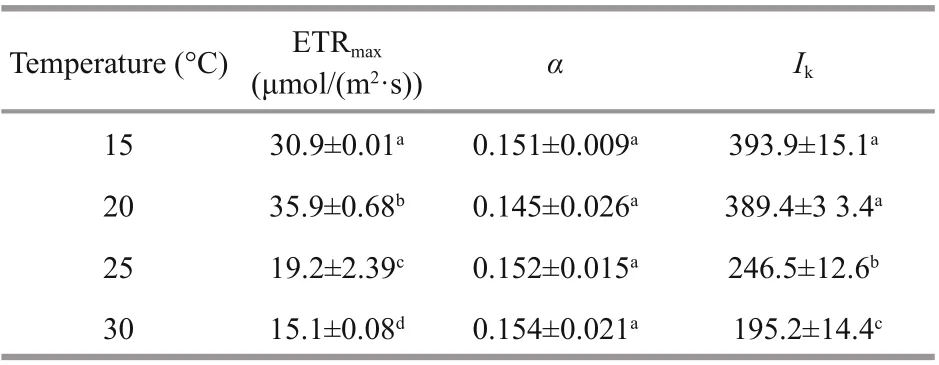
Table 2 Rapid light curve (RLC) of G. le maneiformis growing at 15, 20, 25, and 30 °C separately
The correlation between nitrate reductase activity and electron transport rate (ETR) illustrates the relationship between nitrate assimilation, photosynthetic character and physiological status. The convincing correlation observed indicated a strong relationship between nitrate assimilation and photosynthesis ( R2=0.861 16, P <0.05; Fig.5a). As shown in Fig.5b, the maximal ratio of C-fixation rate and N-incorporation rate occurred at 25 °C. At higher temperature (30 °C) the ratio tended to decrease.
4 DISCUSSION
Although the growth of G. lemaneiformis remained positive under different temperatures, and clear effects of temperature on the growth were observed (Fig.1). We found the maximum RGR occurred at 20 °C with 4.36%/d, Zou and Gao (2009) found G. lemaneiformis at 20±1 °C with RGR 6%/d, which was 37.6% more, but Xu and Gao (2009) found that RGR (2.4%/d) is less 80%. Different results may be due to the different cultural conditions.
Gracilariopsis lemaneiformis exhibited identical thermal responses for the nitrate reductase activity in the present study. To investigate the optimum temperature for the nitrate reductase activity of G. lemaneiformis in vivo assay, four differentincubation temperatures (15, 20, 25, and 30 °C) were determined, with the incubation temperature optima for the nitrate reductase activity assay in vivo should be the growth temperature in the present study. At 20 °C, the maximum nitrate reductase activity was 0.045 μmol/(NO2·h·g FW), which is 31% less than that Xu and Gao (2012) found. NH4+concentration in our natural filtrated seawater was 1.92 μmol/L, which might inhibit nitrate reductase activity partly (Chow and De Oliveira, 2008). The natural filtrated seawater we enriched with 100-μmol/L NaNO3. Nitrate is one of the most important factors regulating nitrate reductase activity (Chow and De Oliveira, 2008). Addition of nitrate can induce high nitrate reductase activity and reduce the toxic effect on the algae. On the optimum incubation temperature selected for the nitrate reductase activity assay in vivo, previous studies suggested that the nitrate reductase activity shall be determined at the optimum incubation temperature, which should be close to the maximum activity (Kristiansen, 1983; Corzo and Niell, 1991; Gao et al., 2000; Chow et al., 2004). Regardless of species and growth temperature. G. lemaneiformis cultured at different temperatures had its maximum nitrate reductase activity at 30 °C (incubation temperature), 15 °C-cultured alga had 0.135 μmol/(NO2·h·g FW) similar to the nitrate reductase activity of Ulva rigida (Corzo and Niell, 1991) and Hizikia fusiforme (Zou, 2005) when assayed in vivo at 30 °C, but lower than Gracilaria chilensis assayed in vitro, which represents a theoretical maximum for in vivo activity of the enzyme (Chow and De Oliveira, 2008). According to previous studies, 30 °C should be the optimum incubation temperature for the nitrate reductase activity assay, as shown in the Fig.2. However, the actual optimum temperature for algal growth was 20 °C (Fig.1), the same as the temperature for the maximum nitrate reductase activity of thalli determined at its growth temperature (Table 1). The optimum temperature for the maximum nitrate reductase activity did not accord with the optimum temperature for the algal growth. For Thalassiosira nordenskioeldii and Heterocapsa triquetra, the optimum temperature for nitrate reductase activity corresponded well with the optimum temperature for the growth (Jitts et al., 1964). Considering the importance of algal physiological status, we believe that the optimum temperature for the nitrate reductase activity in vivo assay is 20 °C.
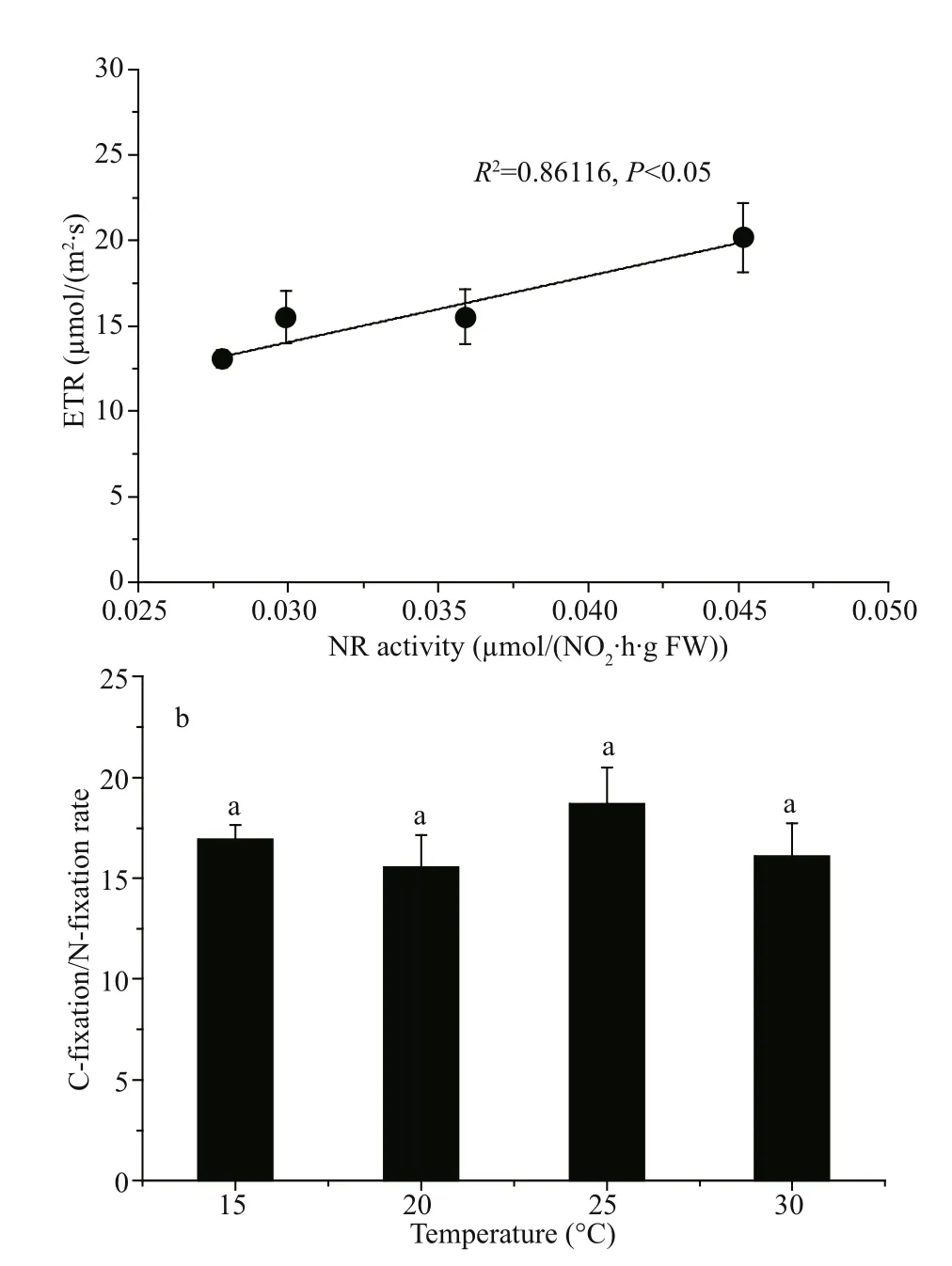
Fig.5 Correlation between nitrate reductase activity (measured at its growth temperature) and ETR (a) and the C-f ixation / N-f ixation rate varied with temperature (b)
To determine the optimum temperature for nitrate reductase activity in vivo assay, we studied the relationship between the photosynthesis and nitrate reductase activity. The assimilation of nitrate and the synthesis of enzymes, such as nitrate reductase are linked fundamentally with photosynthesis in alga (Thomas et al., 1976). After long-term acclimation (7 days), the physiological performances of alga were changed. Thalli usually have a series of mechanisms to respond the changes caused by environmental factors variation. For example, plants can optimize photosynthesis irradiance at different temperatures (Staehr and Wernberg, 2009) and increase carbon concentration mechanism expression when light intensity increases (Raven et al., 2011). Photosynthesis often displays an optimal temperature, which corresponds to the median of the non-harmful range, and decreases when the temperature increases above the thermal optimum (Sage and Kubien, 2007). At 15 and 20 °C, the cultured algae had better photosynthetic performance than those cultured at 25 and 30 °C (Fig.4). This tendency of change about ETRmaxwas the same as the nitrate reductase activity measured at its growth temperature. To promote the growth of alga, suき cient carbohydrate and protein are required. As a result, nitrate assimilation and carbon metabolism are tightly correlated (Turpin and Weger, 1988; Vanlerberghe et al., 1990; Turpin, 1991). This close connection arises from the reducing power and carbon-skeleton requirements of the synthesis of amino acids from ammonium produced during nitrate assimilation. Consequently, if either of them is affected, the other will also be affected. Meanwhile, the occurrence of maximal activities of both processes should be synchronized because it is crucial in decreasing nitrite toxicity, given that nitrite assimilation reduces nitrite to ammonium and requires reduced ferredoxin. Therefore, tight regulation between nitrate assimilation and photosynthesis has been found in many macroalgae (Chow et al., 2004; Gao et al., 2016; Xu et al., 2017), as showed in Fig.5.
Temperature is one of the most important factors controlling plant distribution and productivity (Davison, 1991; Sage and Kubien, 2007). When algae adapted to different temperatures for a long term (7 days), its photosynthetic performance showed significantly different characteristics (Figs.3 & 4). Low temperature can impair the synthesis and function of photosynthetic pigment-protein complexes and down-regulate the activities of key enzymes in the Calvin cycle to influence the photosynthesis, while the fluidity of membranes can be enhanced in high temperature, leading to a disintegration of the lipid bilayer ultimately (Nie et al., 1995; Los and Murata, 2004). Temperature at 20 °C was the optimum temperature for algal photosynthesis, at which algae had the maximum ETRmaxand Y(II), but the lowest NPQ, indicating that plants cultured at 20 °C could provide more energy for carboxylation operation, stimulated organic material synthesis for faster growth. At the same time, the maximum nitrate reductase activity occurred also at 20 °C when determined at its growth temperature, so nitrate assimilation could obtain enough needed material and power to insert the nitrogen into the carbon skeleton without producing more toxic nitrite to damage alga (Chow et al., 2004). When the growth temperature decreased to 15 °C, algal photosynthesis remained relatively unchanged (Fig.4), but the nitrate reductase activity was much lowered when determined at its growth temperature (Table 1), showing lower nitrate assimilation. As a consequence, the growth of alga was relatively lower (Fig.1). According to the results in Table 1, the lower temperature promoted high soluble protein contents, and at 15 °C cultured alga had the maximum nitrate reductase activity measured at different incubation temperatures and Q10 (15, 20, 25, and 30 °C; Table 1). This may be attributed to the fact that the contents of nitrate reductase was higher at 15 °C. However, when the growth temperature increased to 25 and 30 °C, both algal photosynthesis and nitrate reductase activity were strongly inhibited. Compared with that of algae cultured at 15 and 20 °C, high-temperature cultured algae had lower photosynthetic performance and nitrate reductase activity but higher NPQ levels, demonstrating that at 25 and 30 °C damage had already occurred in the thalli. At the end of the culturing period at 30 °C, a part of the alga had begun to decompose (data not shown), indicating that G. lemaneiformis cannot tolerate high temperature beyond 25 °C. Besides, C:N fixation rate ratio varied with temperature, and the optimum temperature was found at 25 °C. It began to decrease at 30 °C, which might be due to the inhibited photosynthesis.
5 CONCLUSION
In conclusion, the photosynthetic performance and nitrate reductase activity of G. lemaneiformis were significantly affected by temperature, and the optimum temperature for nitrate reductase activity in vivo assay was the same as growth temperature. Additionally, low temperature (15 ℃) was in favour of accumulation of nitrate reductase, but higher temperature (30 ℃) could enhance the activity of nitrate reductase. Low temperature (15 ℃) cultured G. lemaneiformis meets sudden high temperature (25 and 30 ℃) is beneficial to N assimilation.
6 DATA AVAILABILITY STATEMENT
The authors declare that all data in the present study are available upon request.
 Journal of Oceanology and Limnology2021年1期
Journal of Oceanology and Limnology2021年1期
- Journal of Oceanology and Limnology的其它文章
- Influence of sequential tropical cyclones on phytoplankton blooms in the northwestern South China Sea*
- Simulated perturbation in the sea-to-air flux of dimethylsulfide and the impact on polar climate
- Performance of ecological restoration in an impaired coral reef in the Wuzhizhou Island, Sanya, China*
- Investigating factors driving phytoplankton growth and grazing loss rates in waters around Peninsular Malaysia
- Effects of oxytetracycline dihydrate and sulfamethoxazole on Microcystis aeruginosa and Chlamydomonas microsphaera*
- Reproductive cycle of Ophiopholis mirabilis (Echinodermata: Ophiuroidea) in Zhangzi Island area, northern Yellow Sea*