
果用型三叶木通的种源选择和生物学特性
2021-02-22 08:46:48李小春胡艳波刘湘林
湖北农业科学 2021年1期
方 毅,毕 勇,李小春,胡艳波,刘湘林
(1.湖北三峡职业技术学院,湖北 宜昌 443000;2.长阳土家族自治县林业局,湖北 长阳 443500)
三叶木通[Akebia trifoliata(Thunb.)Koidz.]全身皆能利用,根、藤、果实和种子均可入药,有消炎解毒、利尿除湿镇痛及通经等功效;果味甜可食,也可酿酒;种子可榨油。近30年来,三叶木通作为新型果树研究和栽培已悄然兴起,为使栽培技术得到整体提升,特将众多学者们的研究成果从果树栽培的视角进行归纳总结。本研究从果用型三叶木通的资源选择和生物学特性方面进行总结。
1 果用型三叶木通的种源选择
1.1 果用型木通树种的选择
《中国植物志》[1]中指出,三叶木通为木通科(Lardizabalaceae)木通属(Akebia)多年生藤本植物。分布于中国的木通属植物有木通(Akebia quinata)、长序木通(Akebia longeracemosa)、三叶木通(Akebia trifoliate)3种,白木通(Akebia trifoliatasubsp.Austra⁃lis)、长萼三叶木通(Akebia trifoliatasubsp.Longise⁃pala)2亚种。木通属3个种的果实均可食用,其中三叶木通及其亚种白木通的果实较大、分布范围较广,适合于果树栽培而加以选择(表1)。本研究中的三叶木通均泛指三叶木通和白木通两亚种,特指三叶木通原亚种时加拉丁名以示区别。《中国植物志》中把种叫原亚种。

表1 木通属植物果实大小及分布情况
1.2 果用型三叶木通种质资源的选择
果用型三叶木通的品种培育还处于起步阶段,目前基本上为选取野生优株资源直接应用。生产上迫切需要从丰富的野生资源中筛选出抗逆性强、丰产性好、果实大、可食率高、品质好的优株满足当前的需求,在此基础上通过育种途径培育出优良品种。
1.2.1 果用型三叶木通地理种源的选择 王玉娟等[2]对采自赣、湘、鄂、皖4省9个地理种源的三叶木通和赣、鄂、川、渝、皖5省8个地理种源的白木通的种质资源于南昌市进行适应性比较,结果适应性最好的地理种源为:三叶木通是江西省峡江县和浮梁县2个、白木通是江西省九江市和官山自然保护区2 个。熊大胜等[3]在湘、鄂两省北纬 25°23′~30°13′(跨度4°50′)、东经 108°56′~112°57′(跨度 4°01′)、海拔240~1 400 m(跨度1 160 m)范围内收集11个点的野生三叶木通单株集中于湖南常德(北纬29°02′、东经111°40′、海拔 185 m)栽植,通过生长期茎藤生长量与月气候因子的典型相关分析得出,随海拔或纬度的升高,产生了喜温光喜湿的气候生态类型变异;随海拔或纬度的降低,产生了喜温光的气候生态类型变异。何小三等[4]采集8个不同地理种源的野生白木通果实进行分析,结果表明,安徽省宁国县军天湖乡天湖街道种源可食率(>30%)、种子含油率(47.09%)、油脂不饱和脂肪酸(72.61%)均为最高;江西省九江市东林寺东林村种源的总糖(32.35%)和还原糖(16.31%)含量最高,果肉品质最好;重庆市武隆县仙女乡白果村种源药用成分最高。
地理种源的选择是引种栽培首先考虑的因素,还应在合适的种源范围里再进一步筛选品质优、抗逆性强的优株。
1.2.2 果用型三叶木通果实品质的选择 优质的果实品质是果用型三叶木通栽培的首要任务,下面就果实品质的主要方面进行讨论。
1)果实大小。人工栽培条件下三叶木通果实的大小,钟彩虹等[5]测定平均单果重≥200 g、最大单果重 546 g;罗克明等[6]测定最大果重 313.5 g、最小97.7 g、平均78.1~244.1 g;万明长等[7]测定最大单果重 313.5 g、最小 89.5 g、平均 158.5 g;仲伟敏等[8]测定优株最大单果重252.00 g,平均果重136.52 g。从以上研究结果来看,选择个大的优株栽培,平均单果重达250 g是可以实现的。
2)可食率。可食率是评价果实品质的重要指标之一。三叶木通的果实可食率较低,钟彩虹等[5]认为人工栽培条件下果皮变薄、果肉增厚、种子数减少、可食率提高,平均可食率为30.7%,最高可食率达 50.4%;万 明 长 等[7]测 定 可 食 率 为 22.81%~34.39%;仲伟敏等[8]测定三叶木通优株可食率为30.44%;李金光等[9]测定野生果实中果皮、种子、果肉分别占69.1%、6.2%、24.7%。三叶木通果实可食率(一般仅为30%)严重偏低且种子多,极大地影响了果实的品质,提高果实的可食率和实现无子化是今后品种选育应突破的难点之一。

表2 三叶木通果实主要营养成分含量
3)果肉营养。三叶木通果实营养丰富,富含氨基酸、蛋白质、脂肪、糖类、矿物质和多种维生素。万明长等[7]、仲伟敏等[8]、李金光等[9]、刘伦沛等[10]对三叶木通,王德智等[11]、何小三等[4]对白木通果肉测定的主要营养成分见表2。从表2可以看出,尽管各位学者的测定结果不尽一致,但三叶木通果实营养成分丰富,优于一般水果。
1.2.3 果用型三叶木通丰产性和抗逆性的选择 果用型三叶木通栽培历史短,大面积大规模栽培还未兴起,对病虫害、抗逆性、丰产性的研究极为少见,培育质优、丰产、高抗的品种是实现高效栽培不可回避的课题,需要科技工作者们长期不懈地努力探索。
2 果用型三叶木通的生物学特性
生物学特性是栽培的基础,根据学者们研究的成果将从物候期、茎藤生长、花芽分化、开花和果实发育4个方面进行归纳。
2.1 三叶木通的物候期
系统进行三叶木通物候期观察的记载较少,根据庞发虎等[12]萌芽期3月中旬至下旬、现蕾期4月上旬、开花期4月下旬,果熟期8月中旬至9月上旬的观察报道,结合熊大胜等[13]对三叶木通各物候期相对时间的报道,绘制物候期示意图(图1)。图1中落叶期为笔者在湖北宜昌的观察结果,与史敏华[14]所观测的落叶期10月20至11月20的结果基本一致。
从图1中可以看到,三叶木通的生长期为260 d左右;开花期、果实生长期与新梢生长重叠,会造成落花落果和抑制果实生长;7—10月是花芽分化的高峰期,与新梢生长和果实发育同步,需要通过栽培措施协调三者之间的矛盾。
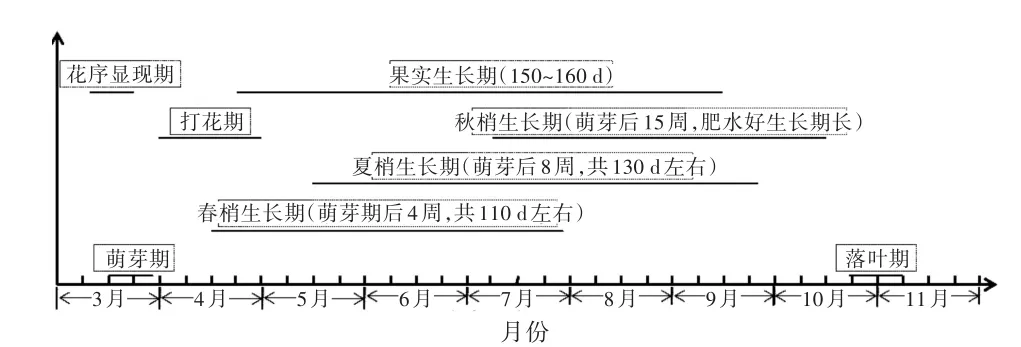
图1 三叶木通物候期示意图
班小重等[15]在海拔1 140 m的贵州省园艺研究所苗圃对三叶木通10个优株物候进行观测,结果表明,萌芽期为2月下旬,开花期为2月下旬至3月中旬,落叶期为2月中旬至3月中旬,展叶期为3月上旬,果实膨大期为5月上旬至8月下旬,果实着色期为6月上中旬,果实成熟期为8月下旬至9月中旬;文中所描述的落叶期为“老叶脱落和开花、新叶生长同步”的结论值得商榷,落叶期不应推迟到2月中旬至3月中旬。主要对班小重等[15]的落叶期有怀疑,一般落叶树为冬季落叶,常绿树种春季发新叶后再落叶,三叶木通为落叶树种。
2.2 三叶木通的茎藤生长
熊大胜等[13,16-18]对三叶木通茎藤生长有比较系统的研究,认为年抽梢3次,春梢生长期110 d,夏梢生长期130 d,秋梢在水肥条件充足下生长期较长、不能分化出花芽、多冬寒枯死;影响茎藤生长快慢的气候因子依次为温度、光照、降雨,高湿少光利于茎藤生长、低湿足光利于茎藤萌梢;钾肥对茎藤生长有显著或极显著的促进作用,5 mg/L生长调节剂(GA3、NAA、KT)对已经抽出的茎藤有促进生长作用,浓度高达20 mg/L时有促进萌梢的作用。
2.3 三叶木通的花芽分化
李金光等[19]对三叶木通花芽形态分化进行了研究,认为三叶木通的花芽为混合芽,着生于当年生新梢叶腋或短缩枝顶端,顶花芽占花芽总数的比率较腋花芽高、座果也较为可靠;顶花芽于7月初在停止生长一个月左右的短缩枝上开始分化,8月初和9月初两个分化高峰;腋花分化比顶花芽分化晚25 d左右、9月下旬至10月中旬为分化高峰期,主要在新梢第2~10节分化,第2~25节也能分化,这与熊大胜等[13,20]的研究结论一致;雌花分化早于雄花,进入冬季休眠前雄花分化出雄蕊原基(花药)、雌花分化出心皮结构;花芽分化可分为未分化期、花蕾分化期、花被分化期、雄蕊分化期和雌蕊分化期5个时期。
熊大胜等[21]采用L8(37)正交设计,研究了栽培技术对三叶木通花芽分化量的影响:全土(不掺砂或煤渣)且有机质含量高、花芽分化始期和盛期追施N∶P∶K为1∶1∶3的肥料、3—9月喷施21次(3次/月)5 mg/L的萘乙酸(NAA),花序和雌花芽分化量大。
2.4 三叶木通的开花和果实发育
2.4.1 三叶木通的开花 熊大胜等[13,20]、李嘉瑞等[22]研究了三叶木通的开花生物学,熊大胜等[13,20]认为开花期为30 d,李嘉瑞等[22]认为是45 d,这与所取材料花芽分化期长短有关,通过栽培调控使花芽集中分化会极大地缩短花期和果熟期,便于生产管理;花期与新梢生长同时,营养生长与生殖生长重叠,是造成落花落果率很高的重要原因;完全花序占82.2%~90.0%,雄花序占9.0%~14.9%,单生雌花占1.0%~1.2%,完全花序在产量构成中占有极其重要的位置;雌花开花高峰比雄花早4~6 d。
张希凤等[23]认为三叶木通雄花散粉当天花粉活力最高,达91.96%,随时间延长花粉的活力逐渐降低,室温下3 d和4 d后花粉活力分别下降至62.99%和11.72%,3~4 d是雄花维持寿命的时间;雌花开放当天柱头具有较弱可授性但无黏液分泌,花后2~3 d可授性增强且黏液分泌旺盛,花后3~5 d为可授性高峰期。三叶木通的雌花比雄花先开4~6 d,基本上能保证雌雄花同熟,不需另外配置授粉品种。
2.4.2 三叶木通的果实发育 钟彩虹等[5]、李金光等[9]、熊大胜等[20]对三叶木通果实纵、横生长呈双“S”曲线的结论完全相同;在划分果实生长期上有差异:钟彩虹划分为快速生长期、缓慢生长期、相对稳定期、熟前膨大期4个发育时期;李金光等[9]划分为速生期、缓长期、停长期、熟前生长期4个时期,与钟彩虹等[5]划分基本相同;熊大胜等[20]划分为受精结实期、速生期、缓生期、充实期、熟前生长期5个时期,后4个时期与钟彩虹等[5]和李金光等[9]的划分基本相同,钟彩虹等[5]、李金光等[9]未包括受精结实期在内。各时期的持续时间和生长情况见表3。
从表3可以看出,果实发育时间差异较大,钟彩虹等[5]的为 170 d、李金光等[9]的为 114 d、熊大胜等[20]的为147 d,主要原因可能是各自划分生长时期的起点和标准不同所致,其次是各自所取材料和所处环境差异的原因;李金光等[9]的114 d再加上受精结实期28 d为142 d,与熊大胜等[20]的147 d基本吻合;从生长速度来看,钟彩虹等[5]的前两个时期可能包含熊大胜等[20]的前3个时期。栽培上提高单果重的重要措施是抓住开花后的40 d快速生长期,其次为30 d的缓慢生长期;熟前膨大期30 d的管理可增加单果重,对品质的提高也至关重要。

表3 三叶木通果实发育时期、持续时间及生长量
3 小结
果用型三叶木通应选择三叶木通及亚种白木通的优良种源,再从单果重、可食率、营养成分、丰产性、抗逆性等方面筛选优良单株,可食率大幅度提高和实现无子化是育种工作需要突破的重点。三叶木通的生长期为260 d左右,影响茎藤生长的气候因子依次为温度、光照和降雨,钾肥对茎藤生长影响显著;新梢生长与开花坐果、果实发育、花芽分化重叠,导致营养生长与生殖生长的矛盾,需要采用合适的栽培手段进行调节。
三叶木通于8月初和9月初在短缩枝顶端两次出现花芽分化高峰;腋花芽仅9月下旬至10月中旬一个分化高峰,花芽主要集中在春、夏梢的第2~10节;提高土壤有机质含量和花芽分化初期增施以钾为主的肥料等栽培技术能促进花芽分化。三叶木通雌花开放早于雄花4~6 d,但雌花开放后3~5 d方为可授粉高峰期,使雌雄基本同熟,不需另配授粉品种;长达30~45 d的开花期和30 d以上的果实采收期,非常不利于集约栽培,需要通过栽培措施使花芽分化期从3个月缩短至20 d以内甚至更短,方能得到彻底解决。
三叶木通果实生长期长达150~170 d,花后40 d的快速生长期和其后30 d的缓慢生长期的管理措施对提高单果重至关重要,30 d的熟前膨大期对单果重和品质有着双重影响。
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
辽宁林业科技(2021年1期)2021-03-16 09:17:08
生态学报(2019年15期)2019-09-04 09:30:22
水产科学(2019年1期)2019-01-17 02:05:26
基层中医药(2018年2期)2018-05-31 08:45:08
广西林业科学(2016年3期)2016-03-16 05:43:29
大连海洋大学学报(2016年6期)2016-02-10 08:10:26
中国民族民间医药·下半月(2014年5期)2014-12-02 05:15:58
食品科学(2013年22期)2013-03-11 18:29:16
