
1 株野生平菇菌株的鉴定及交配型分析
2021-02-16 08:33刘晓雪王春霞刘利娟郭金英郑素月
河南农业科学 2021年12期
刘晓雪,王春霞,刘利娟,郭金英,郑素月
(河北工程大学 园林与生态工程学院,河北 邯郸056038)
平菇(Pleurotus ostreatus)食味鲜美,营养丰富,具有极高的食药用价值,特别是其富含的黄酮类、有机酸、几丁质等多种活性成分可以有效抑制癌细胞生长,增强人体免疫力[1-2]。近年来随着人们对饮食健康越来越重视,对平菇的需求与日剧增。同时,作为适应性较强的侧耳属木腐真菌[3],平菇的栽培范围遍及全球各大洲,我国是平菇栽培大国,据中国食用菌协会统计,2019 年我国平菇产量686.47万t,为我国栽培量仅次于香菇和黑木耳的第三大食用菌品种[4]。
目前我国的食用菌基本实现人工种植,其栽培技术已非常成熟,但是在实际生产过程中经常因为菌株交配型无法准确鉴定,而导致产生亲和率下降、产量不稳定、经济效益下滑等问题[5-7],给食用菌产业带来巨大经济损失,因此菌株交配型的鉴定尤为重要[8]。平菇是担子菌中标准的四极性异宗结合菌[9],其交配型系统研究最早开始于20 世纪20 年代[10],研究者对交配型做了大量工作。季哲等[11]以2个黄伞子实体为材料,用孢子稀释法分离孢子,经3轮杂交后,鉴定出黄伞为四极性异宗配合的担子菌。余梅等[12]采用原生质体单核化和单孢分离的方法发现,真姬菇为四极性异宗结合担子菌。杨军等[13]通过核迁移试验鉴定出灰树花属于四极性交配系统。冯伟林等[14]鉴定出杏鲍菇属于四极性异宗配合系统,并且应用ISSR 分子标记技术对获取的4 种不同交配型的单核体进行DNA 指纹分析,认为ISSR 分子标记技术可以作为食用菌单核菌株鉴别及指纹分析的有效工具。柯斌榕等[15]以赤芝JC 和AL 菌株为试验对象,在常规交配型测定的基础上,采用OWE-SOJ 技术对其单核菌株交配类型进行测定,确定了赤芝为四极性异宗结合菌。程莉等[16]采用标准测交法鉴定糙皮侧耳的交配型系统为四极性异宗配合真菌。单核菌株作为重要的育种材料[17-18],对交配系统的分析至关重要。交配系统以及交配行为的研究不仅仅是杂交育种工作的基础,也是从交配型因子的角度理解和评价菌株资源遗传多样性的基础。交配型涉及亲和杂交、双核菌丝体构建和子实体发育等重要生理过程,是值得深入研究的一个课题[19]。对侧耳属菌株的研究虽然已经获得了部分菌株的基因组序列,但是食用菌交配型位点和基因的研究仍旧存在很多空白,还没有从全基因组的水平对这些物种的交配型位点和交配型基因进行深入研究[20]。并且目前关于野生侧耳属菌种的鉴定及交配型分析更是鲜见报道。为此,在ITS鉴定的基础上,通过单孢分离法对1株野生侧耳属平菇菌株进行孢子单核菌株分离,经过3 轮杂交和核迁移试验,对该平菇菌株的4 种交配型进行鉴定,为今后从交配型因子的角度深入研究侧耳属平菇的遗传规律及扩充种质资源库等工作奠定基础。
1 材料和方法
1.1 试验材料
野生平菇子实体采集于河北工程大学原中华南校区一棵龙爪槐树干上,由河北工程大学食用菌研究室分离、鉴定和保藏,菌株编号为Po15。
1.2 方法
1.2.1 ITS 鉴定 采用DNA 试剂盒(天根生化科技北京有限公司)提取菌丝体DNA,利用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATTGATATGC-3′)进行扩增。PCR 产物送上海生工生物工程有限公司进行克隆测序。将得到的菌株ITS 序列在NCBI 数据库中进行BLAST 比对,利用ClustalX 软件对所得序列进行校正和比对分析,采用MEGA 6.0 软件以Neighbor-joining法构建系统进化树。
1.2.2 单孢分离方法 采集平菇子实体进行多孢分离和单孢分离,具体方法参考文献[21]。
1.2.3 交配型测定方法 单核菌株交配型常规测定参照张树庭等[22]方法,测定程序如表1所示。

表1 四极性交配型测定程序Tab.1 Procedure for determination of tetrapolar mating type
1.2.4 核迁移试验 参考程莉等[16]、李安政等[23]方法,采用核迁移试验对该平菇的交配型进行鉴定。

2 结果与分析
2.1 ITS鉴定和分析
利用ITS1 和ITS4 通用引物对野生菌株进行扩增、测序后,将测序结果进行BLAST 比对,发现目标序列与登录号为LC149608.1 的平菇菌株同源性达到99.71%。并通过构建的系统发育树分析(图1),该野生菌种与Pleurotus pulmonarius(肺形侧耳)、Pleurotus floridanus(佛州侧耳)具有一定的遗传距离,而与Pleurotus ostreatus(MT778826、MT778816、MT778817)3 个平菇菌种相似性达到100%。因此,结合形态学可确定分离菌株为糙皮侧耳。
2.2 单核菌株群体的获得
根据单孢分离法,挑取了100个菌株,经显微观察和生长速度测定,剔除有锁状联合及长速较差菌株,最终获得69株可用的孢子单核菌株。对其生长速度、菌落形态等生物学指标进行测定,结果表明,69 株单核菌株的菌落形态主要分为4 类(图2),与程爽爽等[25]在香菇单核菌丝研究中的结果一致。4种类型包含的菌株见表2。

表2 糙皮侧耳单核菌株的菌落形态分类结果Tab.2 Classification results of monokaryon strains from Pleurotus ostreatus by colony types
2.3 交配型鉴定结果
根据四级性交配型测定程序将获取的69 株单核菌株进行交配型鉴定。首先将P1 菌株假设为T1(AxBx)进行68组配对试验,与T1亲和的有37株,其交配型为AyBy;第2 次试验将从第1 轮亲和菌株中挑选P8 菌株为T2(AyBy),进行31 组配对试验,与T2亲和的有19 株,其交配型为AxBx;第3 次试验从剩余的既不与T1亲和也不与T2亲和的菌株中挑选P10菌株为T3(AxBy),进行11 组配对试验,与T3亲和的有3 株,其交配型为AyBx,不亲和的有8 株,其交配型为AxBy。以此3 轮交配共进行110 组配对试验,将69 株单核菌株分成4 类,结果如表3 所示。在进行配对试验过程中,需将单核菌株中生长速率居中的菌株作为测交菌株T1、T2和T3,以防止试验过程中出现一方长得过快或过慢而影响配对试验结果。
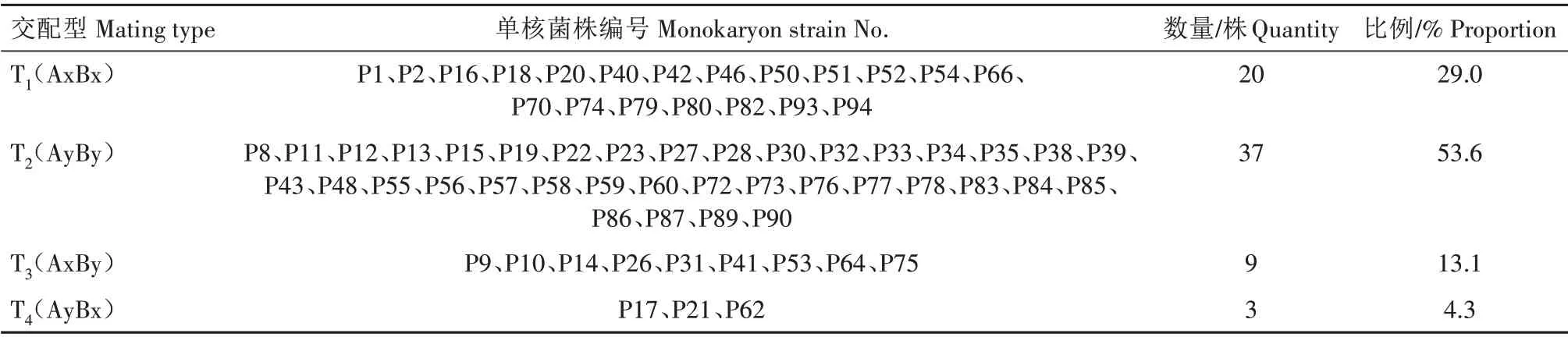
表3 糙皮侧耳单核菌株3轮配对结果统计Tab.3 Statistical results of three rounds of pairing of monokaryon strains from Pleurotus ostreatus
2.4 核迁移试验鉴定结果
根据3 轮交配结果,从T1(AxBx)、T2(AyBy)、T3(AxBy)、T4(AyBx)4 个类型中分别抽取生长速度一致的单核菌株P16、P8、P14、P17 做核迁移试验,流程如图3所示。
由于A 因子控制的是菌丝的融合,B 因子控制核的迁移,所以在一种非标准型四极性的不亲和反应中,想知道哪一种是A=B≠,哪一种是A≠B=,哪一种是A=B=,就必须进行核迁移等补充试验。结果表明,因P8×P16 为可亲和菌株,所以P8 与P16 为A≠B≠,则P8(T2)的交配型为A2B2;P8×P14 未发生细胞核的迁移,所以P8 与P14 为A≠B=,则P14(T3)的交配型为A1B2;而P8×P17 发生了细胞核的迁移,所以P8 与P17 为A=B≠,则P17(T4)的交配型为A2B1。野生菌株孢子单核体的4 种交配型得到鉴定,如表4所示,说明该野生平菇为四极性异宗结合真菌。
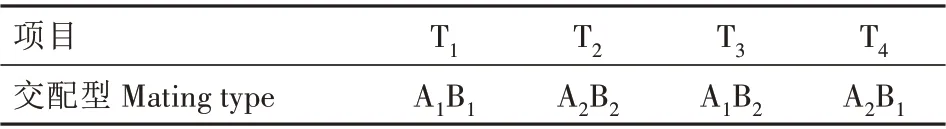
表4 糙皮侧耳单核菌株交配型测定结果Tab.4 Determination of mating type of monokaryons strains of Pleurotus ostreatus
2.5 交配型比例χ2检验结果
交配型鉴定结果显示,采收的野生平菇经单孢分离后,其交配型出现了偏分离现象。在获取的69株单核菌株中,T1、T2、T3、T44 种交配型的比例为20∶37∶9∶3,通过计算得知χ2=39.35。检测结果显示,实际测得的交配型比例与预期的1∶1∶1∶1 的比例不相符。
3 结论与讨论
交配型的鉴定一般采用的是单孢分离或原生质体单核化等方式获得单核菌株,再根据标准测交法及核迁移试验对菌种进行鉴定[11-16]。本试验在ITS 鉴定的基础上,通过以上方法对野生平菇的69株单核菌株进行交配型鉴定,结果表明,该野生平菇为四极性异宗结合的糙皮侧耳。
在69 株单核菌株的培养过程中发现其菌落形态大致分成了4 种类型,该结果与对香菇[25]、灵芝[26]等研究结果一致。造成菌丝形态、菌落特征的差异可能与其交配的A、B 控制因子有关,因为在交配型鉴定的常规程序中,同样也会根据菌落交接处的菌落形态来确定其交配类型。但是本试验中4种交配类型与菌落形态分类并不一致,因此是否可以根据菌落的形态来初步确定其交配型,或者说菌落类型与交配型之间是否有一定的关系还有待研究。
本试验中获得的T1、T2、T3、T44种交配型的比例与孟德尔的分离规律1∶1∶1∶1 不符,实际比例出现了偏分离现象。林芳灿等[24]对香菇研究发现,原生质体单核体的交配型比例同样出现严重的偏分离,且2种交配型的比例相差可达43∶1。朱朝辉等[27]以香菇为例研究了交配型与单核体再生能力之间的关系,结果显示,不同交配型单核体的再生能力是有差异的,造成比例出现偏差可能与某种交配型的单核体再生能力弱有关。程水明[28]同样以香菇为试材,发现来自同一双核菌株F1担孢子的菌丝萌发能力和生长速度是有差异的,这种差异导致了4种交配型分布不均衡,从而造成偏分离现象的发生。本研究中,来自同一野生平菇菌株的69株孢子单核体在菌丝培养阶段,同样出现了菌落形态、菌丝生长速度及萌发能力的差异,而这些现象均有可能导致偏分离现象的发生。
猜你喜欢
农业知识(2022年9期)2022-10-13
新疆农业科学(2022年5期)2022-06-15
中学生报·教育教学研究(2022年1期)2022-04-18
电脑报(2021年8期)2021-07-01
电脑爱好者(2020年24期)2020-12-22
小学生(看图说画)(2020年10期)2020-10-29
医学信息(2019年19期)2019-11-15
时代英语·高一(2019年5期)2019-09-03
农产品市场周刊(2014年15期)2014-08-22
数理化学习·高一二版(2009年2期)2009-03-30
