
四川山胡椒叶绿体基因组特征及山胡椒属系统发育*
2021-02-12 05:58唐利洲韩利红
林业科学 2021年12期
刘 潮 唐利洲 韩利红
(曲靖师范学院生物资源与食品工程学院 云南省高校云贵高原动植物多样性及生态适应性进化重点实验室 云南省高校特色果酒技术创新与应用工程研究中心 曲靖 655011)
四川山胡椒(Linderasetchuenensis)为樟科(Lauraceae)山胡椒属(Lindera)常绿灌木,多生于山坡路旁及疏林中,分布于我国四川、贵州等地(http:∥www.iplant.cn/frps)。四川山胡椒叶片提取物香精油对金黄色葡萄球菌(Staphylococcusaureus)、白色念珠菌(Candidaalbicans)和大肠杆菌(Escherichiacoli)均具有明显的抑制作用(Weietal., 2016),其叶片中提取的多种化合物具有抗肿瘤活性(袁娟娟, 2018; 张嘉穗, 2016)。
叶绿体是质体的一种,为绿色植物光合作用的场所,具有相对独立的遗传系统。陆生植物叶绿体基因组大小范围为107~218 kb,具有高度保守的四分体结构,包括2个反向重复区(inverted repeats,IR),以及被IR分隔的大单拷贝区(large single copy,LSC)和小单拷贝区(small single copy,SSC)(Danielletal., 2016)。叶绿体基因组编码120~130个基因,主要参与光合作用、基因转录和翻译(Danielletal., 2016)。目前多个山胡椒属物种叶绿体基因组已被相继报道(Songetal., 2020; Zhaoetal., 2018; Tianetal., 2019)。完整的叶绿体基因组序列对于了解植物相近类群之间的系统发育关系和物种进化具有重要价值(Danielletal., 2016),基于高通量测序的系统进化基因组学研究,成为全面理解生物进化的重要工具(Songetal., 2020)。叶绿体基因组数据已经很好地解决了樟科等多种植物类群的系统发育关系(Songetal., 2017; 2020; Zhaoetal., 2020; Xieetal., 2020)。密码子使用偏性可以反映叶绿体基因组进化特征及其影响因素,在基因表达中起重要作用(Boeletal., 2016)。叶绿体基因组条形码可用于品种鉴定和种质资源保护,物种特有的重复序列可以作为一种潜在的遗传多样性标记。叶绿体基因组结构特征和进化研究,对于分析物种遗传背景和系统发育关系都具有重要的意义。
四川山胡椒具有重要的潜在药用价值,然而关于四川山胡椒物种遗传多样性、进化和发育方面的研究较少,未见叶绿体基因组方面的报道。本研究首次通过高通量测序技术获得四川山胡椒叶绿体基因组序列,并对叶绿体基因组序列重复、密码子使用偏性,山胡椒属物种叶绿体基因组高变区和进化发育关系进行分析,以期为四川山胡椒的遗传背景、进化发育和种质资源保护提供借鉴。
1材料与方法
1.1 试验材料 四川山胡椒采自江苏省南京中山植物园(118°50′3.19″E,32°03′9.67″N),新鲜叶片置于硅胶中保存。标本存放于中国科学院西双版纳热带植物园(标本号XTBG-BRG-SY36159)。用改良的CTAB法从叶组织中提取四川山胡椒总DNA(Doyleetal., 1987),构建测序文库。基于Illumina Genome Analyzer Hiseq 2000测序平台进行叶绿体基因组测序。
1.2 四川山胡椒叶绿体基因组组装与注释 使用GetOrganelle软件(Jinetal., 2020)组装获得完整叶绿体基因组。四川山胡椒完整叶绿体基因组数据上传LCGDB 数据库(http:∥lcgdb.wordpress.com),访问号LAU00108。以山胡椒(Linderaglauca)叶绿体基因组(登录号MG581443)为参考,利用GeSeq(https:∥chlorobox.mpimp-golm.mpg.de/geseq.html)进行基因组注释。利用OGDRAW软件(https:∥chlorobox.mpimp-golm.mpg.de/OGDraw.html)绘制叶绿体基因组图谱(Greineretal., 2019)。
1.3 四川山胡椒叶绿体密码子使用偏性分析 利用CodonW 1.4.2软件分析四川山胡椒叶绿体基因组编码基因的有效密码子数(effective number of codon,ENC)以及鸟嘌呤(guanine)和胞嘧啶(cytosine)含量(GC),通过EMBOSS在线网站(http:∥emboss.toulouse.inra.fr/)的cusp软件分析相对同义密码子使用度(relative synonymous codon usage,RSCU)。分别选取ENC值较小和较大的10%基因作为高表达基因(high expression gene,HEG)和低表达基因(low expression gene,LEG),计算ΔRSCU值(ΔRSCU=RSCU高表达-RSCU低表达)。密码子RSCU>1则为高频密码子,同时满足ΔRSCU≥0.08则为最优密码子。
1.4 四川山胡椒叶绿体序列重复分析 通过REPuter软件(Kurtzetal., 2001)分析序列重复,最小重复长度为30 bp,序列同源性90%,Hamming距离3,分析了互补(complementary)、正向(forward)、反向(reverse)和回文(palindromic)重复。利用MISA在线工具检测SSR(Beieretal., 2017),分析的最小阈值为单核苷酸重复次数8,二核苷酸重复次数4,三核苷酸重复次数4,四核苷酸、五核苷酸和六核苷酸重复次数均为3。
1.5 山胡椒属核苷酸多样性分析 通过MAFFT v. 7.450(Katohetal., 2019)对山胡椒属物种叶绿体基因组序列进行多序列比对,利用DnaSP 6.0计算核苷酸多态性值(Rozasetal., 2017),设置搜索窗口长度为600 bp,步长为200 bp,使用R程序绘图。
1.6 山胡椒属系统发育分析 从NCBI和LCGDB数据库下载30种山胡椒属和樟属(Cinnamomum)物种的叶绿体基因组序列,其中三桠乌药(Linderaobtusiloba,LAU00057)序列由LCGDB数据库下载,其他物种序列由NCBI的GenBank数据库下载。利用四川山胡椒和28个山胡椒属物种的叶绿体基因组序列,以樟(Cinnamomumcamphora)和沉水樟(C.micranthum)为外类群,使用MAFFT v. 7.450进行多序列比对,通过IQ-TREE v. 2.1.1软件(Minhetal., 2020)基于最大似然法(maximum likelihood,ML)构建系统进化树,建树模型为K3Pu+F+R3,步长值为1 000。
2 结果与分析
2.1 叶绿体基因组结构特征 四川山胡椒叶绿体基因组全长为152 851 bp(图1),与其他山胡椒属物种大小相当(Tianetal., 2019; Songetal., 2020; Zhaoetal., 2018),呈典型的四分结构,LSC、SSC和IR区大小分别为93 742 bp、18 977 bp和20 066 bp。基因组总GC含量为39.15%,IR区GC含量最高(44.42%),其次为LSC(37.94%),SSC最低(33.95%)。四川山胡椒叶绿体基因组共编码125个基因,其中蛋白编码基因81个,转运RNA(tRNA)36个,核糖体RNA(rRNA)8个,13个基因为双拷贝基因。
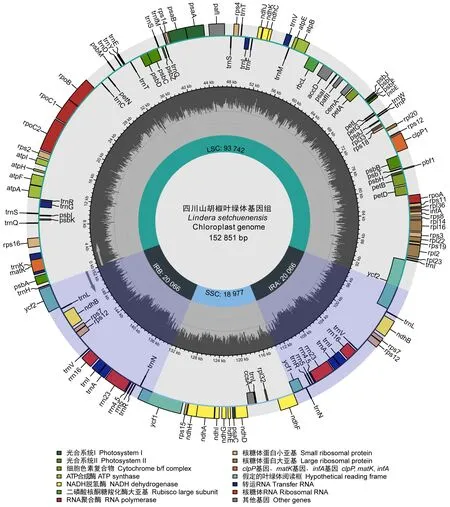
图1 四川山胡椒叶绿体基因组结构Fig. 1 Complete chloroplast genome map of Lindera setchuenensis
2.2 叶绿体蛋白编码基因密码子偏性分析 四川山胡椒叶绿体蛋白编码基因GC含量为39.13%,鸟嘌呤和胞嘧啶碱基的出现频率GC3s为27.96%,平均ENC值为49.11。蛋白编码基因高频密码子有31个(表1),其中13个以A结尾,16个以U(T)结尾,以G和C结尾的分别有1个。最优密码子有21个,其中9个以A结尾,11个以U(T)结尾,1个以G结尾。
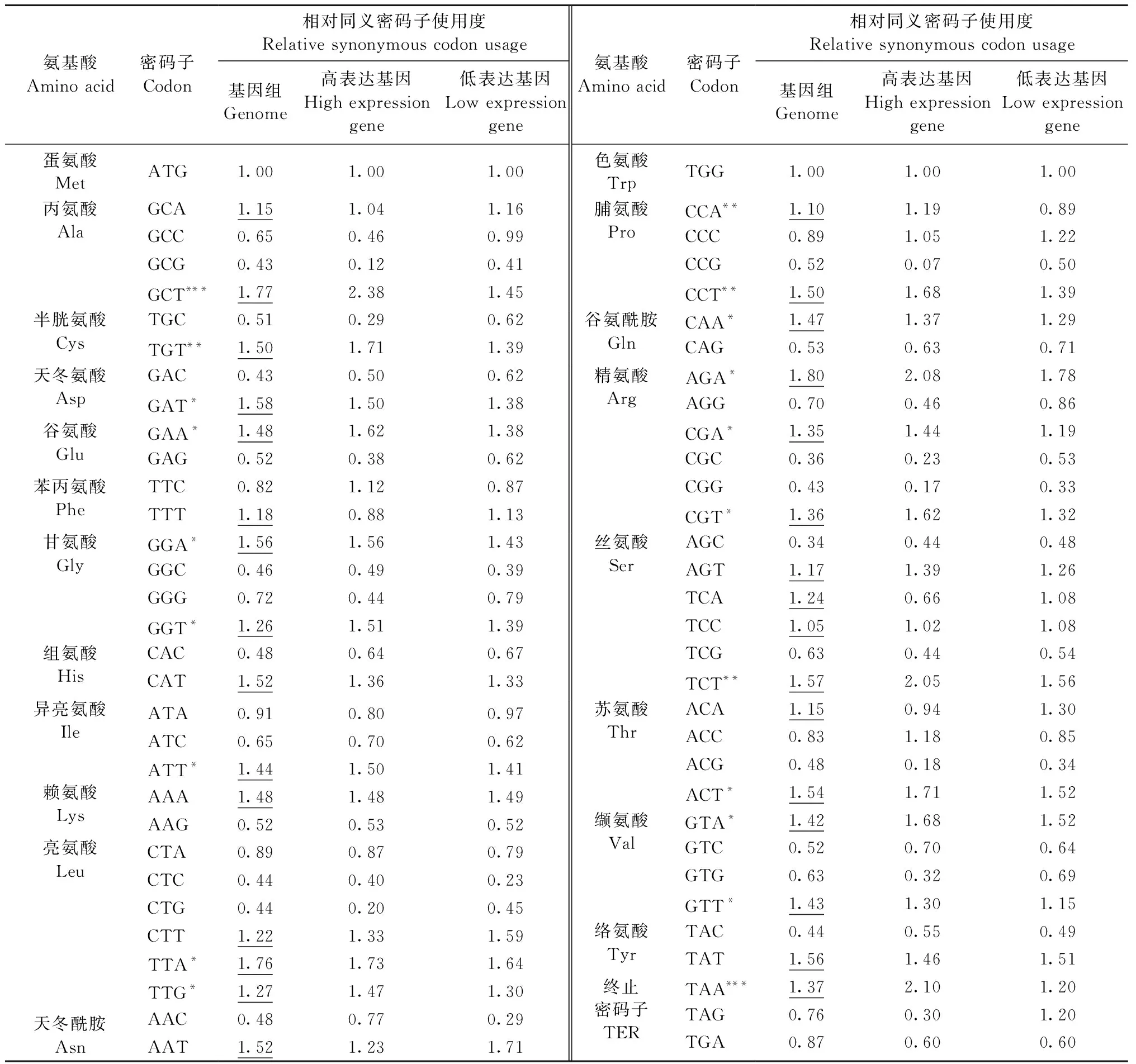
表1 四川山胡椒叶绿体基因组相对同义密码子使用度①Tab.1 Relative synonymous codon usage of genes in the chloroplast genome of Lindera setchuenensis
2.3 叶绿体基因组序列重复分析 四川山胡椒叶绿体基因组中共检测到27对长序列重复(图2),长度30 bp的序列数目最多,位于IR、LSC和SSC上的数目分别为8、13和3,其次为33 bp的序列,分别有14和2个位于IR和LSC上。长序列重复中,正向重复最多,达24个(占44%),其次为回文重复22个(占41%),反向重复和互补重复分别为6个和2个,其中33 bp和30 bp的序列在正向重复和回文重复中均占较高比例,30 bp重复多数位于LSC和IR上,33 bp重复多数位于IR上。
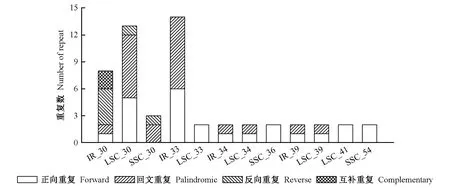
图2 四川山胡椒叶绿体基因组序列重复Fig. 2 Repeats in the chloroplast genome of Lindera setchuenensisLSC、SSC和IR分别表示大单拷贝区、小单拷贝区和反向重复区,其后数值表示序列重复长度。LSC, SSC and IR represent large single copy, small single copy and inverted repeats, respectively, and the subsequent values represent the sequence repeat length.
四川山胡椒叶绿体基因组中发现13种类型共181个SSR位点(图3)。其中,单碱基SSR数目最多,占SSR总数的71.27%,其次为双碱基SSR,占20.44%。A/T类SSR占单碱基SSR的95.35%,AT/AT类碱基占双碱基SSR的51.35%(图3)。55%的SSR位于基因间区,29%位于编码序列(coding sequence,CDS)上,15%位于内含子区,仅有1%位于rRNA上(图4)。
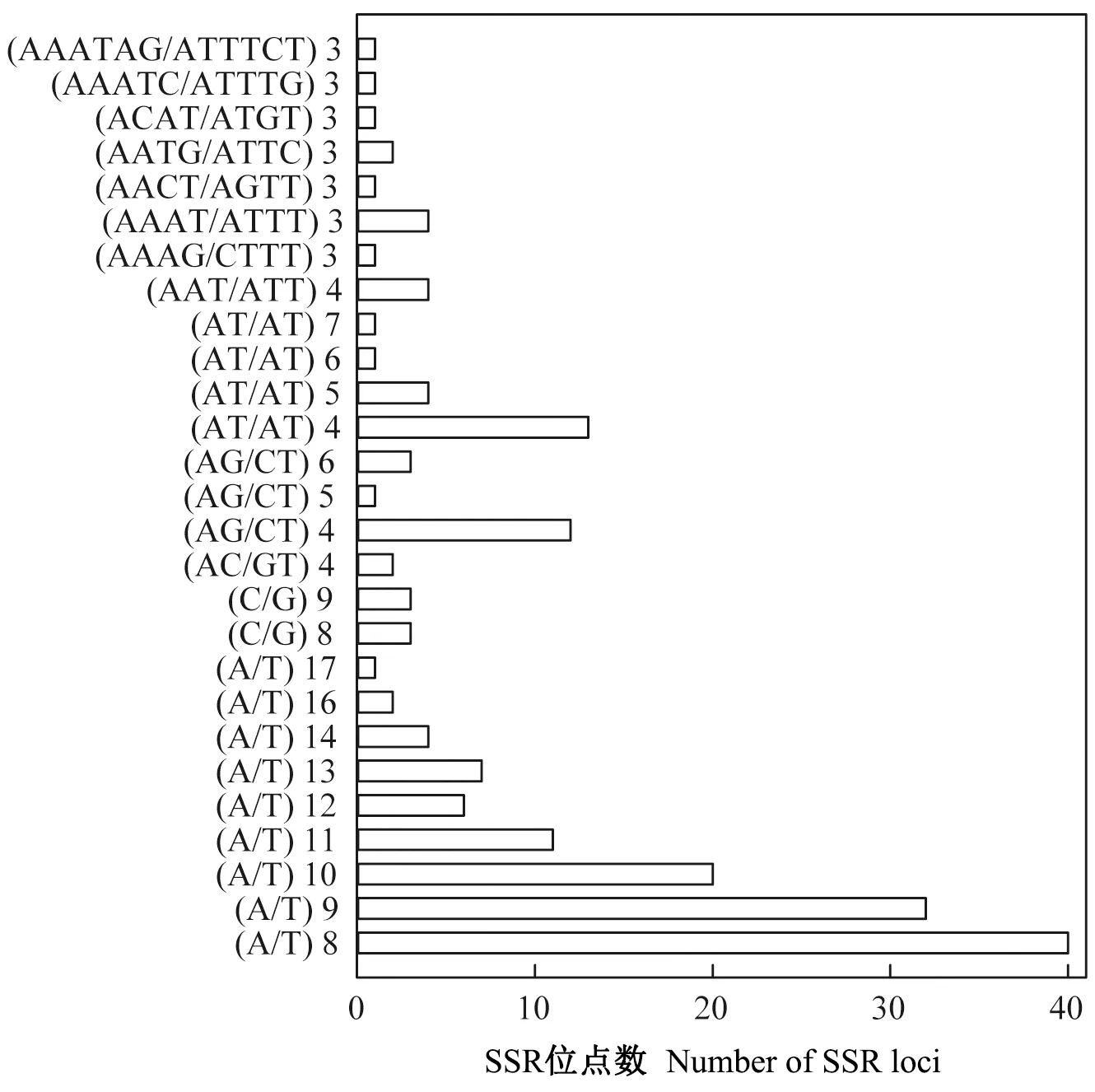
图3 四川山胡椒叶绿体基因组SSR位点类型及数量Fig. 3 The types and number of SSRs in the chloroplast genome of Lindera setchuenensis
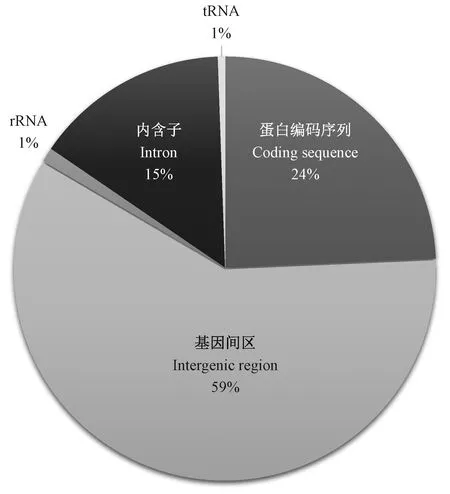
图4 四川山胡椒叶绿体基因组SSR 位点分布Fig. 4 The distribution of SSRs in the chloroplast genome of Lindera setchuenensis
2.4 山胡椒属序列突变热点分析 序列突变热点(mutational hotspots)又称高变区(hyper-variable regions),可为设计准确、稳定而高效的分子标记和物种条形码提供参考(Niuetal., 2021)。使用DnaSP 6.0软件分析了山胡椒属核苷酸多态性位点(图5),序列比对总长度为156 450 bp,序列一致位点长度为150 149 bp,突变位点数为4 351,大约2.78%为多态性位点,核苷酸多样性值范围为0~0.019 79,平均值为0.004 08。鉴定的6个高变区中,3个位于LSC区(petA-psbJ、trnH-psbA、petK-psbI),3个位于SSC区(ycf1、ccsA-ndhD、rpl32-trnL),IR区没有高变区,表明单拷贝区核苷酸多态性明显高于IR区。

图5 山胡椒属叶绿体基因组序列核苷酸多态性分析Fig. 5 The nucleotide diversity(Pi) of chloroplast genome sequence of the genus Lindera
2.5 山胡椒属系统发育分析 利用IQtree软件使用最适模型K3Pu+F+R3构建的ML进化树,展示了山胡椒属物种的系统发育关系(图6)。山胡椒属物种归为5个聚类组: 聚类组1成员最多,为11个,其次是聚类组4,有8个成员,聚类组2和聚类组5分别有5个和4个成员,聚类组3仅有三桠乌药1种。四川山胡椒与黑壳楠(Linderamegaphylla)、红脉钓樟(L.rubronervia)、大果山胡椒(L.praecox)、绿叶甘橿(L.neesiana)、绒毛钓樟(L.floribunda)、山橿(L.reflexa)归在聚类组4,与黑壳楠亲缘关系最近。
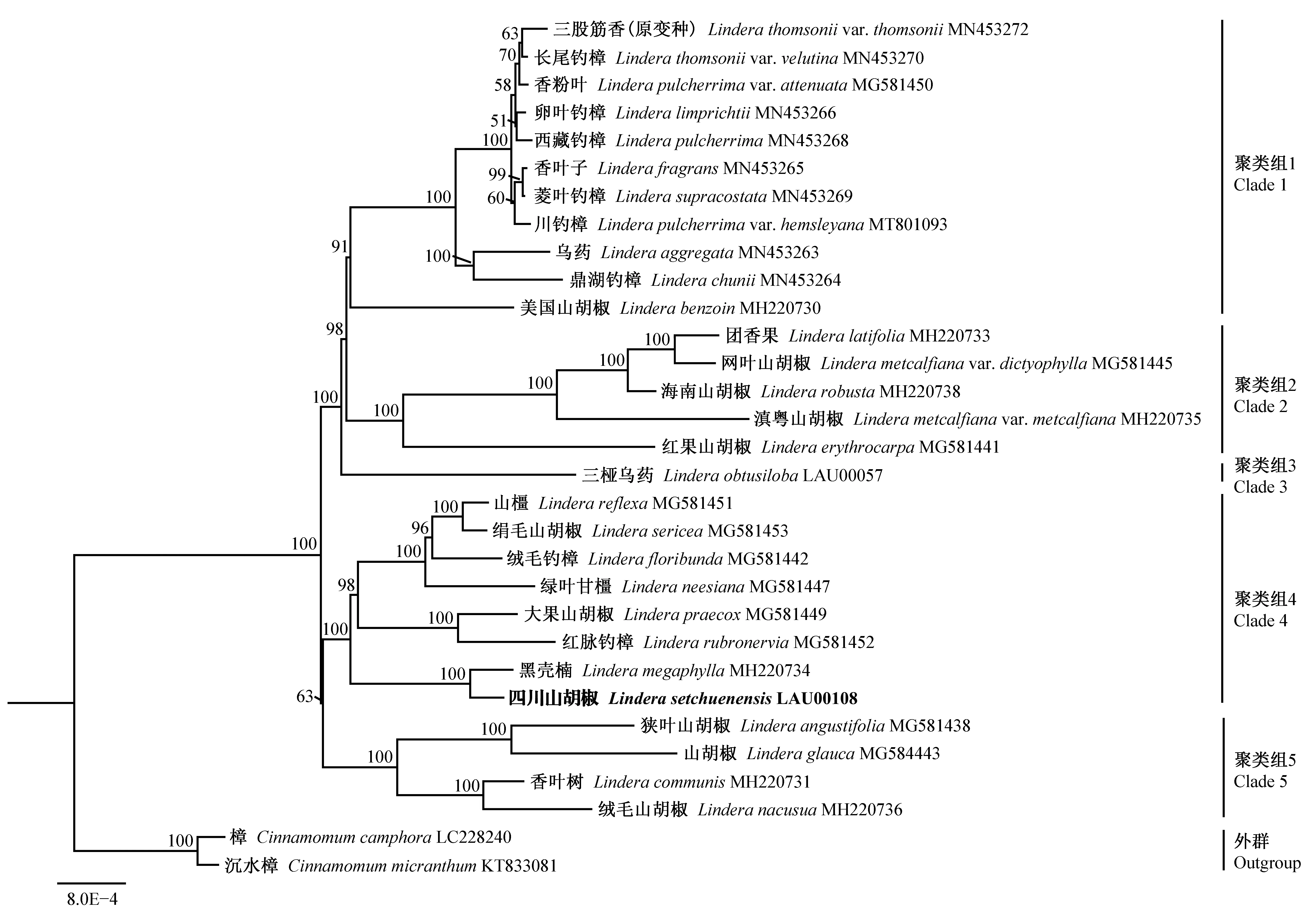
图6 基于最大似然法构建的山胡椒属叶绿体基因组系统发育树Fig. 6 The Maximum-likelihood tree of Lindera species based on analyses of the chloroplast genomes
3 讨论
植物叶绿体基因组为了解叶绿体功能、细胞内基因转移、物种多样性、植物资源保护和遗传育种提供分子基础(Danielletal., 2016)。本研究对四川山胡椒叶绿体基因组进行测序、组装和注释,获得了完整的叶绿体基因组数据。四川山胡椒叶绿体基因组大小为152 851 bp,具有典型的四分体结构,与之前公布的山胡椒属物种一致(Zhaoetal., 2018)。本研究以山胡椒叶绿体基因组(MG581443)为参考,对四川山胡椒叶绿体基因组进行注释,发现四川山胡椒基因组共编码125个基因,其中单拷贝基因112个,该结果与Zhao等(2018)和Tian等(2019)的注释略有不同,这与基因组注释时的参考基因组不同有关。
密码子使用偏性是物种对环境的长期适应性进化的结果,受多种因素的共同影响(Duanetal., 2021)。RSCU值反映了不同基因的密码子使用模式,数值越高表示该密码子使用频率越高(Sharpetal., 1987)。四川山胡椒叶绿体基因组总GC含量为39.15%,IR区GC含量明显高于单拷贝区,蛋白编码基因GC含量与叶绿体基因组接近,而GC3s明显较低(27.96%),这与山胡椒属其他物种类似(Zhaoetal., 2018; Tianetal., 2019; Xiaoetal., 2020)。31个高频密码子中以A/U(T)结尾的占94%,21个最优密码子中以A/U(T)结尾的占95%,这与其他植物叶绿体基因组密码子使用偏性(Duanetal., 2021)一致,表明叶绿体基因密码子第3位碱基存在不对称性,并表现出对A/U(T)的偏好性(Duanetal., 2021; 郑祎等, 2020; 周晓君等, 2020)。
序列重复广泛存在于植物叶绿体基因组中,在基因表达和调控中起着重要作用,由于这些区域存在高度序列多态性,对于研究分子进化和遗传多样性以及开发植物育种所必需的分子标记具有重要意义(蒋明等, 2020; Zhengetal., 2020; Zhuetal., 2021)。四川山胡椒叶绿体基因组长序列重复中,正向重复和回文重复含量较高,30 bp序列重复主要分布在单拷贝区,33 bp序列重复主要分布在IR区。基因组SSR位点含有较高比例的A/T,其中单碱基SSR以A/T型为主,55%的SSR位于基因间区,与Tian等(2019)结果类似,SSR较高的A/T含量可能与叶绿体基因组A/T含量较高有关。丰富的回文序列可能为基因组提供一些二级结构,而单核苷酸重复序列可能比其他SSR提供了更多的系统发育信号。基因组中6个高变区均位于单拷贝区,其中3个位于LSC,3个位于SSC,高变区trnH-psbA、psbK-psbI、petA-psbJ、ccsA-ndhD、ycf1与Zhao等(2018)和Tian等(2019)研究结果一致,可作为山胡椒属DNA条形码用于物种鉴定。所筛选的多态性SSR和高变区可为山胡椒属物种的鉴定和分子标记辅助育种提供参考。
山胡椒属具有重要的经济和药用价值。多个山胡椒属物种叶绿体基因组的公布,为山胡椒属系统发育研究提供了强有力的数据支撑。为了解四川山胡椒与山胡椒属其他物种之间的系统发育关系,本研究基于已公布的叶绿体基因组数据构建了山胡椒属物种的系统发育树。Zhao等(2018)将山胡椒属分为2个或4个支系,本研究基于进化树支持率将山胡椒属分为5个聚类组,四川山胡椒分在聚类组4,与黑壳楠亲缘关系最近,该结果与Tian等(2019)研究结果一致。根据崔鸿宾(1987)基于生物学性状的分类,山胡椒属分为5个组(Section),本研究中聚类组1、聚类组2和聚类组3均属于单系组,其中聚类组1物种属于三出脉组(LinderaSectionDaphnidium)(除美国山胡椒未记录外),聚类组2属于长梗组(LinderaSectionAperula),聚类组3仅有三桠乌药一种,属于掌脉组(LinderaSectionPalminerviae); 聚类组4为多系组,属于山胡椒组(LinderaSectionLindera)、三出脉组、球果组(LinderaSectionSphaerocarpae)和杯托组(LinderaSectionCupuliformes),聚类组5属于山胡椒组和多蕊组(LinderaSectionPolyadenia)。分析表明,仅基于是否落叶、叶片或顶芽等生物学特征进行的山胡椒属物种分类具有一定局限性,基于叶绿体基因组的系统发育分析对于明确山胡椒属物种的系统演化具有重要的参考价值。
4 结论
本研究基于高通量测序技术,从头测序并组装了完整的四川山胡椒叶绿体基因组。该基因组呈典型的四分体结构,基因组全长为152 851 bp,LSC、SSC和IR区大小分别为93 742、18 977、20 066 bp。27对长序列重复中,正向重复最多,其中33 bp和30 bp的序列在正向重复和回文重复均占较高比例。181个SSR位点中,单碱基SSR数目最多,多数SSR位于基因间区。四川山胡椒叶绿体基因组共编码125个基因,偏好以A或U(T)结尾的密码子。6个高变区中,3个位于LSC区(petA-psbJ、trnH-psbA、petK-psbI),3个位于SSC区(ycf1、ccsA-ndhD、rpl32-trnL)。系统发育关系上,山胡椒属物种归为5个聚类组,四川山胡椒与黑壳楠亲缘关系最近。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
福建农业学报(2021年6期)2021-08-18
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
发明与创新·中学生(2019年6期)2019-06-26
生物学教学(2018年2期)2018-08-07
安徽农业科学(2018年1期)2018-05-14
江苏农业科学(2016年2期)2016-04-11
