
不同改良剂对旱地苹果园温室气体排放的影响
2021-02-04 10:16:38李钊刘帅丁艳宏孙文浩高晓东赵西宁
农业环境科学学报 2021年1期
李钊,刘帅,丁艳宏,孙文浩,高晓东,赵西宁*
(1.西北农林科技大学水利与建筑工程学院,陕西 杨凌 712100;2.西北农林科技大学旱区农业水土工程教育部重点实验室,陕西 杨凌 712100;3.中国科学院水利部水土保持研究所,陕西 杨凌 712100)
气候变化已成为全球关注的热点问题,其给人类赖以生存的环境带来很多严峻问题和挑战。温室气体(CH4、CO2和N2O)排放是引起气候变化的主要原因之一[1],因此如何减少温室气体排放引起了众多学者的关注。研究表明,我国农业活动所带来的碳排放占排放总量的16%,其中由农业活动造成的CO2、CH4和N2O 排放量分别约占全球排放总量的12%、50%和60%[2-3]。国家实施退耕还林(草)工程以来,苹果产业已成为黄土高原地区有效应对退耕后耕地大面积减少及促进区域经济发展的支柱产业,也是保障国家退耕还林(草)工程持续健康发展的重要着力点。以陕西省为例,截至2018年,在“北扩西进”战略布局推进下,苹果林面积达59.757万hm2,产量突破1 008.69万t,占我国总产量的四分之一,占世界总产量的七分之一[4]。但是在苹果林推广过程中,粗放的管理方式不仅导致产量低下,更加剧了土壤退化、土壤肥力下降,增加了温室气体排放,进而导致一系列环境问题[5]。因此,在果园管理过程中如何减少土壤温室气体排放成为黄土高原亟待解决的问题之一。
大量学者已经研究了如何通过改善管理及耕作措施、合理施肥灌溉等多种途径减少土壤温室气体的排放,如秸秆覆盖、秸秆还田、水分管理等[6-8]。目前,关于施加土壤改良剂对温室气体排放的研究也较多,但由于改良剂种类繁多及研究环境的不同,研究结果也有所差异。众多研究表明,生物炭添加到土壤后可明显抑制土壤温室气体排放[9]。Barracosa 等[10]的研究指出,施加生物炭后,N2O 的排放量可减少36%,但是对CO2、CH4的影响不明显,这可能与施加环境有关。目前,有关生物炭对温室气体排放的影响研究多集中在早稻田和农田[11-12]。微生物菌剂类如胶质芽孢杆菌与枯草芽孢杆菌等改良剂的相关研究主要关注其对土壤理化性质的改良效果,并未对其在温室气体排放的影响方面给予充分的研究,在果园中的研究则更是鲜有报道[13-14]。因此,本研究选用几种不同的土壤改良剂,探讨旱地果园温室气体排放效果,以期为果园固碳减排提供理论依据。
1 材料与方法
1.1 试验设计
试验在陕西省延安市宝塔区河庄坪镇余家沟村山地苹果园示范基地(109°21′24″ E,36°41′15″ N,海拔1 277 m)进行,该地区为典型的黄土高原丘陵沟壑区,属于干旱半干旱气候,年平均风速1.2 m·s-1,年均降雨量500 mm 左右,年内降雨主要集中在7—9月,占全年降水量的73%左右,年均气温为9.4 ℃,年总辐射量为 492.6 kJ·cm-2,无霜期 170~186 d。土壤类型为黄绵土。试验地土壤理化性质:容重1.3 g·cm-3,pH 7.81,土壤有机碳含量2.69 g·kg-1,全氮含量0.3 g·kg-1,田间持水量约24%。
试验共设置4 个处理,布设在同一水平梯田上,分别为空白对照处理(CK)、施加胶质芽孢杆菌处理(JZ)、施加枯草芽孢杆菌处理(KC)及施加生物炭处理(BC)。由于研究区存在苹果树枝过剩问题,试验选择以苹果树枝为主要原材料的生物炭为其中一种土壤改良剂,该生物炭源自陕西亿鑫生物能源科技开发有限公司,在500 ℃高温无氧条件下裂解,粒径为0.02~2 mm,有机碳含量306.25 g·kg-1,全氮含量8.97 g·kg-1,pH 8.98;其他两种改良剂均来自济南金华峰辉生物科技有限公司,胶质芽孢杆菌和枯草芽孢杆菌的有效活菌数分别为≥2.0×1010CFU·g-1和≥5.0×109CFU·g-1。供试果树为山地红富士苹果,树龄15 a,行距为4.5 m×4.5 m,树高平均2.5 m,平均冠幅半径为2.3 m。试验地为旱地雨养果园,修剪拉枝和土壤管理等措施均完全相同,为避免施肥影响试验结果,果园均未施肥。为使改良剂发挥作用且降低对果树根系的伤害,施加改良剂的位置在树冠2/3处,即距树干150 cm 的环形带,环形带宽度为60 cm,通过人工方式将改良剂翻入土壤,并与0~60 cm 土层土壤混合均匀。参考已有文献的最适用量[15-16],确定生物炭用量为65 t·hm-2,胶质芽孢杆菌及枯草芽孢杆菌用量均为20 t·hm-2。每个处理选取3 株长势相似的果树作为重复,在果树施加改良剂的环带上随机设置1 个温室气体排放监测点,共设置12 个;土壤水分监测点布设在环带上,每个重复环带上等分3点设置,共计36个。在温室气体排放监测点处布置1组地温计,共布置12组。具体试验布置如图1所示。
试验期间研究区温度和降雨量变化如图2 所示。2019 年全年降水累计总量为608 mm,降水主要集中在8月份,其降水量约占试验期内降水总量的50%。
1.2 测定项目
1.2.1 温室气体监测
采用静态暗箱-气相色谱法测定土壤CH4、N2O和CO2排放通量,采样时间为6—10月,每7 d进行一次,气体采集时间为8:30—11:30。改良剂施用后在环带上选取地表相对平坦的区域安放底座,采样箱规格为50 cm×50 cm×50 cm,顶箱外覆盖白色优质保温棉,防止太阳辐射下采样箱壁和采样箱内空气温度升高,保证箱内温度在测量期间恒定,以减小温度带来的误差;采样箱内装有小型风扇,采样时通电持续转动,以充分混匀箱内气体。
采样时将采样箱放置在底座上并用水密封,扣箱后0、10、20、30 min时用60 mL注射器通过插在采样箱中部采气孔的橡胶长软管从采集箱内抽取200 mL气体转存于铝塑复合气袋中,并于当天采用Agilent 7890A气相色谱仪测定气体样品中的CH4、CO2和N2O。
采样当天应为天气晴朗或多云天气,降雨(一般超过20 mm)后分别在第1、2、3、5 d 和7 d 采集气体,若遇连续间隔降雨,则在降雨期间连续监测,以最后一次降雨为准,连续测定后恢复之前的频率。
1.2.2 土壤含水量测定
采用Trime-TDR 土壤水分测定仪监测土壤含水量,测量深度为0~100 cm 土层,每 10 cm 土层测定一次,测定时间同采气时间同步。土壤孔隙含水率[17]计算公式如下:
式中:WFPS为土壤孔隙含水率,%;θ为各土层体积含水率,%;γ为土壤容重,g·cm-3。
1.2.3 土壤温度
采用直角地温计测量土壤温度,在收集温室气体时同步测定 5、10、15、20 cm 和 25 cm 处的土壤温度,分别测定温室气体开始及结束收集时的土壤温度。
1.2.4 气象数据
由布设在田间的自动气象站监测气象数据,包括降雨量、风速、气压、太阳辐射、空气温湿度等。
1.3 统计分析
(1)温室气体排放通量采用Adviento-Borbe 等[18]的计算方法,计算公式为:
式中:F为气体排放通量,CH4、CO2和 NO2的单位分别为 mg·m-2·h-1、mg·m-2·h-1和 µg·m-2·h-1;H为采样箱高度,m;M为气体的摩尔质量,g·mol-1;P为采样点气压,Pa;R为普适气体常数,8.314 Pa·m3·mol-1·K-1;T为采样时箱内温度,℃。
温室气体累积排放量(ω)计算公式如下:
式中:ω为土壤 CH4(CO2或 N2O)的排放总量,kg·hm-2;Ri为第i次采样时CH4(CO2或N2O)的日排放通量,mg·m-2·d-1;Di为第i次采样到第i+1 次采样间隔的天数,d。
(2)使用SPSS 25.0 统计软件对试验数据进行方差分析,使用最小显著性差异法(LSD)在0.05 的显著性水平上进行分析。
2 结果与分析
2.1 不同改良剂对土壤水热的影响
不同处理土壤WFPS 变化趋势基本一致(图3),在6月末—8月初土壤水分含量较低,受降雨影响,在8—10 月土壤水分含量较高。CK、JZ、KC 和 BC 处理0~100 cm 平均WFPS 分别为43.94%、47.98%、39.52%和41.24%。不同土层平均WFPS大小分别为:表层JZ>BC>KC>CK,中层 JZ>CK>KC>BC,深层 JZ>CK>BC>KC。且CK、JZ 和 KC 均表现为中层>深层>表层,BC表现为表层>深层>中层。与CK相比,JZ、KC和BC表层(0~20 cm)WFPS 分 别 增 加 19.41%、1.83% 和16.29%;中层(20~60 cm)土壤 JZ 增加7.47%,KC 和BC 分别降低16.32%和18.13%;深层(60~100 cm)土壤 JZ 增 加 2.46%,KC 和 BC 分 别 降 低 13.43% 和12.29%。JZ WFPS 高于其他处理,在 0~20 cm 土层较CK 增加效果显著(P<0.05);BC 在0~30 cm 水分较高,10~20 cm 保水效果显著(P<0.05),在30~100 cm 低于CK,50~60 cm 差异显著(P<0.05);KC 在0~20 cm 高于CK,而在 20~100 cm 低于 CK,在 50~70 cm 时差异显著(P<0.05)。
不同处理0~25 cm 平均土壤温度变化均呈下降趋势(图4),CK、JZ、KC 和BC 处理平均土壤温度分别为 17.4、17.5、17.7 ℃和 16.9 ℃。0~5 cm 土壤温度受气温影响波动幅度较大,20~25 cm 土壤波动幅度较小,较为稳定。6 月 25 日—7 月 15 日 BC 处理 10~25 cm 土壤温度较 CK 低;在 8 月 26—27 日累计降雨 20 mm 后,各处理土壤温度下降,BC 较 CK 在 10~20 cm土层温度低 2.85 ℃,JZ 和 KC 较 CK 在 0~10 cm 土层温度高0.86 ℃和 1.57 ℃;在 10 月18—19 日累计降雨0.4 mm 后,BC较其他处理土壤温度最低,为19.08 ℃,但是不同处理与不同土层之间均无显著差异。
2.2 不同改良剂对温室气体排放的影响
2.2.1 对CH4排放的影响
不同处理CH4排放通量均为负值(图5),在旱地果园中表现为CH4的汇。不同处理变化趋势表现出相同的季节变化规律,但各处理间CH4的吸收总量差异不显著,CK、JZ、KC 和 BC 吸收总量分别为 1.64、1.38、1.42 kg·hm-2和1.17 kg·hm-2,3 种改良剂处理低于CK 处理。可见,土壤改良剂会降低旱地果园CH4吸收效果。
2.2.2 对CO2排放的影响
不同处理CO2排放通量(图6)在8 月达到最大,之后排放通量逐渐减小。在8月5—11日的连续监测中,5日各处理CO2排放较少,随着温度的回升,8日排放最高达到 314.52~704.53 mg·m-2·h-1。在 8 月 27 日—9月1日的连续监测中,27日排放通量达到最低,随后排放量逐渐升高。CO2累计排放通量大小依次为CK>JZ>KC>BC,BC 处理累计排放总量为5 922.2 kg·hm-2,CK、JZ 和 KC 较 BC 分别增加 40.5%、40.4% 和34.9%。与CK 处理相比,BC 极显著降低了CO2排放(P<0.001),JZ和KC无显著差异。
2.2.3 对N2O排放的影响
不同处理土壤N2O 排放整体趋势一致,均随着时间的延长排放通量逐渐降低(图7)。在8 月5—11 日的连续监测中,5 日出现N2O 排放通量的峰值,CK、JZ、KC 和 BC 处理分别为 24.28、38.09、22.09 µg·m-2·h-1和49.46 µg·m-2·h-1。8月27日—9月1日各处理维持在较低的水平。CK、JZ、KC 和BC 累计排放通量分别为 0.31、0.29、0.34 kg·hm-2和 0.64 kg·hm-2。BC 排放通量较CK 增加103%(P<0.01),JZ、KC 与 CK 处理无显著差异(P>0.05)。
2.3 环境因子对温室气体排放的综合影响效应及评价
综合分析土壤温度和土壤水分对气体排放通量的影响见表1、表2、表3,所有处理CO2排放通量与土壤温度呈极显著正相关(P<0.01),JZ、KC 和BC 处理CO2通量与表层土壤水分呈极显著负相关(P<0.01)。通过分析N2O 排放通量发现,CK 处理与各层土壤水分均呈显著负相关(P<0.05),KC处理只与0~60 cm土壤水分呈显著负相关(P<0.05),JZ 和BC 处理与土壤温度呈显著正相关。不同处理土壤温度和水分与CH4排放通量无显著相关性。

表1 不同处理土壤温度、水分与CH4相关分析Table 1 Correlation analysis of soil temperature,moisture and CH4 emission flux under different treatments

表2 不同处理土壤温度、水分与CO2相关分析Table 2 Correlation analysis of soil temperature,moisture and CO2 emission flux under different treatments
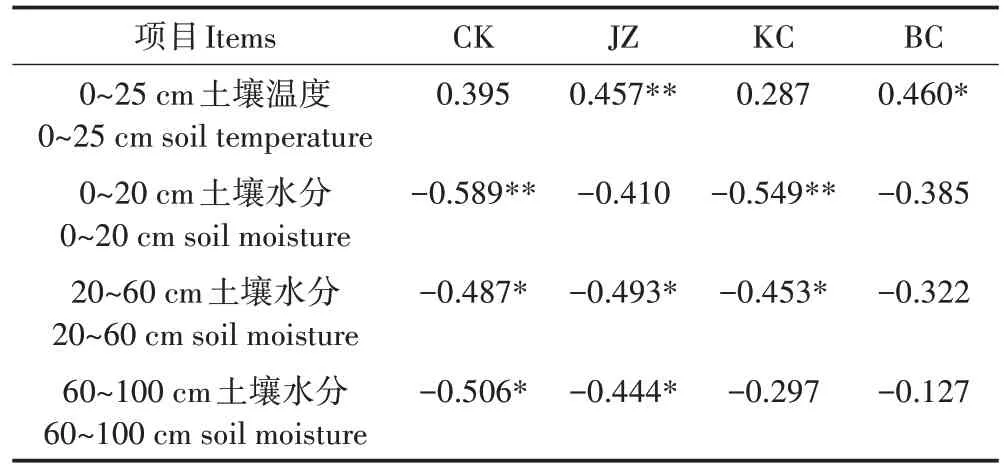
表3 不同处理土壤温度、水分与N2O相关分析Table 3 Correlation analysis of soil temperature,moisture and N2O emission flux under different treatments
3 讨论
3.1 不同改良剂对土壤水热的影响
土壤改良剂对土壤保水作用效果显著,所以广泛应用于土壤的改良[19]。在本研究中,JZ处理表层土壤保水效果最佳,可能是因为胶质芽孢杆菌能够产生胞外多糖和荚膜多糖,对改善土壤结构、促进土壤团聚体的形成具有一定的作用效果,进而提高了土壤的保水能力[20]。BC 在 0~20 cm 保水效果较好,这与生物炭本身特性相关,多孔隙生物炭能够吸持土壤水分,同时降低土壤容重[21],增大了土壤持水能力;而在50~60 cm 水分较CK 显著降低,可能是因为土壤表层中的生物炭吸持了大量水分,导致入渗量减少,水分大都集中在土壤表层,深层土壤含水量降低。枯草芽孢杆菌自身会产生絮凝剂,吸水后呈现凝胶状态,可以保水减渗,同时改善土壤团粒结构,增大水稳性团聚体占比,进而提高表层土壤水分。KC 处理表层土壤水分略高,深层较低,50~70 cm 土层水分显著降低,可能是因为枯草芽孢杆菌促进果树根系的生长发育[22],加快了根系对土壤水分的吸收利用,导致该层土壤耗水加剧。
本研究中,生物炭有降低土壤温度的效果,但并不显著。这可能是因为生物炭自身的多孔隙结构导致土壤容重减小,土壤总孔隙度增加[23],良好的孔隙结构可以保持水分,增大比热容,并滞后升温过程。Ding等[24]研究表明,生物炭减少进入土壤的热量以及热量散失,从而减少土壤温度的波动。胶质芽孢杆菌和枯草芽孢杆菌在0~20 cm 土层会增加土壤温度,在20~25 cm 土层则呈现降温的效果,但无显著差异。这可能是因为两种微生物的数量增多、活性增强、呼吸作用增强,致使更多的热量产生。
3.2 不同改良剂对CH4排放的影响
目前,关于旱地CH4的排放多表现为汇[25],与本研究结果一致,但施加改良剂后吸收量较小。本研究分析了CH4排放与土壤温度、土壤水分的相关性,但均表现为不相关,这进一步说明土壤温度和土壤水分在本研究中不是影响CH4排放的主要因素。CH4主要是在缺氧条件下由产甲烷菌产生。本研究中施加改良剂后CH4排放量均大于CK,这主要是由于施加改良剂增加了土壤微生物和土壤酶的活性[13-14,26],微生物活动的加剧增加了土壤耗氧量,使土壤中可能形成厌氧环境,导致产甲烷菌活动加剧,促进了CH4排放。本研究中,3种改良剂CH4累计吸收量大小为:BC<JZ<KC。大部分研究表明,施加生物炭后CH4吸收量增大[12],但本研究结果与之相反,这主要是由于生物炭与其他改良剂相比,施入土壤后使土壤呈弱碱性(CH4最适氧化pH 范围为4~6),不利于CH4吸收。本研究中,BC 处理对 CH4吸收效果最差,与 Song 等[27]研究得出旱地土壤施加生物炭后CH4吸收量大幅增加的结果不一致,这可能与土壤类型有关。本研究中,JZ 和KC 处理CH4吸收量低于CK,可能是因为微生物菌剂施入土壤后,会增加有益菌的数量[28]和微生物群落的多样性[29],导致微生物呼吸加剧,形成厌氧环境,从而不利于土壤CH4的氧化。
3.3 不同改良剂对CO2排放的影响
生物因素、非生物因素及人为因素都会对土壤CO2排放产生重要影响,而土壤温度和水分两种环境因子在非生物因素中起主要作用[30]。研究表明,较低的土壤温度会减少CO2排放[31]。在本研究中,各处理CO2排放与土壤温度呈极显著正相关,BC 处理CO2排放量最少,可能是因为土壤温度较低,微生物活动减弱,土壤呼吸速率降低,减少了土壤CO2排放。CK、JZ和KC 处理土壤温度相差较小,土壤呼吸速率基本一致,没有显著增加或减少CO2排放。旱地土壤水分是制约微生物活性的重要因素[32],微生物活性对水分的敏感性远大于温度。本研究中,施加改良剂处理CO2排放与表层土壤水分呈极显著负相关。BC 处理显著增加表层土壤水分,并显著降低了CO2排放。可能是水分增加后,微生物活动在一定程度上加剧,但是生物炭本身大孔隙、大比表面积会吸附土壤中的CO2以及活性酶,降低碳分解速率[33],也会促进土壤形成芳烃等大分子物质[34],不利于微生物的分解作用,减少土壤碳排放。同时,较高的C/N 限制微生物活动,降低了土壤呼吸速率[35]。JZ处理显著增加土壤水分,但是CO2排放同CK 无显著差异,可能是因为胶质芽孢杆菌在增加微生物数量、促进土壤呼吸的同时,改善了土壤结构,促进了团聚体的形成[20],因此并没有因为微生物呼吸的加剧而增加CO2排放。KC 处理表层水分、CO2排放同CK 无显著差异,可能是较低的表层土壤水分限制了微生物的活性,不利于微生物呼吸增加 CO2排放[29]。
3.4 不同改良剂对N2O排放的影响
土壤中N2O 主要由硝化作用和反硝化作用产生,而土壤水分、土壤温度会对其产生重要影响。本研究中,BC 和JZ 处理土壤温度和N2O 排放显著正相关。BC 和JZ 处理显著增加表层土壤水分,表层土壤平均WFPS 分别为44.82%和45.66%,适宜土壤微生物活动,此时温度成为N2O 排放的主要影响因素。本研究中,BC 处理N2O 排放总量显著高于其他处理,这与前人研究结果[36-37]不一致,可能是土壤温度处于硝化作用最适温度范围内,硝化作用加剧,同时,生物炭会提高土壤有机碳含量,为硝化反硝化作用提供良好的条件,进而增加 N2O 排放[38]。JZ 处理温度较 BC 高,但施加大量胶质芽孢杆菌后,微生物间营养竞争加强[29],因此不利于硝化细菌产生N2O。本研究中,CK 和KC处理土壤水分和N2O 排放显著负相关,KC 处理表层土壤水分较低,对微生物活动有一定影响,同时,土壤施加枯草芽孢杆菌后,竞争作用、拮抗作用及溶菌作用[39]等均不利于硝化作用产生N2O。
4 结论
(1)生物炭和胶质芽孢杆菌可以显著增加土壤表层含水量,各处理下土壤温度没有显著差异。
(2)各处理CH4吸收总量差异不显著,累计吸收量大小为 BC<JZ<KC<CK。CO2排放总量 BC 处理显著低于其他处理,N2O 排放总量显著高于其他处理,JZ、KC和CK处理间无显著差异。
(3)旱地果园CH4排放与土壤水分和温度无显著相关性。CO2排放与土壤温度极显著正相关,JZ、KC和BC 处理CO2排放与表层土壤水分呈极显著负相关。JZ 和BC 处理N2O 排放与土壤温度极显著正相关,CK 和KC 处理N2O 排放与表层土壤水分呈极显著负相关。
综上,生物炭在保水、固碳及减排上综合效果最好,是一种较为合适的土壤添加剂。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:05:10
河南建材(2021年2期)2021-03-11 08:18:50
辽宁林业科技(2017年4期)2017-06-22 10:23:36
国际汉学(2017年2期)2017-01-28 04:05:39
高原山地气象研究(2016年2期)2016-11-10 06:06:27
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
大气科学(2015年5期)2015-12-04 03:04:44
塔里木大学学报(2014年3期)2014-03-11 18:47:27
自然资源遥感(2014年2期)2014-02-27 11:56:12
自然资源遥感(2014年2期)2014-02-27 11:56:11
